
Overexpressing of OsAMT1-3, a High Affinity Ammonium Transporter Gene, Modifies Rice Growth and Carbon-Nitrogen Metabolic Status

Abstract
:1. Introduction
2. Results
2.1. Accumulated AMT1-3 mRNA Transcripts in T0 Transgenic Plants


2.2. Effect of AMT1-3-Overexpressing on Growth Phenotype and Yield


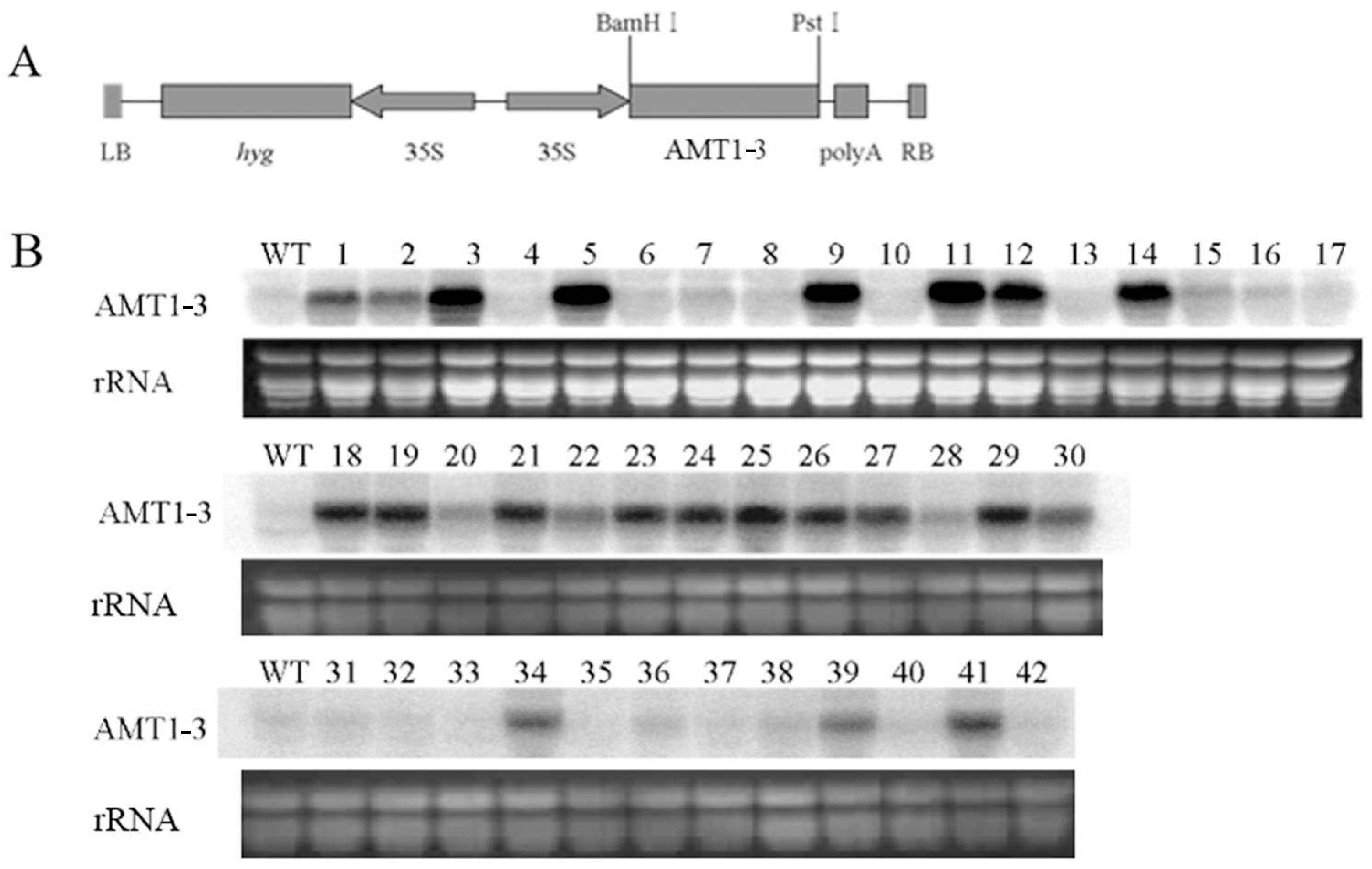{kind=link}
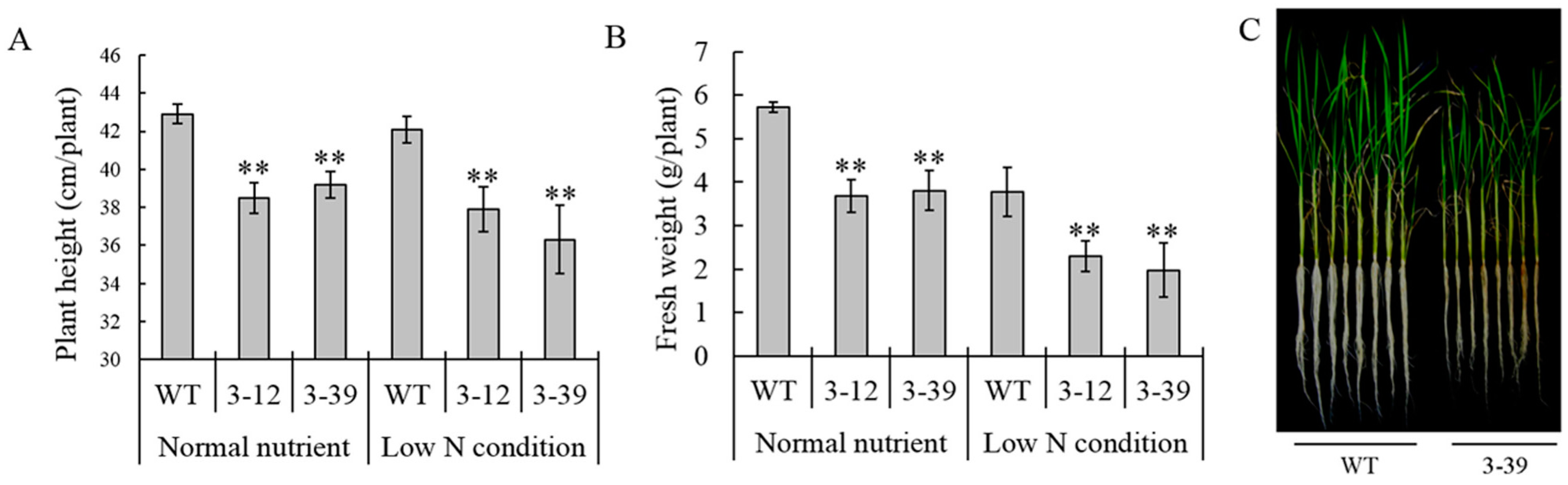{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | SPAD | Photosynthesis Parameters at Heading Stage | ||||
|---|---|---|---|---|---|---|
| Tillering Stage | Heading Stage | Photosynthetic Rate (μmol CO2·m−2·s−2) | Stomatal Conductance (mmol·m−2·s−1) | Intercellular CO2 Concentration (μL·L−1) | Transpiration Rate (mmol H2O·m−2·S−1) | |
| 0× N | ||||||
| WT | 34.3 ± 2.4 | 32.7 ± 1.7 | 13.24 ± 1.47 | 0.28 ± 0.04 | 280.48 ± 1.12 | 7.54 ± 0.65 |
| OX-AMT1-3 | 34.3 ± 2.5 | 33.0 ± 1.4 | 11.84 ± 2.34 | 0.25 ± 0.05 | 280.16 ± 8.55 | 6.71 ± 0.95 |
| 0.1× N | ||||||
| WT | 40.7 ± 2.0 | 40.7 ± 2.2 | 10.58 ± 1.64 | 0.29 ± 0.02 | 297.52 ± 10.16 | 7.24 ± 0.25 |
| OX-AMT1-3 | 42.3 ± 3.4 | 40.4 ± 1.2 | 12.88 ± 1.59 | 0.30 ± 0.05 | 287.51 ± 11.30 | 7.20 ± 0.71 |
| 1× N | ||||||
| WT | 43.6 ± 2.0 | 47.1 ± 0.5 | 16.07 ± 1.78 | 0.41 ± 0.04 | 290.97 ± 8.26 | 7.49 ± 0.47 |
| OX-AMT1-3 | 44.4 ± 1.3 | 46.5 ± 1.3 | 12.49 ± 1.76 * | 0.27 ± 0.09 ** | 278.65 ± 23.78 | 6.09 ± 1.47 |
| 5× N | ||||||
| WT | 44.4 ± 1.5 | 48.4 ± 1.3 | 19.12 ± 3.45 | 0.53 ± 0.05 | 305.62 ± 15.74 | 7.85 ± 0.17 |
| OX-AMT1-3 | 43.6 ± 4.0 | 47.4 ± 1.3 | 16.95 ± 2.39 | 0.48 ± 0.09 | 300.49 ± 14.01 | 7.86 ± 1.04 |
| Treatment | Panicals/Plant | Filled Grains/Panicle | Seed Rate (%) | Thousand Grains Weight (g) | Yield (g/Plant) |
|---|---|---|---|---|---|
| 0× N | |||||
| WT | 3.4 ± 0.3 | 19.1 ± 1.5 | 35.1 ± 1.5 | 24.46 ± 0.65 | 1.60 ± 0.11 |
| OX-AMT1-3 | 2.5 ± 0.4 * | 15.6 ± 2.6 | 33.7 ± 1.2 | 22.89 ± 0.94 | 0.87 ± 0.05 ** |
| 0.1× N | |||||
| WT | 4.0 ± 0.2 | 21.4 ± 3.5 | 36.4 ± 4.3 | 23.05 ± 0.74 | 2.00 ± 0.44 |
| OX-AMT1-3 | 3.5 ± 0.2 | 21.9 ± 3.8 | 33.6 ± 4.8 | 23.68 ± 0.91 | 1.83 ± 0.23 |
| 1× N | |||||
| WT | 11.5 ± 1.1 | 44.9 ± 4.9 | 66.9 ± 5.9 | 24.98 ± 0.21 | 12.78 ± 0.42 |
| OX-AMT1-3 | 11.6 ± 0.6 | 39.5 ± 4.3 | 64.8 ± 6.1 | 25.35 ± 0.31 | 11.56 ± 0.87 |
| 5× N | |||||
| WT | 14.6 ± 0.9 | 32.5 ± 2.3 | 65.9 ± 10.9 | 22.77 ± 0.75 | 10.80 ± 0.41 |
| OX-AMT1-3 | 15.6 ± 0.7 | 23.3 ± 0.6 ** | 54.0 ± 4.4 | 21.87 ± 0.91 | 7.95 ± 0.26 ** |
2.3. Effect of AMT1-3-Overexpressing on Nitrogen Uptake and Accumulation

| Treatment | C% | N% | C/N | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Root | Stem | Leaf | Root | Stem | Leaf | Root | Stem | Leaf | |
| 0× N | |||||||||
| WT | 38.54 ± 0.60 | 38.26 ± 0.43 | 40.10 ± 0.20 | 1.76 ± 0.06 | 0.96 ± 0.04 | 2.14 ± 0.03 | 21.85 ± 0.77 | 39.79 ± 1.05 | 18.71 ± 0.25 |
| OX-AMT1-3 | 36.36 ± ND | 36.07 ± 1.63 | 38.25 ± 0.39 ** | 1.62 ± ND | 1.02 ± 0.03 | 2.04 ± 0.09 | 22.39 ± ND | 35.26 ± 0.61 ** | 18.80 ± 0.67 |
| 0.1× N | |||||||||
