
Structural Insights into tRNA Dynamics on the Ribosome

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction


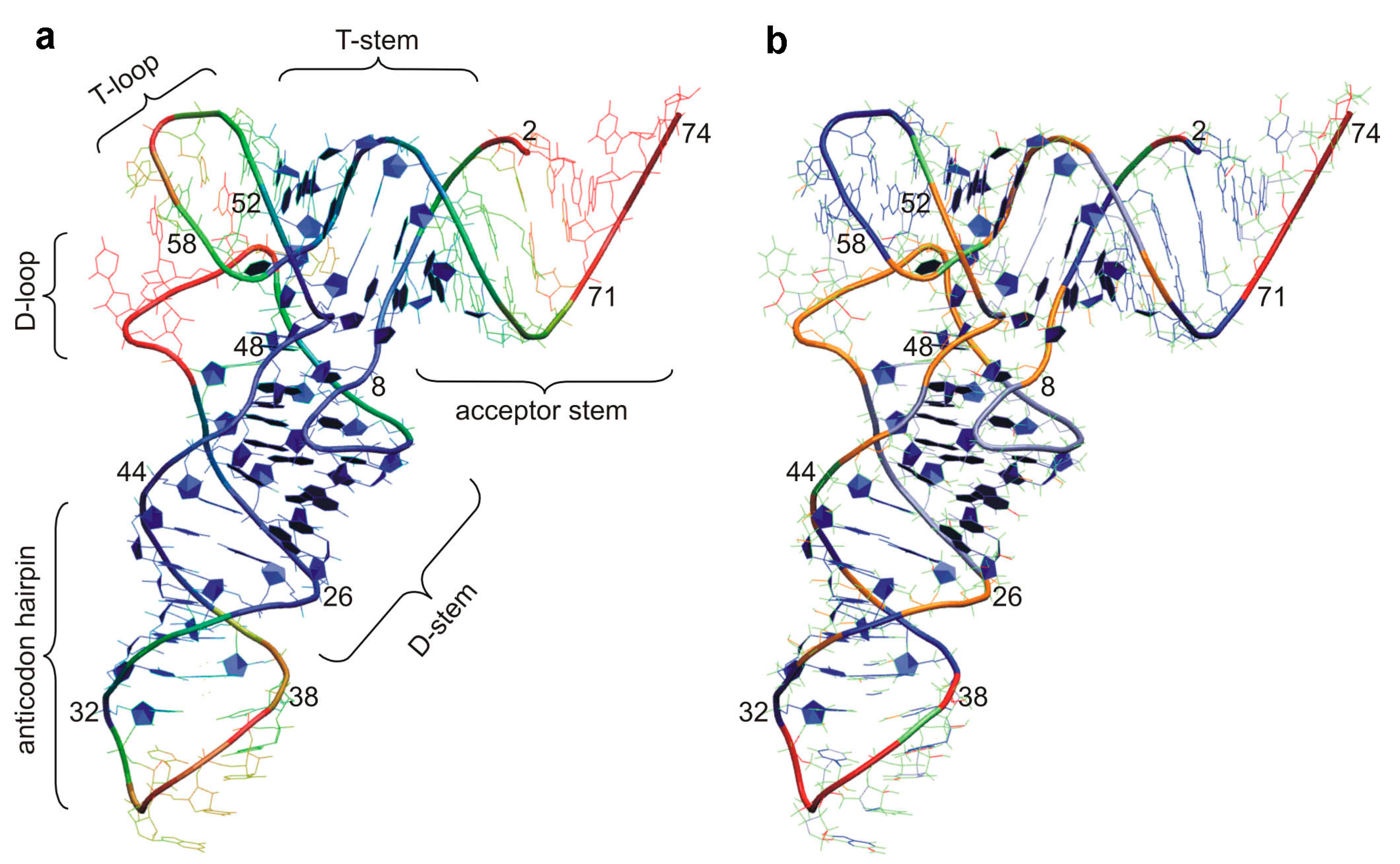
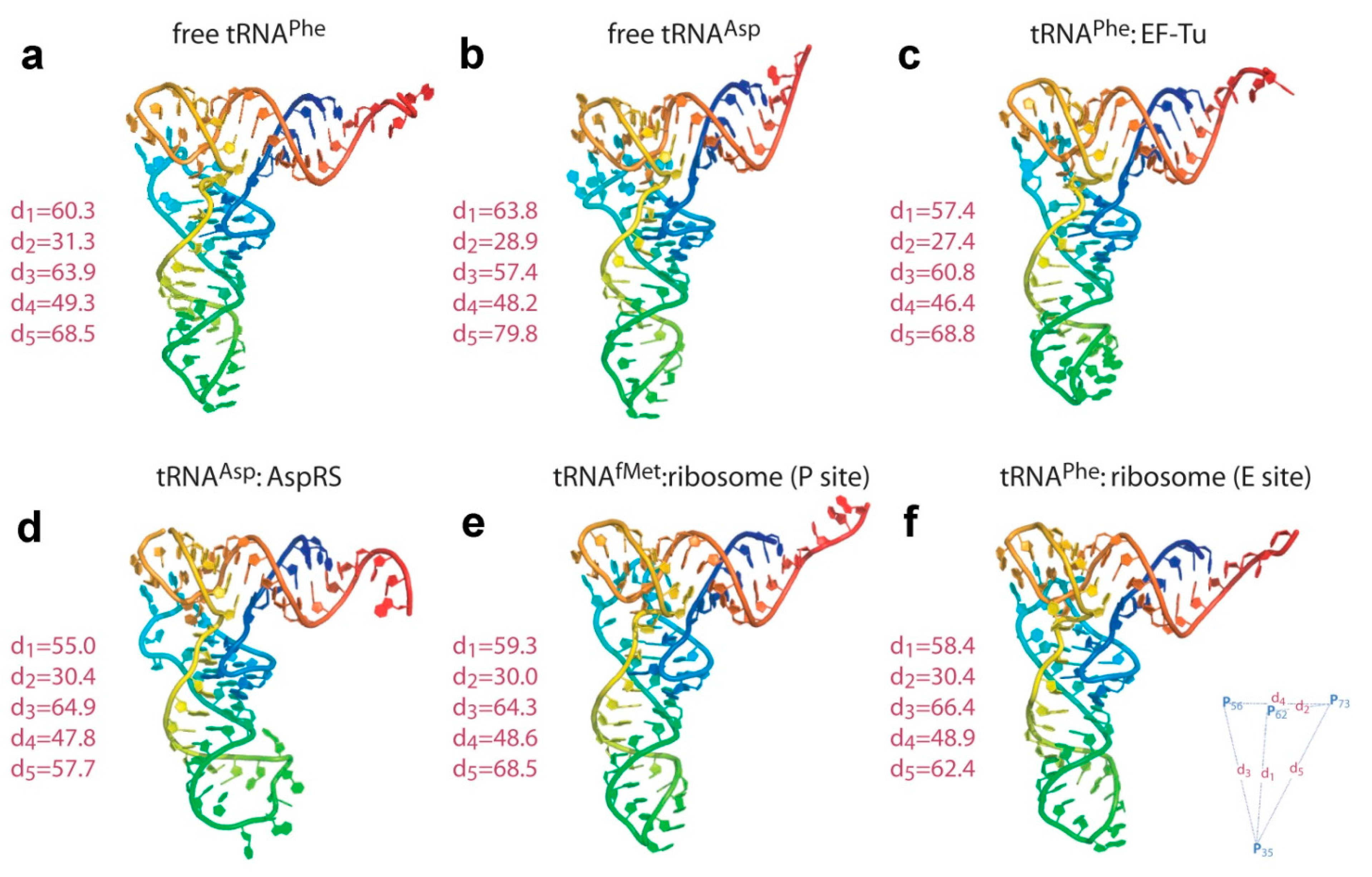
2. tRNA Dynamics an Overview
3. Structural Snapshots of tRNA Conformation Plasticity during Translation
3.1. Initiation

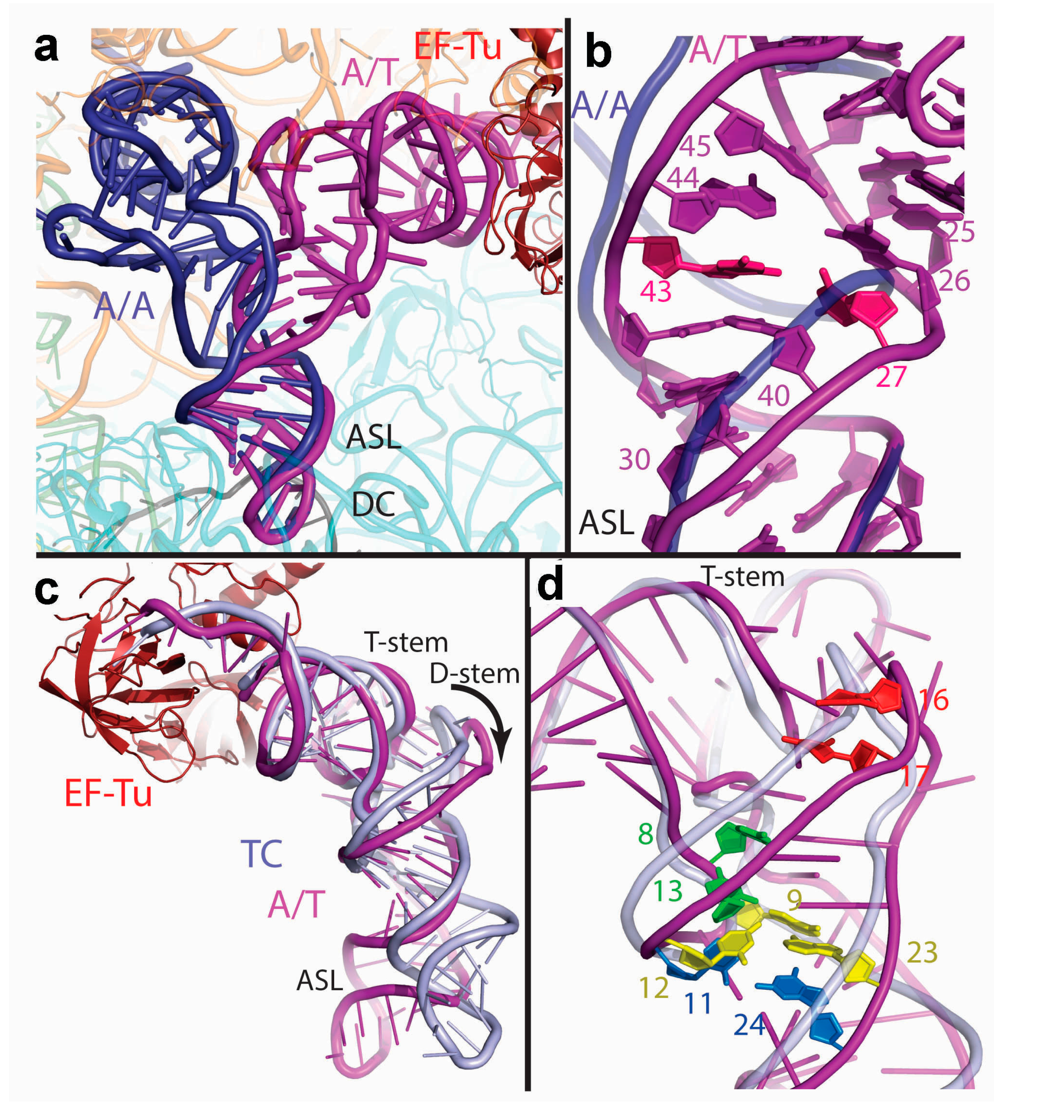
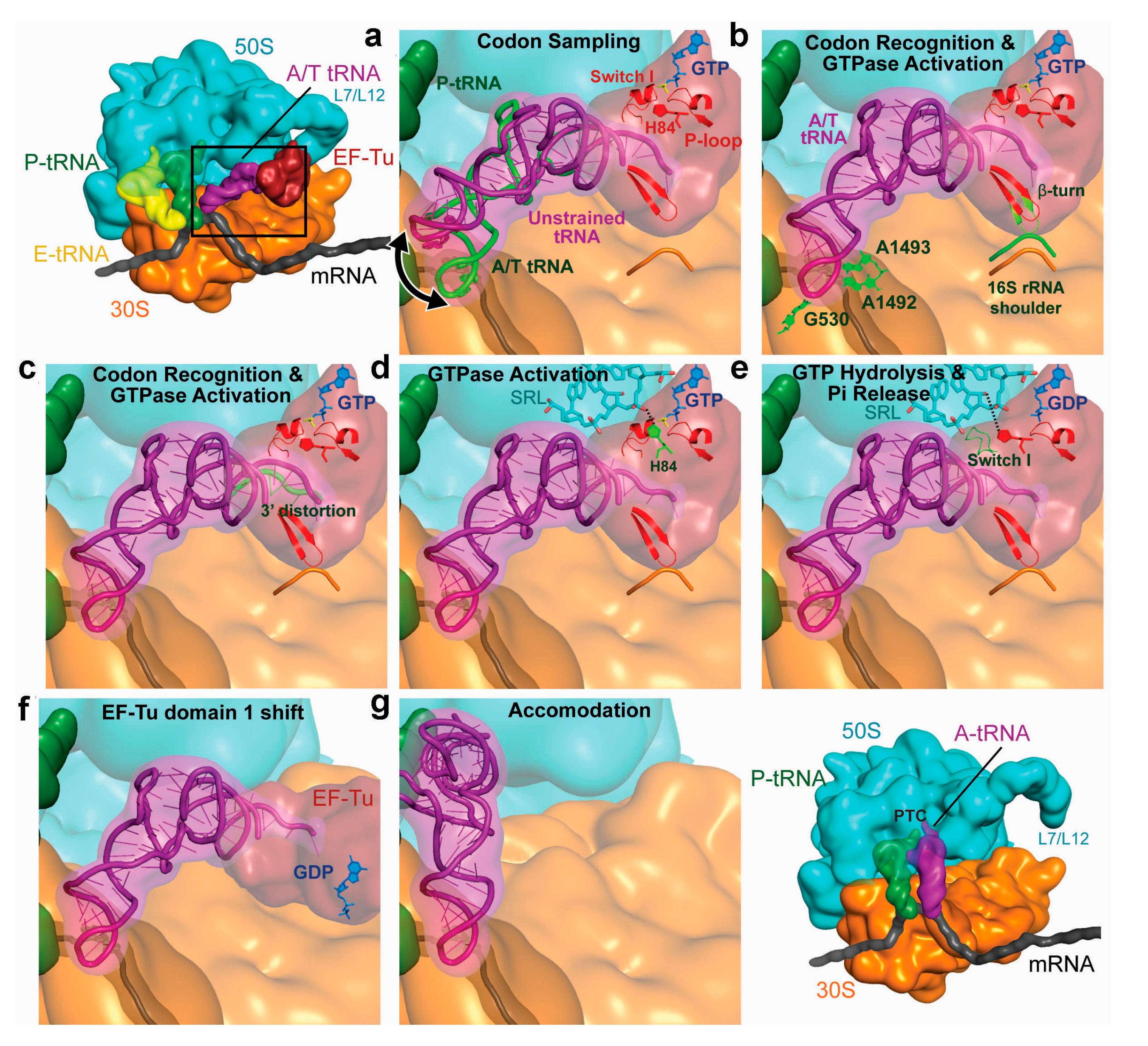
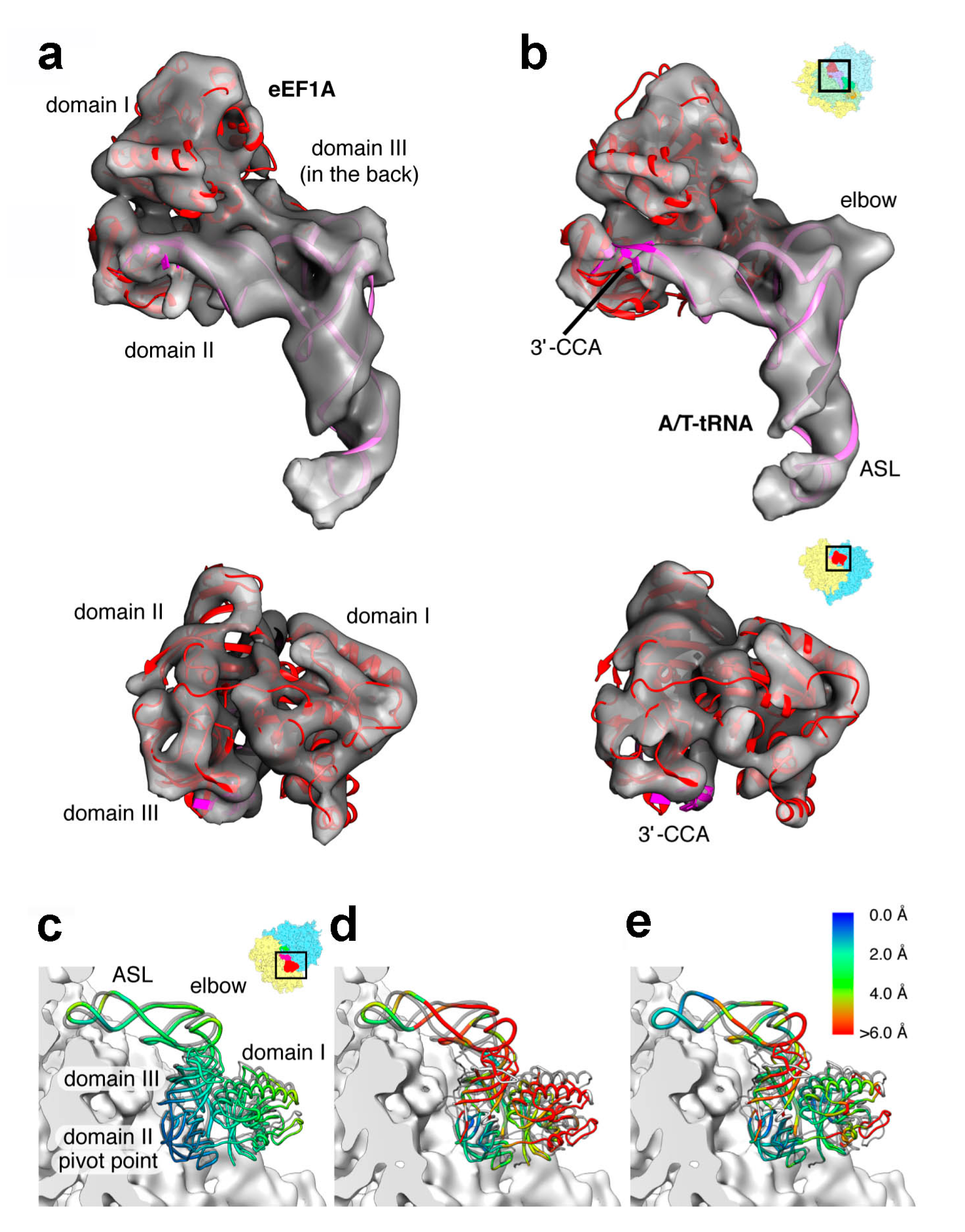
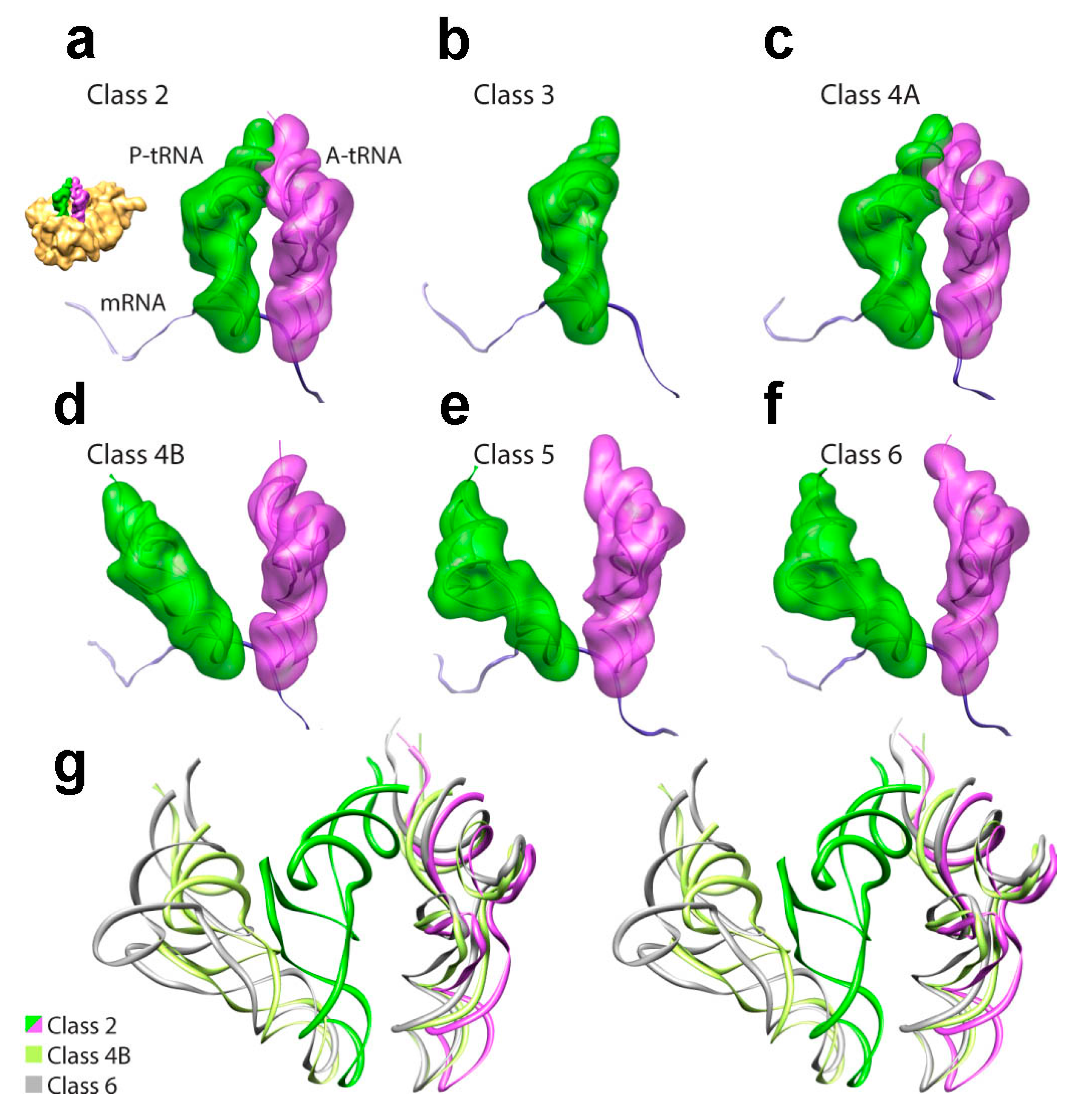
3.2. Decoding



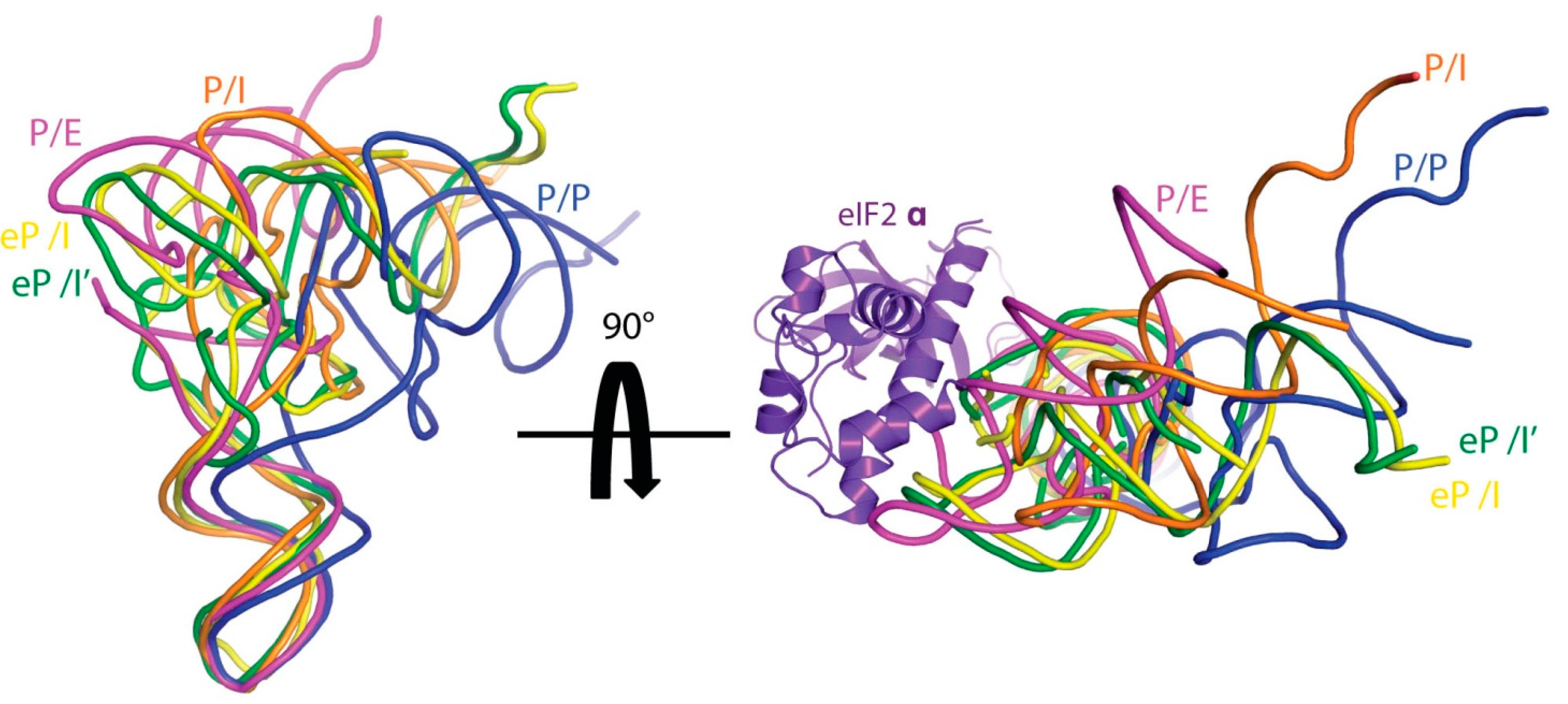
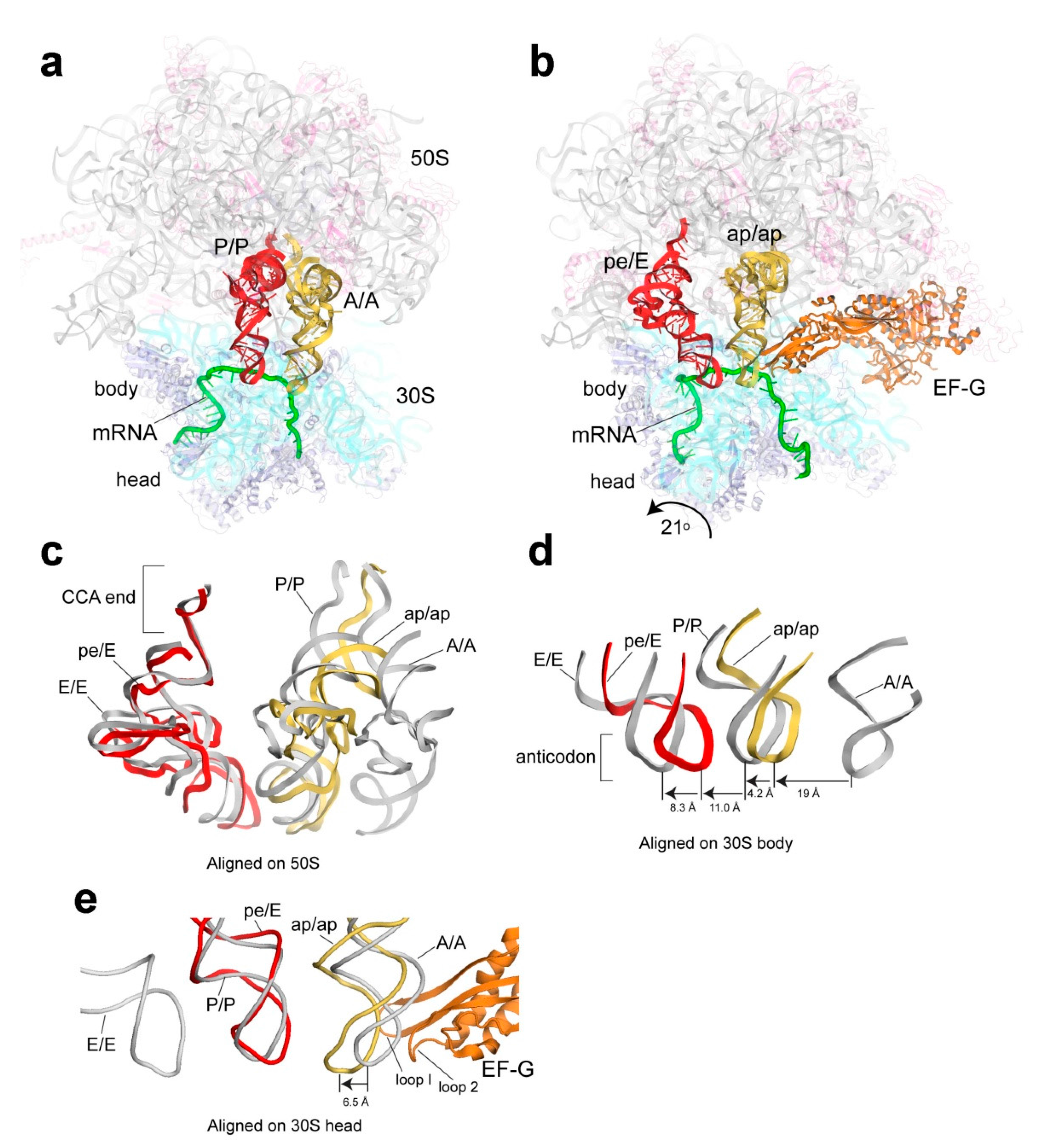
3.3. Translocation


4. The Stringent Response

5. Concluding Remarks
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Uemura, S.; Aitken, C.E.; Korlach, J.; Flusberg, B.A.; Turner, S.W.; Puglisi, J.D. Real-time tRNA transit on single translating ribosomes at codon resolution. Nature 2010, 464, 1012–1017. [Google Scholar] [CrossRef] [PubMed]
- Holley, R.W.; Apgar, J.; Everett, G.A.; Madison, J.T.; Marquisee, M.; Merrill, S.H.; Penswick, J.R.; Zamir, A. Structure of a ribonucleic acid. Science 1965, 147, 1462–1465. [Google Scholar] [CrossRef] [PubMed]
- Kim, S.H.; Quigley, G.J.; Suddath, F.L.; McPherson, A.; Sneden, D.; Kim, J.J.; Weinzierl, J.; Rich, A. Three-dimensional structure of yeast phenylalanine transfer RNA: Folding of the polynucleotide chain. Science 1973, 179, 285–288. [Google Scholar] [CrossRef] [PubMed]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.; Klug, A. Structure of yeast phenylalanine tRNA at 3 Å resolution. Nature 1974, 250, 546–551. [Google Scholar] [CrossRef] [PubMed]
- Robertus, J.D.; Ladner, J.E.; Finch, J.T.; Rhodes, D.; Brown, R.S.; Clark, B.F.; Klug, A. Correlation between three-dimensional structure and chemical reactivity of transfer RNA. Nucleic Acids Res. 1974, 1, 927–932. [Google Scholar] [CrossRef] [PubMed]
- Suddath, F.L.; Quigley, G.J.; McPherson, A.; Sneden, D.; Kim, J.J.; Kim, S.H.; Rich, A. Three-dimensional structure of yeast phenylalanine transfer RNA at 3.0 Å resolution. Nature 1974, 248, 20–24. [Google Scholar]
- Crick, F.H. On protein synthesis. Symp. Soc. Exp. Biol. 1958, 12, 138–163. [Google Scholar] [PubMed]
- Westhof, E.; Dumas, P.; Moras, D. Restrained refinement of two crystalline forms of yeast aspartic acid and phenylalanine transfer RNA crystals. Acta Crystallogr. A 1988, 44, 112–123. [Google Scholar] [CrossRef] [PubMed]
