
The Drosophila Retinoblastoma Binding Protein 6 Family Member Has Two Isoforms and Is Potentially Involved in Embryonic Patterning


Abstract
:1. Introduction
2. Results
2.1. Transcriptional Start Site Mapping by 5' Rapid Amplification of cDNA Ends and Functional Analysis of the Promoter Region


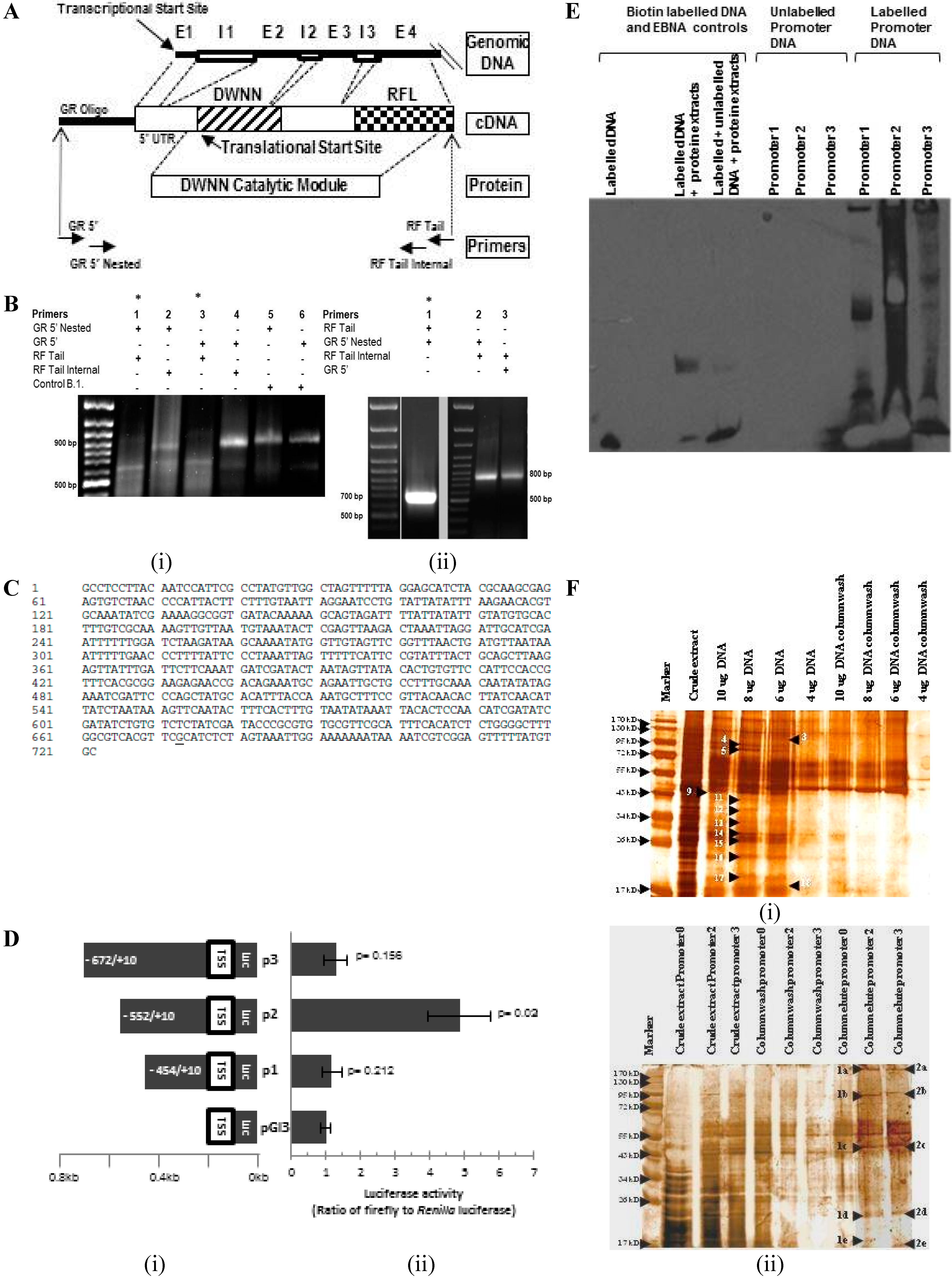{kind=link}
{kind=link}
{kind=link}
| Band | MS Prediction | Accession Number | Size (AA) | ProtScore |
|---|---|---|---|---|
| TRANSCRIPTION FACTORS | ||||
| 4 | DREF transcription factor | O96083 | 709 | 2.03 |
| TRANSCRIPTION CO-ACTIVATOR | ||||
| 2c | Pyruvate Kinase | AAO24935 | 2 | |
| OTHER NUCLEAR PROTEINS | ||||
| 5 | Heat shock protein 83 | P02828 | 717 | 1.66 |
| 14 | Heat shock protein 27 | HHFF27 | 213 | 2 |
| 17 | Ribosome biogenesis protein WDR12 homolog | Q9VKQ3 | 420 | 1.11 |
| Similar to PHAX | Q8SYG4 | 479 | 1.17 | |
| PROTEINS ASSOCIATED WITH PRE-MRNA PROCESSING | ||||
| 3 | CG4266 | Q9W2K4 | 1215 | 2 |
| OTHER PROTEINS | ||||
| 1a | Mind-meld | CG9163-PB Q7KUY7 | 783/840 | 0.93 |
| 1c | With coiled coil and lipase domain | Q29QE3, GH19966p | 482 | 2 |
| 1d | elongation factor 1alpha48D | AAF58608 | 15.56 | |
| Actin 87E-fruit fly | S04538 | 1.92 | ||
| Q9VEV0, CG10405-PA | 252 | 0.8 | ||
| 1e | Unknown (CG17374) | Q7PLB8 | 2284 | 2 |
| Tubulin | Q9V7Y7,CG15611 | 508 | 1.6 | |
| 2a | Unknown | Q9VGU9 | 332 | 2 |
| 2c | CG9277-PA (Tubulin family) | Q9V8V3 | 10 | |
| ATP synthase subunit β | Q05825.3 | 505 | 3.2 | |
| 2d | Unknown | Q9VLX3 | 281 | 1.26 |
| Hypothetical protein | Q9NG77 | 81 | 1.06 | |
| Q7KQM6 | 0.74 | |||
| 2e | Unknown | Q9VU21 | 308 | 2.04 |
| Unknown | Q4ABH1 | 11,707 | 0.77 | |
| Unknown | Q9VQ84 | 124 | 2 | |
| 9 | CG18063-PA, isoform A | Q9VJM6 | 337 | 0.9 |
| 11 | Unknown | Q1RKR2 | 270 | 2 |
| Q9VEJ9 | 753 | 0.84 | ||
| 13 | Q9VP46 | 1.05 | ||
| 16 | Q9VJM6/ CG18063-PA | 337 | 0.75 | |
| 17 | Unknown | Q9VQ84 | 124 | 1.64 |
| 18 | With the Domain of Unknown Function and coiled coil | Q9VQ83 | 559 | 1.52 |
| Group | Genes | References |
|---|---|---|
| Embryonic patterning | Caudal, bicoid, tailless, hunchback | [21,22] |
| Segment polarity | Engrailed, LEF-1 (pangolin) | [23,24] |
| Regulation of development | Adf-1 (Myb) | [25] |
| Germ-line development and sex determination | dsxf/dsxm, sry- = β, CF-2 | [26,27,28] |
| Replication | DREF | [29] |
| Myogenesis | MEF2, twist, CF2 | [27,30,31,32,33] |
| Chromatin structure | BEAF, GAGA | [34,35,36] |
2.2. Differential Expression of Snama during Drosophila Development

2.3. Subcellular Localization of SNAMA and Potential Interacting Partners