| WT | 38.47 ± 0.07 | 38.88 ± 0.10 | 40.52 ± 0.16 | 2.10 ± 0.14 | 1.40 ± 0.11 | 2.91 ± 0.18 | 18.34 ± 1.17 | 27.80 ± 2.20 | 13.94 ± 0.81 |
| OX-AMT1-3 | 39.13 ± 0.76 | 37.53 ± 0.39 | 36.51 ± 0.35 ** | 2.13 ± ND | 1.39 ± 0.07 | 2.52 ± 0.15 * | 17.17 ± ND | 26.97 ± 1.59 | 15.57 ± 0.85 * |
| 1× N | |||||||||
| WT | 37.48 ± 0.60 | 37.00 ± 0.36 | 41.46 ± 0.25 | 2.72 ± 0.004 | 2.44 ± 0.08 | 3.75 ± 0.08 | 13.77 ± 0.20 | 15.19 ± 0.49 | 11.05 ± 0.16 |
| OX-AMT1-3 | ND | 36.21 ± 0.46 | 39.01 ± 0.13 ** | ND | 2.37 ± 0.21 | 3.17 ± 0.14 ** | ND | 15.36 ± 1.24 | 12.33 ± 0.59 * |
| 5× N | |||||||||
| WT | 37.69 ± 0.61 | 36.96 ± 0.18 | 41.46 ± 0.25 | 2.85 ± 0.16 | 3.04 ± 0.08 | 3.98 ± 0.08 | 13.27 ± 1.00 | 12.15 ± 0.31 | 10.41 ± 0.15 |
| OX-AMT1-3 | 37.21 ± ND | 36.08 ± 0.78 | 39.04 ± 0.72 ** | 2.56 ± ND | 2.96 ± 0.02 | 3.43 ± 0.23 * | 14.52 ± ND | 12.18 ± 0.21 | 11.39 ± 0.60 * |
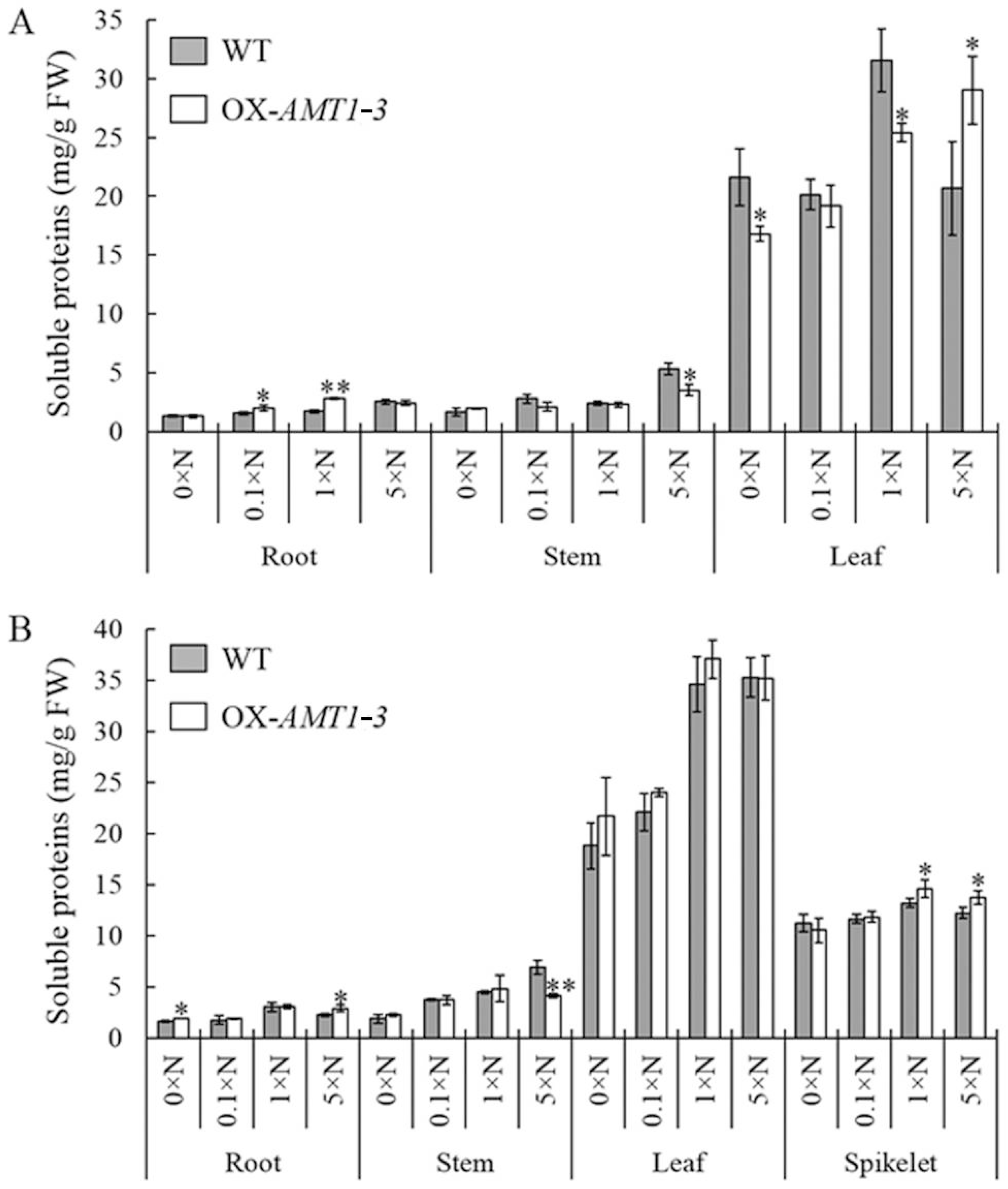
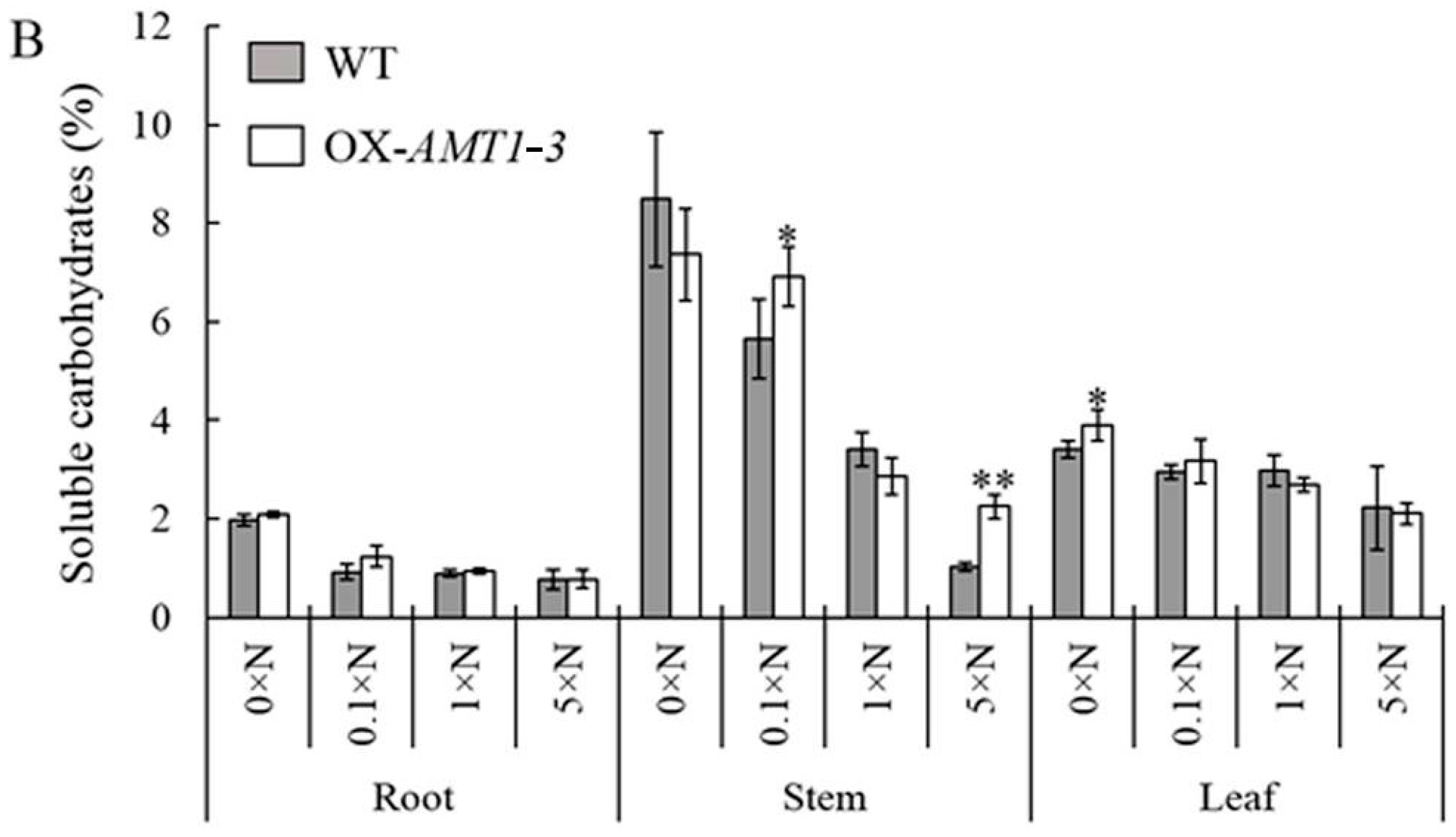
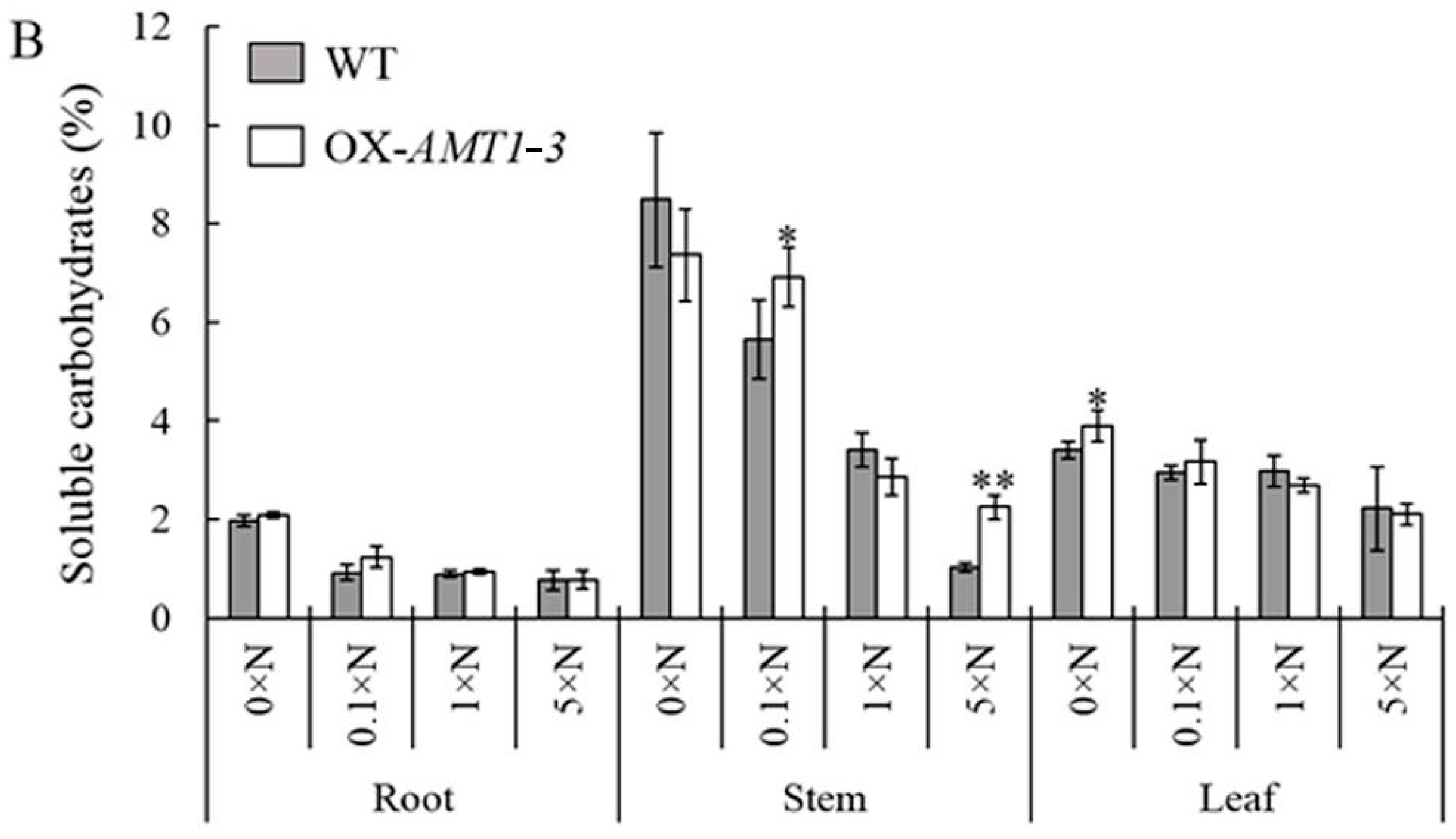
2.4. Effect of AMT1-3-Overexpressing on Soluble Proteins and Carbohydrates Concentrations



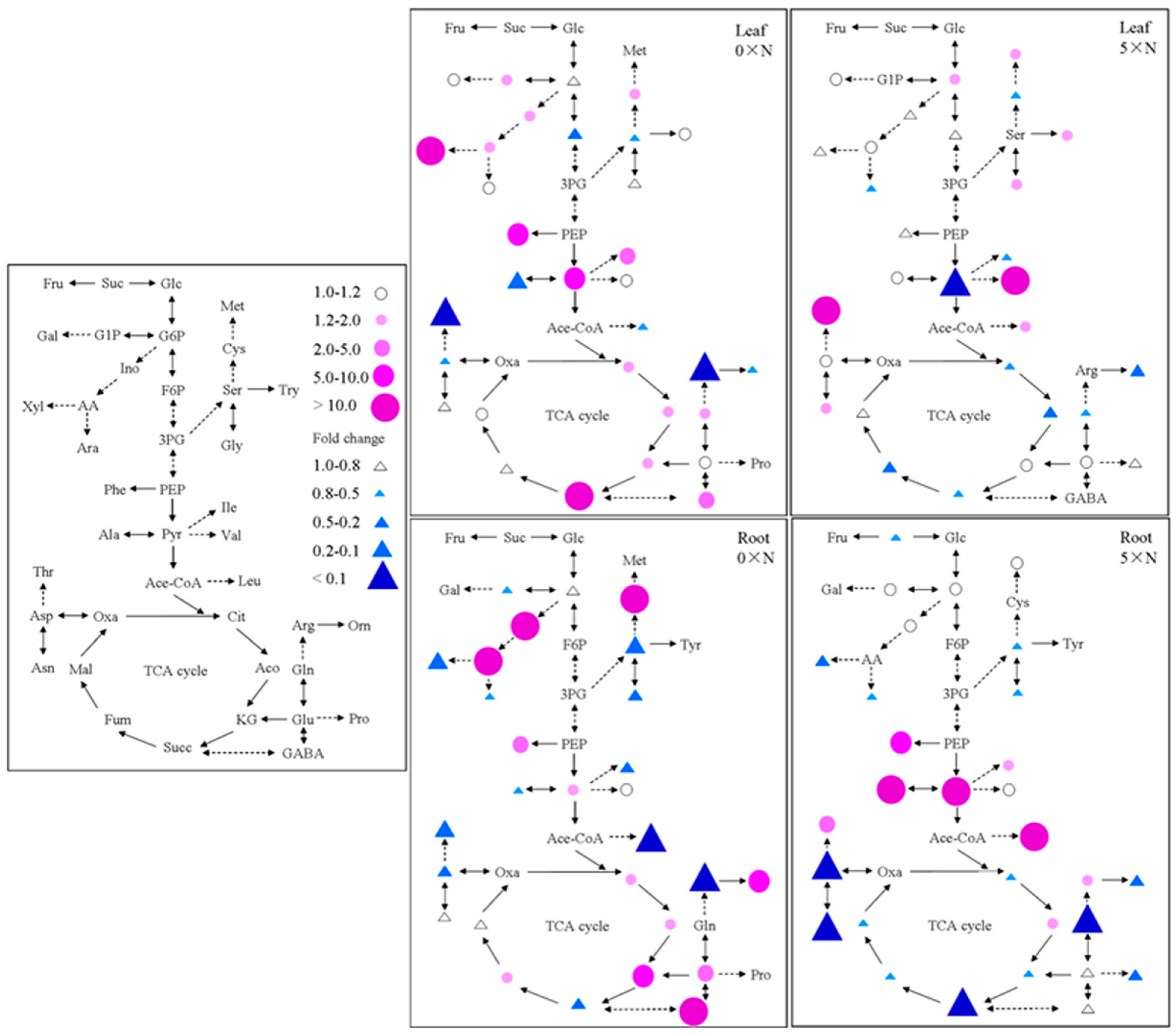
2.5. Effect of AMT1-3-Overexpressing on Carbon and Nitrogen Metabolites

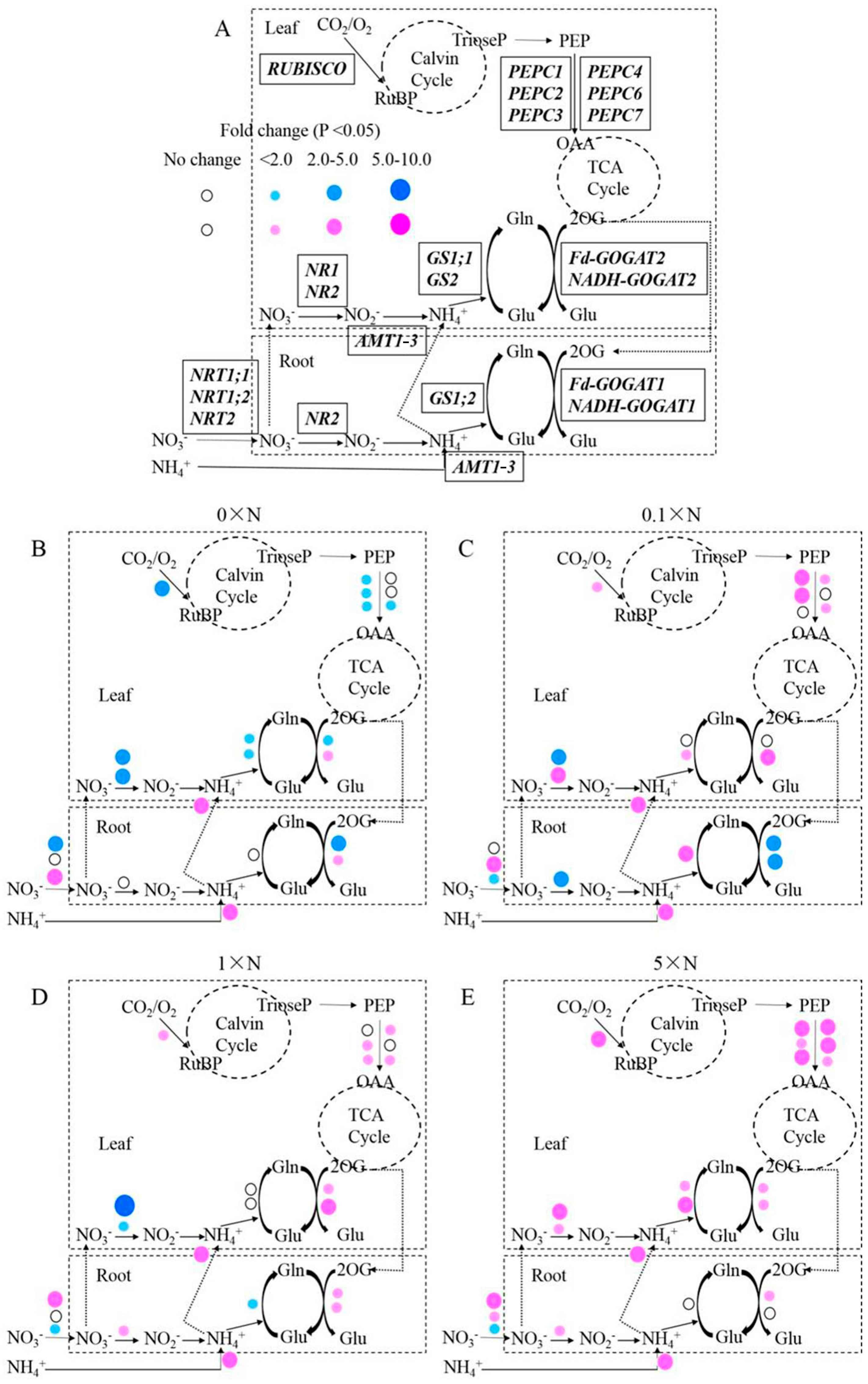
2.6. Effect of AMT1-3-Overexpressing on Gene Expression Level

3. Discussion
4. Materials and Methods
4.1. Constructs and Transformation
4.2. Plant Growth Conditions
4.3. Northern Blot, Southern Blot and Gene Expression Analysis
4.4. Nitrogen Uptake Assay
4.5. Physiological Parameters Determination
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lam, H.M.; Coschigano, K.T.; Oliveira, I.C.; Melo-Oliveira, R.; Coruzzi, G.M. The molecular-genetics of nitrogen assimilation into amino acids in higher plants. Ann. Rev. Plant Physiol. Mol. Biol. 1996, 47, 569–593. [Google Scholar] [CrossRef]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef]
- Frink, C.R.; Waggoner, P.E.; Ausubel, J.H. Nitrogen fertilizer: Retrospect and prospect. Proc. Natl. Acad. Sci. USA 1999, 96, 1175–1180. [Google Scholar] [CrossRef] [PubMed]
- Crawford, N.M.; Forde, B.G. Molecular and developmental biology of inorganic nitrogen nutrition. In The Arabidopsis Book; Meyerowitz, E., Somerville, C., Eds.; American Society of Plant Biologists: Rockville, MD, USA, 2002. [Google Scholar]
- Howitt, S.M.; Udvardi, M.K. Structure, function and regulation of ammonium transporters in plants. Biochim. Biophys. Acta 2000, 1465, 152–170. [Google Scholar] [CrossRef]
- Bloom, A.J.; Sukrapanna, S.S.; Warner, R.L. Root respiration associated with ammonium and nitrate absorption and assimilation by barley. Plant Physiol. 1992, 99, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Gazzarrini, S.; Lejay, L.; Gojon, A.; Ninnemann, O.; Frommer, W.B.; von Wirén, N. Three functional transporters for constitutive, diurnally regulated, and starvation-induced uptake of ammonium into Arabidopsis roots. Plant Cell 1999, 11, 937–948. [Google Scholar] [CrossRef] [PubMed]