- Fulle, S.; Gohlke, H. Analyzing the flexibility of RNA structures by constraint counting. Biophys. J. 2008, 94, 4202–4219. [Google Scholar] [CrossRef] [PubMed]
- Giege, R.; Juhling, F.; Putz, J.; Stadler, P.; Sauter, C.; Florentz, C. Structure of transfer RNAs: Similarity and variability. Wiley Interdiscip. Rev. RNA 2012, 3, 37–61. [Google Scholar] [CrossRef] [PubMed]
- Nissen, P.; Kjeldgaard, M.; Thirup, S.; Polekhina, G.; Reshetnikova, L.; Clark, B.F.; Nyborg, J. Crystal structure of the ternary complex of Phe-tRNAPhe, Ef-Tu, and a GTP analog. Science 1995, 270, 1464–1472. [Google Scholar] [CrossRef] [PubMed]
- Ruff, M.; Krishnaswamy, S.; Boeglin, M.; Poterszman, A.; Mitschler, A.; Podjarny, A.; Rees, B.; Thierry, J.C.; Moras, D. Class II aminoacyl transfer RNA synthetases: Crystal structure of yeast aspartyl-tRNA synthetase complexed with tRNAAsp. Science 1991, 252, 1682–1689. [Google Scholar] [CrossRef] [PubMed]
- Selmer, M.; Dunham, C.M.; Murphy, F.V.T.; Weixlbaumer, A.; Petry, S.; Kelley, A.C.; Weir, J.R.; Ramakrishnan, V. Structure of the 70S ribosome complexed with mRNA and tRNA. Science 2006, 313, 1935–1942. [Google Scholar] [CrossRef] [PubMed]
- Giege, R. Toward a more complete view of tRNA biology. Nat. Struct. Mol. Biol. 2008, 15, 1007–1014. [Google Scholar] [CrossRef] [PubMed]
- Moazed, D.; Noller, H.F. Interaction of tRNA with 23S rRNA in the ribosomal A, P, and E sites. Cell 1989, 57, 585–597. [Google Scholar] [CrossRef] [PubMed]
- Bretscher, M.S. Translocation in protein synthesis: A hybrid structure model. Nature 1968, 218, 675–677. [Google Scholar] [CrossRef] [PubMed]
- Moazed, D.; Noller, H.F. Intermediate states in the movement of transfer RNA in the ribosome. Nature 1989, 342, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V. Quality control of mRNA decoding on the bacterial ribosome. Adv. Protein Chem. Struct. Biol. 2012, 86, 95–128. [Google Scholar] [PubMed]
- Aitken, C.E.; Petrov, A.; Puglisi, J.D. Single ribosome dynamics and the mechanism of translation. Annu. Rev. Biophys. 2010, 39, 491–513. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V.; Fricke, R.; Wintermeyer, W. Transient conformational states of aminoacyl-tRNA during ribosome binding catalyzed by elongation factor Tu. Biochemistry 1994, 33, 12267–12275. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V.; Fricke, R.; Kuhn, L.; Wintermeyer, W. Codon-dependent conformational change of elongation factor Tu preceding GTP hydrolysis on the ribosome. EMBO J. 1995, 14, 2613–2619. [Google Scholar] [PubMed]