| Sample 1 | UniProt Accession Code | Name | Molecular Weight (kDa) | ProtScore |
|---|---|---|---|---|
| CBB55 | Q9TWZ1_DROME | D-ERp60 (PDI) | 55.373 | 2.26 |
| CBB35 | Q9UAN1_DROME | 60S RIBOSOMAL PROTEIN L22 | 30.611 | 2 |
| CBB43 | RL4_DROME | 60S RIBOSOMAL PROTEIN L4 | 45.026 | 2.37 |
| SS95 | Q9VKQ3_DROME | RIBOSOME BIOGENESIS PROTEIN WDR12 HOMOLOG | 47.222 | 1.47 |
| SS22 | Q94883_DROME | DNA REPLICATION-RELATED ELEMENT FACTOR (DREF) | 80.727 | 1.24 |
| SS95 | HP1_DROME | HETEROCHROMATIN PROTEIN 1 | 23.185 | 1.31 |
| TRANSCRIPTION CO-ACTIVATORS | ||||
| CBB90 | PYG_DROME | GLYCOGEN PHOSPHORYLASE | 96.997 | 2 |
| CBB55 | KPYK_DROME | PYRUVATE KINASE | 57.44 | 2.75 |
| SS18 and CBB33 | Q9W2K4_DROME | CG4266-PA | 130.427 | 1.74 and 0.89 |
| CBB28 | HSP27_DROME | HEAT SHOCK PROTEIN 27 | 23.617 | 4 |
| SS95 | Q8SYG4_DROME | PHOSPHORYLATED ADAPTER FOR RNA EXPORT (PHAX) | 53.965 | 1.17 |
| SS18 and SS26b | Q9W1K4_DROME | EGALITARIAN | 112.129 | 1.43 and 1.15 |
| SS18, SS21 and SS22 | Q24156_DROME | STONEWALL | 112.913 | 0.87, 0.94 and 1.08 |
| CBB28 | VDAC_DROME | VOLTAGE-DEPENDENT ANION-SELECTIVE CHANNEL | 30.55 | 8.13 |
| CBB43 | VIT2_DROME | VITELLOGENIN-2 | 49.66 | 14.35 |
| CBB55 | TBB1_DROME | TUBULIN Β-1 CHAIN | 50.147 | 16.55 |
| CBB90 | TERA_DROME | TRANSITIONAL ENDOPLASMIC RETICULUM ATPASE TER94 | 88.859 | 9.72 |
| SS95 | Q9W003_DROME | SPINOPHILIN | 233.209 | 1.86 |
| SS22d | Q4ABH1_DROME | MUSCLE-SPECIFIC PROTEIN 300, isoform D | 1405.148 | 0.97 |
| SS33 | Q9VXL1_DROME | MIND-MELD, isoform C | 150.547 | 0.87 |
| CBB28 | ADT_DROME | ADP, ATP CARRIER PROTEIN | 34.215 | 5.64 |
| SS95b | Q6IGH5_DROME | HDC06258 | 17.442 | 0.78 |
| SS22 | Q9VQ83_DROME | CG10874-PA | 35.041 | 1.57 |
| SS38 | Q9VSY1_DROME | CG4022-PA | 57.294 | 2 |
| SS26 | Q4V5H1_DROME | PEPTIDYL-PROLYL CIS-TRANS ISOMERASE | 20.182 | 1.62 |
| CBB90 | EF2_DROME | ELONGATION FACTOR 2 | 94.459 | 27.74 |
| CBB43 | EF1G_DROME | ELONGATION FACTOR 1-GAMMA | 48.968 | 6 |
| SS29 | EF1A2_DROME | ELONGATION FACTOR 1-ALPHA 2 | 50.663 | 2.5 |
3. Discussion
4. Experimental Section
4.1. Maintenance of Fly Stocks
4.2. RNA Extraction and RT-PCR
4.3. 5' Rapid Amplification of cDNA Ends
4.4. Preparation of Reporter Constructs and Assays
4.5. Fractionation of Cell Extracts
4.6. Modification of RS Domain Proteins: Acetylation and Phosphorylation and Mg+2-Precipitation
4.7. Electromobility Shift Assays and Streptavidin Chromatography
4.8. Preparation of Antibodies against SNAMA-DCM
4.9. Co-Immunoprecipitation Assays