- Coruzzi, G.M.; Zhou, L. Carbon and nitrogen sensing and signaling in plants: Emerging “matrix effects”. Curr. Opin. Plant Biol. 2001, 4, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Martin, T.; Oswald, O.; Graham, I.A. Arabidopsis seedling growth, storage lipid mobilization and photosynthetic gene expression are regulated by carbon: Nitrogen availability. Plant Physiol. 2002, 128, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A.; Truong, H.N. Regulation of C/N interaction in model plant species. In Enhancing the Efficiency of Nitrogen Utilization in Plants; Goyal, S., Tischner, R., Basra, A., Eds.; Haworth Press: New York, NY, USA, 2005; pp. 127–173. [Google Scholar]
- Zheng, Z. Carbon and nitrogen nutrient balance signaling in plants. Plant Signal. Behav. 2009, 4, 584–591. [Google Scholar] [CrossRef] [PubMed]
- Nunes-Nesi, A.; Fernie, A.R.; Stitt, M. Metabolic and signaling aspects underpinning the regulation of plant carbon nitrogen interactions. Mol. Plant 2010, 3, 973–996. [Google Scholar] [CrossRef] [PubMed]
- Loqué, D.; von Wirén, N. Regulatory levels for the transport of ammonium in plant roots. J. Exp. Bot. 2004, 55, 1293–1305. [Google Scholar] [CrossRef] [PubMed]
- Gaur, V.S.; Singh, U.S.; Gupta, A.K.; Kumar, A. Understanding the differential nitrogen sensing mechanism in rice genotypes through expression analysis of high and low affinity ammonium transporter genes. Mol. Biol. Rep. 2012, 39, 2233–2241. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.Y.; Glass, A.; Shaff, J.E.; Kochian, L.V. Ammonium uptake by rice roots. Plant Physiol. 1994, 104, 899–906. [Google Scholar] [PubMed]
- Ludewig, U.; Wilken, S.; Wu, B.; Jost, W.; Obrdlik, P.; El Bakkoury, M.; Marini, A.-M.; André, B.; Hamacher, T.; Boles, E.; et al. Homo- and hetero-oligomerization of ammonium transporter-1 NH4+ uniporters. J. Biol. Chem. 2003, 278, 45603–45610. [Google Scholar]
- Bakouh, N.; Benjelloun, F.; Hulin, P.; Brouillard, F.; Edelman, A.; Chérif-Zahar, B.; Planelles, G. NH3 is involved in the NH4+ transport induced by the functional expression of the human RhC glycoprotein. J. Biol. Biochem. 2004, 279, 15975–15983. [Google Scholar]
- Khademi, S.; O’Connell, J.; Remis, J.; Robles-Colmenares, Y.; Miercke, L.J.; Stroud, R.M. Mechanism of ammonia transport by Amt/MEP/Rh: Structure of AmtB. Science 2004, 305, 1587–1594. [Google Scholar] [CrossRef]
- Ripoche, P.; Bertrand, O.; Gane, P.; Birkenmeier, C.; Colin, Y.; Cartron, J.-P. Human Rhesusassociated glycoprotein mediates facilitated transport of NH3 into red blood cells. Proc. Nat. Acad. Sci. USA 2004, 101, 17222–17227. [Google Scholar] [CrossRef] [PubMed]
- Zheng, L.; Kostrewa, D.; Bernèche, S.; Winkler, F.K.; Li, X.-D. The mechanism of ammonia transport based on the crystal structure of AmtB of Escherichia coli. Proc. Natl. Acad. Sci. USA 2004, 101, 17090–17095. [Google Scholar] [CrossRef] [PubMed]