- Rodnina, M.V.; Pape, T.; Fricke, R.; Wintermeyer, W. Elongation factor Tu, a GTPase triggered by codon recognition on the ribosome: Mechanism and GTP consumption. Biochem. Cell Biol. 1995, 73, 1221–1227. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V.; Pape, T.; Fricke, R.; Kuhn, L.; Wintermeyer, W. Initial binding of the elongation factor Tu·GTP·aminoacyl-tRNA complex preceding codon recognition on the ribosome. J. Biol. Chem. 1996, 271, 646–652. [Google Scholar] [CrossRef] [PubMed]
- Pape, T.; Wintermeyer, W.; Rodnina, M.V. Complete kinetic mechanism of elongation factor Tu-dependent binding of aminoacyl-tRNA to the A site of the E. coli ribosome. EMBO J. 1998, 17, 7490–7497. [Google Scholar] [CrossRef]
- Rodnina, M.V.; Daviter, T.; Gromadski, K.; Wintermeyer, W. Structural dynamics of ribosomal RNA during decoding on the ribosome. Biochimie 2002, 84, 745–754. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, S.C.; Gonzalez, R.L.; Kim, H.D.; Chu, S.; Puglisi, J.D. tRNA selection and kinetic proofreading in translation. Nat. Struct. Mol. Biol. 2004, 11, 1008–1014. [Google Scholar] [CrossRef] [PubMed]
- Geggier, P.; Dave, R.; Feldman, M.B.; Terry, D.S.; Altman, R.B.; Munro, J.B.; Blanchard, S.C. Conformational sampling of aminoacyl-tRNA during selection on the bacterial ribosome. J. Mol. Biol. 2010, 399, 576–595. [Google Scholar] [CrossRef] [PubMed]
- Semenkov, Y.P.; Rodnina, M.V.; Wintermeyer, W. Energetic contribution of tRNA hybrid state formation to translocation catalysis on the ribosome. Nat. Struct. Biol. 2000, 7, 1027–1031. [Google Scholar] [CrossRef] [PubMed]
- Blanchard, S.C.; Kim, H.D.; Gonzalez, R.L., Jr.; Puglisi, J.D.; Chu, S. tRNA dynamics on the ribosome during translation. Proc. Natl. Acad. Sci. USA 2004, 101, 12893–12898. [Google Scholar] [CrossRef] [PubMed]
- Dorner, S.; Brunelle, J.L.; Sharma, D.; Green, R. The hybrid state of tRNA binding is an authentic translation elongation intermediate. Nat. Struct. Mol. Biol. 2006, 13, 234–241. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Kirillov, S.V.; Cooperman, B.S. Kinetically competent intermediates in the translocation step of protein synthesis. Mol. Cell 2007, 25, 519–529. [Google Scholar] [CrossRef] [PubMed]