4.10. Preparation of Proteins from Gels and Identification via Tandem Mass Spectrometry
4.11. Western Blot Analysis
4.12. Immunocytochemical Staining of Whole Mount Embryos and Ovaries
4.13. Immunofluorescence and Confocal Microscopy
4.14. Bioinformatics Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Mbita, Z.; Meyer, M.; Skepu, A.; Hosie, M.; Rees, J.; Dlamini, Z. De-regulation of the RBBP6 isoform 3/DWNN in human cancers. Mol. Cell. Biochem. 2012, 362, 249–262. [Google Scholar] [CrossRef] [PubMed]
- Yoshitake, Y.; Nakatsura, T.; Monji, M.; Senju, S.; Matsuyoshi, H.; Tsukamoto, H.; Hosaka, S.; Komori, H.; Fukuma, D.; Ikuta, Y.; et al. Proliferation potential-related protein, an ideal esophageal cancer antigen for immunotherapy, identified using complimentary DNA microarray analysis. Clin. Cancer Res. 2004, 10, 6437–6448. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Tang, H.; Wu, Z.; Zhou, C.; Jiang, T.; Xue, Y.; Huang, G.; Yan, D.; Peng, Z. Overexpression of RBBP6, alone or combined with mutant TP53, is predictive of poor prognosis in colon cancer. PLoS ONE 2013, 8, e66524. [Google Scholar] [CrossRef] [PubMed]
- Motadi, L.R.; Bhoola, K.D.; Dlamini, Z. Expression and function of retinoblastoma binding protein 6 (RBBP6) in human lung cancer. Immunobiology 2011, 216, 1065–1073. [Google Scholar] [CrossRef] [PubMed]
- Harutyunyan, A.S.; Giambruno, R.; Krendl, C.; Stukalov, A.; Klampfl, T.; Berg, T.; Milosevic, J.D.; Chen, D.; Gisslinger, B.; Gisslinger, H.; et al. Germline RBBP6 mutations in myeloproliferative neoplasms. Blood 2013, 122, 267. [Google Scholar]
- Simons, A.; Melamed-Bessudo, C.; Wolkowicz, R.; Sperling, J.; Sperling, R.; Eisenbach, L.; Rotter, V. PACT: Cloning and characterization of a cellular p53 binding protein that interacts with Rb. Oncogene 1997, 14, 145–155. [Google Scholar] [CrossRef] [PubMed]
- Witte, M.M.; Scott, R.E. The proliferation potential protein-related (P2P-R) gene with domains encoding heterogeneous nuclear ribonucleoprotein association and Rb1 binding shows repressed expression during terminal differentiation. Proc. Natl. Acad. Sci. USA 1997, 94, 1212–1217. [Google Scholar] [CrossRef] [PubMed]
- Mather, A.; Rakghotho, M.; Ntwasa, M. SNAMA, a novel protein with a DWNN domain and a RING finger-like motif: A possible role in apoptosis. Biochim. Biophys. Acta 2005, 1727, 169–176. [Google Scholar] [CrossRef] [PubMed]
- Pretorius, A.; Kaur, M.; Wamalwa, M.; February, M.F.; Essack, M.; Bajic, V.B.; Rees, D.J.G. Functional analysis and characterization of the human RBBP6 promoters based on a combination of molecular biology and in silico approaches provide additional evidence for RBBP6 role in apoptosis. J. Biosci. 2013, 1, 2251–3159. [Google Scholar]