- Benjelloun, F.; Bakouh, N.; Fritsch, J.; Hulin, P.; Lipecka, J.; Edelman, A.; Planelles, G.; Thomas, S.R.; Chérif-Zahar, B. Expression of the human erythroid Rh glycoprotein (RhAG) enhances both NH3 and NH4+ transport in HeLa cells. Pflüg. Arch. 2005, 450, 155–167. [Google Scholar] [CrossRef]
- Mak, D.-O.D.; Dang, B.; Weiner, I.D.; Foskett, J.K.; Westhoff, C.M. Characterization of ammonia transport by the kidney Rh glycoproteins RhBG and RhCG. Am. J. Physiol. 2006, 290, F297–F305. [Google Scholar]
- Mayer, M.; Dynowski, M.; Ludewig, U. Ammonium ion transport by the AMT/Rh homologue LeAMT1;1. Biochem. J. 2006, 396, 431–437. [Google Scholar] [CrossRef] [PubMed]
- Fong, R.N.; Kim, K.S.; Yoshihara, C.; Inwood, W.B.; Kustu, S. The W148L substitution in the Escheria coli ammonium channel AmtB increase flux and indicates that the substrate is an ion. Proc. Natl. Acad. Sci. USA 2007, 104, 18706–18711. [Google Scholar] [CrossRef] [PubMed]
- Ortiz-Ramirez, C.; Mora, S.I.; Trejo, J.; Pantoja, O. PvAMT1;1, a highly selective ammonium transporter that functions as H+/NH4+ symporter. J. Biol. Chem. 2011, 286, 31113–31122. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.M.; André, B. In vivo N-glycosylation of the mep2 high-affinity ammonium transporter of Saccharomyces cerevisiae reveals an extracytosolic N-terminus. Mol. Microbiol. 2000, 38, 552–564. [Google Scholar] [CrossRef] [PubMed]
- Thomas, G.H.; Mullins, J.G.; Merrick, M. Membrane topology of the Mep/Amt family of ammonium transporters. Mol. Microbiol. 2000, 37, 331–344. [Google Scholar] [CrossRef] [PubMed]
- Litman, T.; Søgaard, R.; Zeuthen, T. Ammonia and urea permeability of mammalian aquaporins. In Handbook of Experimental Pharmacology; Springer: Berlin, Germany, 2008; pp. 327–358. [Google Scholar]
- Yuan, L.; Graff, L.; Loqué, D.; Kojima, S.; Tsuchiya, Y.N.; Takahashi, H.; von Wirén, N. AtAMT1;4, a pollen-specific high-affinity ammonium transporter of the plasma membrane in Arabidopsis. Plant Cell Physiol. 2009, 50, 13–25. [Google Scholar] [CrossRef] [PubMed]
- Loqué, D.; Yuan, L.; Kojima, S.; Gojon, A.; Wirth, J.; Gazzarrini, S.; Ishiyama, K.; Takahashi, H.; von Wirén, N. Additive contribution of AMT1;1 and AMT1;3 to high-affinity ammonium uptake across the plasma membrane of nitrogen-deficient Arabidopsis roots. Plant J. 2006, 48, 522–534. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Loqué, D.; Kojima, S.; Rauch, S.; Ishiyama, K.; Inoue, E.; Takahashi, H.; von Wirén, N. The organization of high-affinity ammonium uptake in Arabidopsis roots depends on the spatial arrangement and biochemical properties of AMT1-type transporters. Plant Cell 2007, 19, 2636–2652. [Google Scholar] [CrossRef] [PubMed]
- Engineer, C.B.; Kranz, R.G. Reciprocal leaf and root expression of AtAmt1.1 and root architectural changes in response to nitrogen starvation. Plant Physiol. 2007, 143, 236–250. [Google Scholar]