- Ermolenko, D.N.; Spiegel, P.C.; Majumdar, Z.K.; Hickerson, R.P.; Clegg, R.M.; Noller, H.F. The antibiotic viomycin traps the ribosome in an intermediate state of translocation. Nat. Struct. Mol. Biol. 2007, 14, 493–497. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.D.; Puglisi, J.D.; Chu, S. Fluctuations of transfer RNAs between classical and hybrid states. Biophys. J. 2007, 93, 3575–3582. [Google Scholar] [CrossRef] [PubMed]
- Fei, J.; Kosuri, P.; MacDougall, D.D.; Gonzalez, R.L., Jr. Coupling of ribosomal l1 stalk and tRNA dynamics during translation elongation. Mol. Cell 2008, 30, 348–359. [Google Scholar] [CrossRef] [PubMed]
- Fei, J.; Richard, A.C.; Bronson, J.E.; Gonzalez, R.L., Jr. Transfer RNA-mediated regulation of ribosome dynamics during protein synthesis. Nat. Struct. Mol. Biol. 2011, 18, 1043–1051. [Google Scholar] [CrossRef] [PubMed]
- Munro, J.B.; Altman, R.B.; O’Connor, N.; Blanchard, S.C. Identification of two distinct hybrid state intermediates on the ribosome. Mol. Cell 2007, 25, 505–517. [Google Scholar] [CrossRef] [PubMed]
- Munro, J.B.; Altman, R.B.; Tung, C.S.; Cate, J.H.; Sanbonmatsu, K.Y.; Blanchard, S.C. Spontaneous formation of the unlocked state of the ribosome is a multistep process. Proc. Natl. Acad. Sci. USA 2010, 107, 709–714. [Google Scholar] [CrossRef] [PubMed]
- Munro, J.B.; Altman, R.B.; Tung, C.S.; Sanbonmatsu, K.Y.; Blanchard, S.C. A fast dynamic mode of the EF-G-bound ribosome. EMBO J. 2010, 29, 770–781. [Google Scholar] [CrossRef] [PubMed]
- Cornish, P.V.; Ermolenko, D.N.; Noller, H.F.; Ha, T. Spontaneous intersubunit rotation in single ribosomes. Mol. Cell 2008, 30, 578–588. [Google Scholar] [CrossRef] [PubMed]
- Cornish, P.V.; Ermolenko, D.N.; Staple, D.W.; Hoang, L.; Hickerson, R.P.; Noller, H.F.; Ha, T. Following movement of the L1 stalk between three functional states in single ribosomes. Proc. Natl. Acad. Sci. USA 2009, 106, 2571–2576. [Google Scholar] [CrossRef] [PubMed]
- Walker, S.E.; Shoji, S.; Pan, D.; Cooperman, B.S.; Fredrick, K. Role of hybrid tRNA-binding states in ribosomal translocation. Proc. Natl. Acad. Sci. USA 2008, 105, 9192–9197. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Stevens, B.; Kaur, J.; Cabral, D.; Liu, H.; Wang, Y.; Zhang, H.; Rosenblum, G.; Smilansky, Z.; Goldman, Y.E.; et al. Single-molecule fluorescence measurements of ribosomal translocation dynamics. Mol. Cell 2011, 42, 367–377. [Google Scholar]