- Ntwasa, M. The retinoblastoma binding protein 6 is a potential target for therapeutic drugs. Biotechnol. Mol. Biol. Rev. 2008, 3, 24–31. [Google Scholar]
- Chibi, M.; Meyer, M.; Skepu, A.; Rees, D.J.G.; Moolman-Smook, J.C.; Pugh, D.J.R. RBBP6 Interacts with multifunctional protein YB-1 through its RING finger domain, leading to ubiquitination and proteosomal degradation of YB-1. J. Mol. Biol. 2008, 384, 908–916. [Google Scholar] [CrossRef] [PubMed]
- Pugh, D.; Ab, E.; Faro, A.; Lutya, P.; Hoffmann, E.; Rees, D.J. DWNN, a novel ubiquitin-like domain, implicates RBBP6 in mRNA processing and ubiquitin-like pathways. BMC Struct. Biol. 2006, 6. [Google Scholar] [CrossRef]
- Di Giammartino, D.C.; Li, W.; Ogami, K.; Yashinskie, J.J.; Hoque, M.; Tian, B.; Manley, J.L. RBBP6 isoforms regulate the human polyadenylation machinery and modulate expression of mRNAs with AU-rich 3' UTRs. Genes Dev. 2014, 28, 2248–2260. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.; Reifegerste, R.; Moses, K. Characterization of Drosophila mini-me, a gene required for cell proliferation and survival. Genetics 2006, 173, 793–808. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Deng, B.; Xing, G.; Teng, Y.; Tian, C.; Cheng, X.; Yin, X.; Yang, J.; Gao, X.; Zhu, Y.; et al. PACT is a negative regulator of p53 and essential for cell growth and embryonic development. Proc. Natl. Acad. Sci. USA 2007, 104, 7951–7956. [Google Scholar] [CrossRef] [PubMed]
- Lane, D.P.; Verma, C. Mdm2 in Evolution. Genes Cancer 2012, 3, 3–4. [Google Scholar] [CrossRef] [PubMed]
- Huang, P.; Ma, X.; Zhao, Y.; Miao, L. The C. elegans homolog of RBBP6 (RBPL-1) regulates fertility through controlling cell proliferation in the germline and nutrient synthesis in the intestine. PLoS ONE 2013, 8, e58736. [Google Scholar] [CrossRef] [PubMed]
- Gao, S.; Scott, R.E. P2P-R protein overexpression restricts mitotic progression at prometaphase and promotes mitotic apoptosis. J. Cell. Physiol. 2002, 193, 199–207. [Google Scholar] [CrossRef] [PubMed]
- Miotto, B.; Chibi, M.; Xie, P.; Koundrioukoff, S.; Moolman-Smook, H.; Pugh, D.; Debatisse, M.; He, F.; Zhang, L.; Defossez, P.A. The RBBP6/ZBTB38/MCM10 axis regulates DNA replication and common fragile site stability. Cell Rep. 2014, 7, 575–587. [Google Scholar] [CrossRef] [PubMed]
- Hoh, J.; Jin, S.; Parrado, T.; Edington, J.; Levine, A.J.; Ott, J. The p53MH algorithm and its application in detecting p53-responsive genes. Proc. Natl. Acad. Sci. USA 2002, 99, 8467–8472. [Google Scholar] [CrossRef] [PubMed]