- Sohlenkamp, C.; Shelden, M.; Howitt, S.; Udvardi, M. Characterization of Arabidopsis AtAMT2, a novel ammonium transporter in plants. FEBS Lett. 2000, 467, 273–278. [Google Scholar] [CrossRef] [PubMed]
- Sohlenkamp, C.; Wood, C.C.; Roeb, G.W.; Udvardi, M.T. Characterization of Arabidopsis AtAMT2, a high-affinity ammonium transporter of the plasma membrane. Plant Physiol. 2002, 130, 1788–1796. [Google Scholar] [CrossRef] [PubMed]
- Sonoda, Y.; Ikeda, A.; Saiki, S.; von Wirén, N.; Yamaya, T.; Yamaguchi, J. Distinct expression and function of three ammonium transporter genes (OsAMT1;1–1;3) in rice. Plant Cell Physiol. 2003, 44, 726–734. [Google Scholar] [CrossRef] [PubMed]
- Marini, A.M.; Soussi-Boudekou, S.; Vissers, S.; André, B. A family of ammonium transporters in Saccharomyces cerevisiae. Mol. Cell. Biol. 1997, 17, 4282–4293. [Google Scholar] [PubMed]
- Suenaga, A.; Moriya, K.; Sonoda, Y.; Ikeda, A.; von Wirén, N.; Hayakawa, T.; Yamaguchi, J.; Yamaya, T. Constitutive expression of a novel-type ammonium transporter OsAMT2 in rice plants. Plant Cell Physiol. 2003, 44, 206–211. [Google Scholar] [CrossRef] [PubMed]
- Pearson, C.J.; Finnegan, P.M.; Schjoerring, J.K. Regulation of the high-affinity ammonium transporter (BnAMT1;2) in the leaves of Brassica napus by nitrogen status. Plant Mol. Biol. 2002, 49, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Salvemini, F.; Marini, A.M.; Riccio, A.; Patriarca, E.J.; Chiurazzi, M. Functional characterization of an ammonium transporter gene from Lotus japonicus. Gene 2001, 270, 237–243. [Google Scholar] [CrossRef] [PubMed]
- Simon-Rosin, U.; Wood, C.; Udvardi, M.K. Molecular and cellular characterization of LjAMT2;1, ammonium transporter from the model legume Lotus japonicus. Plant Mol. Biol. 2003, 51, 99–108. [Google Scholar] [CrossRef] [PubMed]
- Lauter, F.R.; Ninnemann, O.; Bucher, M.; Riesmeier, J.W.; Frommer, W.B. Preferential expression of an ammonium transporter and of two putative nitrate transporters in root hairs of tomato. Proc. Natl. Acad. Sci. USA 1996, 93, 8139–8144. [Google Scholar] [CrossRef] [PubMed]
- Von WireÂn, N.; Gazzarrini, S.; Gojon, A.; Frommer, W.B. The molecular physiology of ammonium uptake and retrieval. Curr. Opin. Plant Biol. 2000, 3, 254–261. [Google Scholar] [CrossRef] [PubMed]
- Becker, D.; Stahnke, R.; Fendrick, I.; Frommer, W.B.; Vanderleyden, J.; Kaiser, W.M.; Hedrich, R. Expression of the ammonium transporter gene LeAMT1;2 is induced in tomato roots upon association with N2 fixing bacteria. Planta 2002, 215, 424–429. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Loqué, D.; Ye, F.; Frommer, W.B.; von Wirén, N. Nitrogen-dependent posttranscriptional regulation of the ammonium transporter AtAMT1;1. Plant Physiol. 2007, 143, 732–744. [Google Scholar] [CrossRef] [PubMed]
- Loqué, D.; Lalonde, S.; Looger, L.L.; von Wirén, N.; Frommer, W.B. A cytosolic trans-activation domain essential for ammonium uptake. Nature 2007, 446, 195–198. [Google Scholar] [CrossRef] [PubMed]