- Ning, W.; Fei, J.; Gonzalez, R.L., Jr. The ribosome uses cooperative conformational changes to maximize and regulate the efficiency of translation. Proc. Natl. Acad. Sci. USA 2014, 111, 12073–12078. [Google Scholar] [CrossRef] [PubMed]
- Holtkamp, W.; Cunha, C.E.; Peske, F.; Konevega, A.L.; Wintermeyer, W.; Rodnina, M.V. GTP hydrolysis by EF-G synchronizes tRNA movement on small and large ribosomal subunits. EMBO J. 2014, 33, 1073–1085. [Google Scholar] [CrossRef] [PubMed]
- Prabhakaran, M.; Harvey, S.C.; Mao, B.; McCammon, J.A. Molecular dynamics of phenylalanine transfer RNA. J. Biomol. Struct. Dyn. 1983, 1, 357–369. [Google Scholar] [CrossRef] [PubMed]
- Harvey, S.C.; Prabhakaran, M.; Mao, B.; McCammon, J.A. Phenylalanine transfer RNA: Molecular dynamics simulation. Science 1984, 223, 1189–1191. [Google Scholar] [CrossRef] [PubMed]
- Nakamura, S.; Doi, J. Dynamics of transfer RNAs analyzed by normal mode calculation. Nucleic Acids Res. 1994, 22, 514–521. [Google Scholar] [CrossRef] [PubMed]
- Friederich, M.W.; Vacano, E.; Hagerman, P.J. Global flexibility of tertiary structure in RNA: Yeast tRNAPhe as a model system. Proc. Natl. Acad. Sci. USA 1998, 95, 3572–3577. [Google Scholar] [CrossRef] [PubMed]
- Auffinger, P.; Louise-May, S.; Westhof, E. Molecular dynamics simulations of solvated yeast tRNAAsp. Biophys. J. 1999, 76, 50–64. [Google Scholar] [CrossRef] [PubMed]
- Lahiri, A.; Nilsson, L. Molecular dynamics of the anticodon domain of yeast tRNAPhe: Codon-anticodon interaction. Biophys. J. 2000, 79, 2276–2289. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Frank, J. Transfer RNA in the hybrid P/E state: Correlating molecular dynamics simulations with cryo-EM data. Proc. Natl. Acad. Sci. USA 2007, 104, 16540–16545. [Google Scholar] [CrossRef] [PubMed]
- Alexander, R.W.; Eargle, J.; Luthey-Schulten, Z. Experimental and computational determination of tRNA dynamics. FEBS Lett. 2010, 584, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Caulfield, T.R.; Devkota, B.; Rollins, G.C. Examinations of tRNA range of motion using simulations of cryo-EM microscopy and X-ray data. J. Biophys. 2011, 2011, 219515. [Google Scholar] [CrossRef] [PubMed]
- Sethi, A.; Eargle, J.; Black, A.A.; Luthey-Schulten, Z. Dynamical networks in tRNA:protein complexes. Proc. Natl. Acad. Sci. USA 2009, 106, 6620–6625. [Google Scholar] [CrossRef] [PubMed]