- Pignoni, F.; Steingrimsson, E.; Lengyel, J.A. bicoid and the terminal system activate tailless expression in the early Drosophila embryo. Development 1992, 115, 239–251. [Google Scholar] [PubMed]
- Mlodzik, M.; Gehring, W.J. Hierarchy of the genetic interactions that specify the anteroposterior segmentation pattern of the Drosophila embryo as monitored by caudal protein expression. Development 1987, 101, 421–435. [Google Scholar]
- Han, K.; Manley, J.L. Functional domains of the Drosophila Engrailed protein. EMBO J. 1993, 12, 2723–2733. [Google Scholar] [PubMed]
- Brunner, E.; Peter, O.; Schweizer, L.; Basler, K. Pangolin encodes a Lef-1 homologue that acts downstream of Armadillo to transduce the Wingless signal in Drosophila. Nature 1997, 385, 829–833. [Google Scholar] [CrossRef] [PubMed]
- England, B.P.; Admon, A.; Tjian, R. Cloning of Drosophila transcription factor Adf-1 reveals homology to Myb oncoproteins. Proc. Natl. Acad. Sci. USA 1992, 89, 683–687. [Google Scholar] [CrossRef] [PubMed]
- Luo, S.D.; Shi, G.W.; Baker, B.S. Direct targets of the D. melanogaster DSXF protein and the evolution of sexual development. Development 2011, 138, 2761–2771. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.; Gogos, J.; Kirsh, S.; Kafatos, F. Multiple zinc finger forms resulting from developmentally regulated alternative splicing of a transcription factor gene. Science 1992, 257, 1946–1950. [Google Scholar] [CrossRef] [PubMed]
- Hsu, T.; Bagni, C.; Sutherland, J.D.; Kafatos, F.C. The transcriptional factor CF2 is a mediator of EGF-R-activated dorsoventral patterning in Drosophila oogenesis. Genes Dev. 1996, 10, 1411–1421. [Google Scholar] [CrossRef] [PubMed]
- Matsukage, A.; Hirose, F.; Yoo, M.-A.; Yamaguchi, M. The DRE/DREF transcriptional regulatory system: A master key for cell proliferation. Biochim. Biophys. Acta Gene Regul. Mech. 2008, 1779, 81–89. [Google Scholar] [CrossRef]
- Baylies, M.K.; Bate, M. Twist: A myogenic switch in Drosophila. Science 1996, 272, 1481–1484. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Mani, S.A.; Donaher, J.L.; Ramaswamy, S.; Itzykson, R.A.; Come, C.; Savagner, P.; Gitelman, I.; Richardson, A.; Weinberg, R.A. Twist, a master regulator of morphogenesis, plays an essential role in tumor metastasis. Cell 2004, 117, 927–939. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, K.K.K.; Bryantsev, A.L.; Cripps, R.M. Myocyte enhancer factor 2 and chorion factor 2 collaborate in activation of the myogenic program in Drosophila. Mol. Cell. Biol. 2008, 28, 1616–1629. [Google Scholar] [CrossRef] [PubMed]