- Loqué, D.; Mora, S.I.; Andrade, S.L.A.; Pantoja, O.; Frommer, W.B. Pore mutations in ammonium transporter AMT1 with increased electrogenic ammonium transport activity. J. Biol. Chem. 2009, 284, 24988–24995. [Google Scholar] [CrossRef] [PubMed]
- Yuan, L.; Gu, R.; Xuan, Y.; Smith-Valle, E.; Loqué, D.; Frommer, W.B.; von Wirén, N. Allosteric regulation of transport activity by heterotrimerization of Arabidopsis ammonium transporter complexes in vivo. Plant Cell 2013, 25, 974–984. [Google Scholar] [CrossRef] [PubMed]
- Neuhäuser, B.; Dynowski, M.; Mayer, M.; Ludewig, U. Regulation of NH4+ transport by essential cross talk between AMT monomers through the carboxyl tails. Plant Physiol. 2007, 143, 1651–1659. [Google Scholar] [CrossRef] [PubMed]
- Lanquar, V.; Loqué, D.; Hörmann, F.; Yuan, L.; Bohner, A.; Engelsberger, W.R.; Lalonde, S.; Schulze, W.X.; vonWirén, N.; Frommer, W.B. Feedback inhibition of ammonium uptake by a phosphodependent allosteric mechanism in Arabidopsis. Plant Cell 2009, 21, 3610–3622. [Google Scholar] [CrossRef] [PubMed]
- Graff, L.; Obrdlik, P.; Yuan, L.; Loqué, D.; Frommer, W.B.; von Wirén, N. N-terminal cysteines affect oligomer stability of the allosterically regulated ammonium transporter LeAMT1;1. J. Exp. Bot. 2011, 62, 1361–1373. [Google Scholar] [CrossRef] [PubMed]
- Socolow, R. Nitrogen management and the future of food: Lessons from the management of energy and carbon. Proc. Natl. Acad. Sci. USA 1999, 96, 6001–6008. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Cui, Z.; Fan, M.; Zhang, W.; Chen, X.; Jiang, R. Integrated soil–crop system management: Reducing environmental risk while increasing crop pro-ductivity and improving nutrient use efficiency in China. J. Environ. Qual. 2011, 40, 1051–1057. [Google Scholar] [CrossRef] [PubMed]
- Hoque, M.S.; Masle, J.; Udvardi, M.K.; Ryan, P.R.; Upadhyaya, N.M. Over-expression of the rice OsAMT1-1 gene increases ammonium uptake and content, but impairs growth and development of plants under high ammonium nutrition. Funct. Plant Biol. 2006, 33, 153–163. [Google Scholar] [CrossRef]
- Kumar, A.; Kaiser, B.N.; Siddiqi, M.Y.; Glass, A.D.M. Functional characterization of OsAMT 1.1 overexpression lines of rice, Oryza sativa. Funct. Plant Biol. 2006, 33, 339–346. [Google Scholar] [CrossRef]
- Ranathunge, K.; EI-Kereamy, A.; Gidda, S.; Bi, Y.-M. OsAMT1;1 transgenic rice plants with enhanced NH4+ permeability show superior growth and higher yield under optimal and suboptimal NH4+ conditions. J. Exp. Bot. 2014, 65, 965–979. [Google Scholar] [CrossRef] [PubMed]
- Krapp, A.; Ferrario-Méry, S.; Touraine, B. Nitrogen and signaling. In Advances in Photosynthesis; Foyer, C., Noctor, G., Eds.; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2002. [Google Scholar]
- Lin, C.M.; Koh, S.; Stacey, G.; Yu, S.-M.; Lin, T.-Y.; Tsay, Y.F. Cloning and functional characterization of a constitutively expressed nitrate transporter gene, OsNRT1, from rice. Plant. Physiol. 2000, 122, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Araki, R.; Hasegawa, H. Expression of rice (Oryza sativa L.) genes involved in high-affinity nitrate transport during the period of nitrate induction. Breed. Sci. 2006, 56, 295–302. [Google Scholar]