- Eargle, J.; Black, A.A.; Sethi, A.; Trabuco, L.G.; Luthey-Schulten, Z. Dynamics of recognition between tRNA and elongation factor Tu. J. Mol. Biol. 2008, 377, 1382–1405. [Google Scholar] [CrossRef] [PubMed]
- Sanbonmatsu, K.Y.; Joseph, S. Understanding discrimination by the ribosome: Stability testing and groove measurement of codon-anticodon pairs. J. Mol. Biol. 2003, 328, 33–47. [Google Scholar] [CrossRef] [PubMed]
- Sanbonmatsu, K.Y.; Joseph, S.; Tung, C.S. Simulating movement of tRNA into the ribosome during decoding. Proc. Natl. Acad. Sci. USA 2005, 102, 15854–15859. [Google Scholar] [CrossRef] [PubMed]
- Almlof, M.; Ander, M.; Aqvist, J. Energetics of codon-anticodon recognition on the small ribosomal subunit. Biochemistry 2007, 46, 200–209. [Google Scholar] [CrossRef] [PubMed]
- Sund, J.; Ander, M.; Aqvist, J. Principles of stop-codon reading on the ribosome. Nature 2010, 465, 947–950. [Google Scholar] [CrossRef] [PubMed]
- Whitford, P.C.; Geggier, P.; Altman, R.B.; Blanchard, S.C.; Onuchic, J.N.; Sanbonmatsu, K.Y. Accommodation of aminoacyl-tRNA into the ribosome involves reversible excursions along multiple pathways. RNA 2010, 16, 1196–1204. [Google Scholar] [CrossRef] [PubMed]
- Whitford, P.C.; Ahmed, A.; Yu, Y.; Hennelly, S.P.; Tama, F.; Spahn, C.M.; Onuchic, J.N.; Sanbonmatsu, K.Y. Excited states of ribosome translocation revealed through integrative molecular modeling. Proc. Natl Acad. Sci. USA 2011, 108, 18943–18948. [Google Scholar] [CrossRef] [PubMed]
- Whitford, P.C.; Blanchard, S.C.; Cate, J.H.; Sanbonmatsu, K.Y. Connecting the kinetics and energy landscape of tRNA translocation on the ribosome. PLoS Comput. Biol. 2013, 9, e1003003. [Google Scholar] [CrossRef] [PubMed]
- Bock, L.V.; Blau, C.; Schroder, G.F.; Davydov, I.I.; Fischer, N.; Stark, H.; Rodnina, M.V.; Vaiana, A.C.; Grubmuller, H. Energy barriers and driving forces in tRNA translocation through the ribosome. Nat. Struct. Mol. Biol. 2013, 20, 1390–1396. [Google Scholar] [CrossRef] [PubMed]
- Satpati, P.; Sund, J.; Aqvist, J. Structure-based energetics of mRNA decoding on the ribosome. Biochemistry 2014, 53, 1714–1722. [Google Scholar] [CrossRef] [PubMed]
- Aqvist, J.; Lind, C.; Sund, J.; Wallin, G. Bridging the gap between ribosome structure and biochemistry by mechanistic computations. Curr. Opin. Struct. Biol. 2012, 22, 815–823. [Google Scholar] [CrossRef] [PubMed]