- Cripps, R.M.; Black, B.L.; Zhao, B.; Lien, C.-L.; Schulz, R.A.; Olson, E.N. The myogenic regulatory gene Mef2 is a direct target for transcriptional activation by Twist during Drosophila myogenesis. Genes Dev. 1998, 12, 422–434. [Google Scholar] [CrossRef] [PubMed]
- Jiang, N.; Emberly, E.; Cuvier, O.; Hart, C.M. Genome-wide mapping of boundary element-associated factor (BEAF) binding sites in Drosophila melanogaster links BEAF to transcription. Mol. Cell. Biol. 2009, 29, 3556–3568. [Google Scholar] [CrossRef] [PubMed]
- Gilbert, M.K.; Tan, Y.Y.; Hart, C.M. The Drosophila boundary element-associated factors BEAF-32A and BEAF-32B affect chromatin structure. Genetics 2006, 173, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Raff, J.W.; Kellum, R.; Alberts, B. The Drosophila GAGA transcription factor is associated with specific regions of heterochromatin throughout the cell cycle. EMBO J. 1994, 13, 5977–5983. [Google Scholar] [PubMed]
- Morris, D.R.; Geballe, A.P. Upstream open reading frames as regulators of mRNA translation. Mol. Cell. Biol. 2000, 20, 8635–8642. [Google Scholar] [CrossRef] [PubMed]
- Wilkie, G.S.; Dickson, K.S.; Gray, N.K. Regulation of mRNA translation by 5'- and 3'-UTR-binding factors. Trends Biochem. Sci. 2003, 28, 182–188. [Google Scholar] [CrossRef] [PubMed]
- Coll, O.; Villalba, A.; Bussotti, G.; Notredame, C.; Gebauer, F. A novel, noncanonical mechanism of cytoplasmic polyadenylation operates in Drosophila embryogenesis. Genes Dev. 2010, 24, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Blencowe, B.J.; Issner, R.; Kim, J.; McCaw, P.; Sharp, P.A. New proteins related to the Ser-Arg family of splicing factors. RNA 1995, 1, 852–865. [Google Scholar] [PubMed]
- Hart, C.M.; Cuvier, O.; Laemmli, U.K. Evidence for an antagonistic relationship between the boundary element-associated factor BEAF and the transcription factor DREF. Chromosoma 1999, 108, 375–383. [Google Scholar] [CrossRef] [PubMed]
- Hirose, F.; Ohshima, N.; Shiraki, M.; Inoue, Y.H.; Taguchi, O.; Nishi, Y.; Matsukage, A.; Yamaguchi, M. Ectopic expression of DREF induces DNA synthesis, apoptosis, and unusual morphogenesis in the Drosophila eye imaginal disc: Possible interaction with polycomb and trithorax group proteins. Mol. Cell. Biol. 2001, 21, 7231–7242. [Google Scholar] [CrossRef] [PubMed]
- Meinhart, A.; Cramer, P. Recognition of RNA polymerase II carboxy-terminal domain by 3(prime)-RNA-processing factors. Nature 2004, 430, 223–226. [Google Scholar] [CrossRef] [PubMed]