- Lea, P.J.; Morot-Gaudry, J.F. Plant Nitrogen; Springer-Verlag: New York, NY, USA, 2001. [Google Scholar]
- Coruzzi, G.; Bush, D.R. Nitrogen and carbon nutrient and metabolite signaling in plants. Plant Physiol. 2001, 125, 61–64. [Google Scholar] [CrossRef] [PubMed]
- Foyer, C.H.; Parry, M.; Noctor, G. Markers and signals associated with nitrogen assimilation in higher plants. J. Exp. Bot. 2003, 54, 585–593. [Google Scholar] [CrossRef] [PubMed]
- Scheible, W.R.; GonzaÂles-Fontes, A.; Lauerer, M.; MuÈller-RoÈber, B.; Caboche, M.; Stitt, M. Nitrate acts as a signal to induce organic acid metabolism and repress starch metabolism in tobacco. Plant Cell 1997, 9, 783–798. [Google Scholar] [CrossRef] [PubMed]
- Dzuibany, C.; Haupt, S.; Fock, H.; Biehler, K.; Migge, A.; Becker, T. Regulation of nitrate reductase transcript level by glutamine accumulating in the leaves of a ferredoxin-dependent glutamate synthase-deficient gluS mutant of Arabidopsis thaliana, and by glutamine provided via the roots. Planta 1998, 20, 515–522. [Google Scholar] [CrossRef]
- Rawat, S.R.; Silim, S.N.; Kronzucker, H.J.; Siddiqi, M.Y.; Glass, A.D.M. AtAMT1gene expression and NH4+ uptake in roots of Arabidopsis thaliana: Evidence for regulation by root glutamine levels. Plant J. 1999, 19, 143–152. [Google Scholar] [CrossRef] [PubMed]
- Ferrario-MeÂry, S.; Masclaux, C.; Suzuki, A.; Valadier, M.H.; Hirel, B.; Foyer, C.H. Glutamine and a-ketoglutarate are metabolite signals involved in nitrate reductase gene transcription in untransformed and transformed tobacco plants deficient in ferredoxin-glutamine-a-ketoglutarate aminotransferase. Planta 2001, 213, 265–271. [Google Scholar] [CrossRef] [PubMed]
- Tremblay, P.L.; Hallenbeck, P.C. Of blood, brains and bacteria, the Amt/Rh transporter family: Emerging role of Amt as a unique microbial sensor. Mol. Microbiol. 2009, 71, 12–22. [Google Scholar] [CrossRef] [PubMed]
- Lima, J.E.; Kojima, S.; Takahashi, H.; von Wirén, N. Ammonium triggers lateral root branching in Arabidopsis in an AMMONIUM TRANSPORTER1;3—Dependent manner. Plant Cell 2010, 22, 3621–3633. [Google Scholar] [CrossRef] [PubMed]
- Hachiya, T.; Watanabe, C.K.; Fujimoto, M.; Ishikawa, T.; Takahara, K.; Kawai-Yamada, M.; Uchimiya, H.; Uesono, Y.; Terashima, I.; Noguchi, K. Nitrate addition alleviates ammonium toxicity without lessening ammonium accumulation, organic acid depletion and inorganic cation depletion in Arabidopsis thaliana shoots. Plant Cell Physiol. 2012, 53, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Sambrook, J.; Fritsch, E.F.; Maniatis, T. Molecular Cloning: A Laboratory Manual, 2nd ed.; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1989. [Google Scholar]