- Sanbonmatsu, K.Y. Computational studies of molecular machines: The ribosome. Curr. Opin. Struct. Biol. 2012, 22, 168–174. [Google Scholar] [CrossRef] [PubMed]
- Yarus, M.; Valle, M.; Frank, J. A twisted tRNA intermediate sets the threshold for decoding. RNA 2003, 9, 384–385. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.; Sengupta, J.; Gao, H.; Li, W.; Valle, M.; Zavialov, A.; Ehrenberg, M. The role of tRNA as a molecular spring in decoding, accommodation, and peptidyl transfer. FEBS Lett. 2005, 579, 959–962. [Google Scholar] [CrossRef] [PubMed]
- Laursen, B.S.; Sorensen, H.P.; Mortensen, K.K.; Sperling-Petersen, H.U. Initiation of protein synthesis in bacteria. Microbiol. Mol. Biol. Rev. 2005, 69, 101–123. [Google Scholar] [CrossRef] [PubMed]
- Antoun, A.; Pavlov, M.Y.; Lovmar, M.; Ehrenberg, M. How initiation factors tune the rate of initiation of protein synthesis in bacteria. EMBO J. 2006, 25, 2539–2550. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, A.; Marzi, S.; Jenner, L.; Myasnikov, A.; Romby, P.; Yusupova, G.; Klaholz, B.P.; Yusupov, M. A structural view of translation initiation in bacteria. Cell Mol. Life Sci. 2009, 66, 423–436. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.A.; Aitken, C.E.; Puglisi, J.D. GTP hydrolysis by IF2 guides progression of the ribosome into elongation. Mol. Cell 2009, 35, 37–47. [Google Scholar] [CrossRef] [PubMed]
- Milon, P.; Rodnina, M.V. Kinetic control of translation initiation in bacteria. Crit. Rev. Biochem. Mol. Biol. 2012, 47, 334–348. [Google Scholar] [CrossRef] [PubMed]
- Tsai, A.; Petrov, A.; Marshall, R.A.; Korlach, J.; Uemura, S.; Puglisi, J.D. Heterogeneous pathways and timing of factor departure during translation initiation. Nature 2012, 487, 390–393. [Google Scholar] [CrossRef] [PubMed]
- Elvekrog, M.M.; Gonzalez, R.L., Jr. Conformational selection of translation initiation factor 3 signals proper substrate selection. Nat. Struct. Mol. Biol. 2013, 20, 628–633. [Google Scholar] [CrossRef] [PubMed]
- Allen, G.S.; Zavialov, A.; Gursky, R.; Ehrenberg, M.; Frank, J. The cryo-EM structure of a translation initiation complex from Escherichia coli. Cell 2005, 121, 703–712. [Google Scholar] [CrossRef] [PubMed]