- Pauli, D.; Tonka, C.H.; Tissieres, A.; Arrigo, A.P. Tissue-specific expression of the heat shock protein HSP27 during Drosophila melanogaster development. J. Cell Biol. 1990, 111, 817–828. [Google Scholar] [CrossRef] [PubMed]
- Michaud, S.; Lavoie, S.; Guimond, M.-O.; Tanguay, R.M. The nuclear localization of Drosophila Hsp27 is dependent on a monopartite arginine-rich NLS and is uncoupled from its association to nuclear speckles. Biochim. Biophys. Acta Mol. Cell Res. 2008, 1783, 1200–1210. [Google Scholar] [CrossRef]
- Foe, V.E.; Odell, G.M.; Edgar, B.A. Mitosis and morphogenesis in the Drosophila embryo: Point and counterpoin. In The Development of Drosophila Melanogaster; Bate, M., Arias, A.M., Eds.; Cold Spring Harbor Laboratory Press: New York, NY, USA, 1993; pp. 149–300. [Google Scholar]
- De Oca Luna, R.M.; Wagner, D.S.; Lozano, G. Rescue of early embryonic lethality in mdm2-deficient mice by deletion of p53. Nature 1995, 378, 203–206. [Google Scholar] [CrossRef] [PubMed]
- Mach, J.M.; Lehmann, R. An Egalitarian-BicaudalD complex is essential for oocyte specification and axis determination in Drosophila. Genes Dev. 1997, 11, 423–435. [Google Scholar] [CrossRef] [PubMed]
- Bullock, S.L.; Ish-Horowicz, D. Conserved signals and machinery for RNA transport in Drosophila oogenesis and embryogenesis. Nature 2001, 414, 611–616. [Google Scholar] [CrossRef] [PubMed]
- Clark, K.A.; McKearin, D.M. The Drosophila stonewall gene encodes a putative transcription factor essential for germ cell development. Development 1996, 122, 937–950. [Google Scholar] [PubMed]
- Maines, J.Z.; Park, J.K.; Williams, M.; McKearin, D.M. Stonewalling Drosophila stem cell differentiation by epigenetic controls. Development 2007, 134, 1471–1479. [Google Scholar] [CrossRef] [PubMed]
- Manoli, D.S.; Meissner, G.W.; Baker, B.S. Blueprints for behavior: Genetic specification of neural circuitry for innate behaviors. Trends Neurosci. 2006, 29, 444–451. [Google Scholar] [CrossRef] [PubMed]
- Crozatier, M.; Kongsuwan, K.; Ferrer, P.; Merriam, J.R.; Lengyel, J.A.; Vincent, A. Single amino acid exchanges in separate domains of the Drosophila serendipity delta zinc finger protein cause embryonic and sex biased lethality. Genetics 1992, 131, 905–916. [Google Scholar] [PubMed]
- Tie, F.; Furuyama, T.; Prasad-Sinha, J.; Jane, E.; Harte, P.J. The Drosophila Polycomb Group proteins ESC and E(Z) are present in a complex containing the histone-binding protein p55 and the histone deacetylase RPD3. Development 2001, 128, 275–286. [Google Scholar] [PubMed]