- Maniatis, T.A.; Fritsch, E.F.; Sambrook, J. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 1992. [Google Scholar]
- Hiei, Y.; Ohta, S.; Komari, T.; Kumashiro, T. Efficient transformation of rice (Oryza sativa L.) mediated by Agrobacterium and sequence analysis of the boundaries of the T-DNA. Plant J. 1994, 6, 271–282. [Google Scholar]
- Yoshida, S.; Forno, D.A.; Cook, J.H.; Gomez, K.A. Laboratory Manual for Physiological Studies of Rice, 3rd eds; International Rice Research Institute: Manila, Philippine, 1976. [Google Scholar]
- Melo, P.M.; Lima, L.M.; Santos, I.M.; Carvalho, H.G.; Cullimore, J.V. Expression of the plastid-located glutamine synthetase of Medicago truncatula: Accumulation of the precursor in root nodules reveals an in vivo control at the level of protein import into plastids. Plant Physiol. 2003, 132, 390–399. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef] [PubMed]
- Morris, D.L. Quantitative determination of carbohydrates with Dreywood’s anthrone reagent. Science 1948, 107, 254–255. [Google Scholar] [CrossRef] [PubMed]
- Maness, N. Extraction and analysis of soluble carbohydrates. Methods Mol. Biol. 2010, 639, 341–370. [Google Scholar] [PubMed]
- Kusano, M.; Fukushima, A.; Arita, M.; Jonsson, P.; Moritz, T.; Kobayashi, M.; Hayashi, N.; Tohge, T.; Saito, K. Unbiased characterization of genotype-dependent metabolic regulations by metabolomic approach in Arabidopsis thaliana. BMC Syst. Biol. 2007, 1, 53. [Google Scholar] [CrossRef] [PubMed]
- Kusano, M.; Fukushima, A.; Kobayashi, M.; Hayashi, N.; Jonsson, P.; Moritz, T.; Ebana, K.; Saito, K. Application of a metabolomic method combining one-dimensional and two-dimensional gas chromatographytime-of-flight/mass spectrometry to metabolic phenotyping of natural variants in rice. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 2007, 855, 71–79. [Google Scholar] [CrossRef]
- Redestig, H.; Fukushima, A.; Stenlund, H.; Moritz, T.; Arita, M.; Saito, K.; Kusano, M. Compensation for systematic cross-contribution improves normalization of mass spectrometry based metabolomics data. Anal. Chem. 2009, 81, 7974–7980. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bao, A.; Liang, Z.; Zhao, Z.; Cai, H. Overexpressing of OsAMT1-3, a High Affinity Ammonium Transporter Gene, Modifies Rice Growth and Carbon-Nitrogen Metabolic Status. Int. J. Mol. Sci. 2015, 16, 9037-9063. https://doi.org/10.3390/ijms16059037
Bao A, Liang Z, Zhao Z, Cai H. Overexpressing of OsAMT1-3, a High Affinity Ammonium Transporter Gene, Modifies Rice Growth and Carbon-Nitrogen Metabolic Status. International Journal of Molecular Sciences. 2015; 16(5):9037-9063. https://doi.org/10.3390/ijms16059037
Chicago/Turabian StyleBao, Aili, Zhijun Liang, Zhuqing Zhao, and Hongmei Cai. 2015. "Overexpressing of OsAMT1-3, a High Affinity Ammonium Transporter Gene, Modifies Rice Growth and Carbon-Nitrogen Metabolic Status" International Journal of Molecular Sciences 16, no. 5: 9037-9063. https://doi.org/10.3390/ijms16059037