- Bayer, E.A.; Ehrlich-Rogozinski, S.; Wilchek, M. Sodium dodecyl sulfate-polyacrylamide gel electrophoretic method for assessing the quaternary state and comparative thermostability of avidin and streptavidin. Electrophoresis 1996, 17, 1319–1324. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Gong, Q.; Xia, B.; Groves, B.; Zimmermann, M.; Mugler, C.; Mu, D.; Matsumoto, B.; Seaman, M.; Ma, D. A role of histone H3 lysine 4 methyltransferase components in endosomal trafficking. J. Cell Biol. 2009, 186, 343–353. [Google Scholar] [CrossRef] [PubMed]
- Gharahdaghi, F.; Weinberg, C.R.; Meagher, D.A.; Imai, B.S.; Mische, S.M. Mass spectrometric identification of proteins from silver-stained polyacrylamide gel: A method for the removal of silver ions to enhance sensitivity. Electrophoresis 1999, 20, 601–605. [Google Scholar] [CrossRef] [PubMed]
- Shevchenko, A.; Tomas, H.; Havlis, J.; Olsen, J.V.; Mann, M. In-gel digestion for mass spectrometric characterization of proteins and proteomes. Nat. Protoc. 2007, 1, 2856–2860. [Google Scholar] [CrossRef]
- White, R.A.H. Immunolabelling of drosophila. In Drosophila: A Practical Approach, 2nd ed.; Roberts, D.B., Ed.; Oxford University Press: New York, NY, USA, 1998; pp. 215–240. [Google Scholar]
- Verheyen, E.; Cooley, L. Looking at Oogenesis. In Drosophila Melanogaster: Practical Uses in Cell and Molecular Biology; Goldstein, L.S.B., Fyrberg, E.A., Eds.; Academic Press: San Diego, CA, USA, 1994; pp. 545–561. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hull, R.; Oosthuysen, B.; Cajee, U.-F.; Mokgohloa, L.; Nweke, E.; Antunes, R.J.; Coetzer, T.H.T.; Ntwasa, M. The Drosophila Retinoblastoma Binding Protein 6 Family Member Has Two Isoforms and Is Potentially Involved in Embryonic Patterning. Int. J. Mol. Sci. 2015, 16, 10242-10266. https://doi.org/10.3390/ijms160510242
Hull R, Oosthuysen B, Cajee U-F, Mokgohloa L, Nweke E, Antunes RJ, Coetzer THT, Ntwasa M. The Drosophila Retinoblastoma Binding Protein 6 Family Member Has Two Isoforms and Is Potentially Involved in Embryonic Patterning. International Journal of Molecular Sciences. 2015; 16(5):10242-10266. https://doi.org/10.3390/ijms160510242
Chicago/Turabian StyleHull, Rodney, Brent Oosthuysen, Umar-Faruq Cajee, Lehlogonolo Mokgohloa, Ekene Nweke, Ricardo Jorge Antunes, Theresa H. T. Coetzer, and Monde Ntwasa. 2015. "The Drosophila Retinoblastoma Binding Protein 6 Family Member Has Two Isoforms and Is Potentially Involved in Embryonic Patterning" International Journal of Molecular Sciences 16, no. 5: 10242-10266. https://doi.org/10.3390/ijms160510242