- Myasnikov, A.G.; Marzi, S.; Simonetti, A.; Giuliodori, A.M.; Gualerzi, C.O.; Yusupova, G.; Yusupov, M.; Klaholz, B.P. Conformational transition of initiation factor 2 from the GTP- to GDP-bound state visualized on the ribosome. Nat. Struct. Mol. Biol. 2005, 12, 1145–1149. [Google Scholar] [CrossRef] [PubMed]
- Julian, P.; Milon, P.; Agirrezabala, X.; Lasso, G.; Gil, D.; Rodnina, M.V.; Valle, M. The cryo-EM structure of a complete 30S translation initiation complex from Escherichia coli. PLoS Biol. 2011, 9, e1001095. [Google Scholar] [CrossRef] [PubMed]
- Simonetti, A.; Marzi, S.; Myasnikov, A.G.; Fabbretti, A.; Yusupov, M.; Gualerzi, C.O.; Klaholz, B.P. Structure of the 30S translation initiation complex. Nature 2008, 455, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Korostelev, A.; Trakhanov, S.; Asahara, H.; Laurberg, M.; Lancaster, L.; Noller, H.F. Interactions and dynamics of the Shine-Dalgarno helix in the 70S ribosome. Proc. Natl. Acad. Sci. USA 2007, 104, 16840–16843. [Google Scholar] [CrossRef] [PubMed]
- Shine, J.; Dalgarno, L. The 3'-terminal sequence of Escherichia coli 16S ribosomal RNA: Complementarity to nonsense triplets and ribosome binding sites. Proc. Natl. Acad. Sci. USA 1974, 71, 1342–1346. [Google Scholar] [CrossRef] [PubMed]
- Yusupova, G.; Jenner, L.; Rees, B.; Moras, D.; Yusupov, M. Structural basis for messenger RNA movement on the ribosome. Nature 2006, 444, 391–394. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, A.G. The scanning mechanism of eukaryotic translation initiation. Annu. Rev. Biochem. 2014, 83, 779–812. [Google Scholar] [CrossRef] [PubMed]
- Acker, M.G.; Lorsch, J.R. Mechanism of ribosomal subunit joining during eukaryotic translation initiation. Biochem. Soc. Trans. 2008, 36, 653–657. [Google Scholar] [CrossRef] [PubMed]
- Sonenberg, N.; Hinnebusch, A.G. Regulation of translation initiation in eukaryotes: Mechanisms and biological targets. Cell 2009, 136, 731–745. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.J.; Hellen, C.U.; Pestova, T.V. The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 2010, 11, 113–127. [Google Scholar] [CrossRef] [PubMed]
- Aitken, C.E.; Lorsch, J.R. A mechanistic overview of translation initiation in eukaryotes. Nat. Struct. Mol. Biol. 2012, 19, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Voigts-Hoffmann, F.; Klinge, S.; Ban, N. Structural insights into eukaryotic ribosomes and the initiation of translation. Curr. Opin. Struct. Biol. 2012, 22, 768–777. [Google Scholar] [CrossRef] [PubMed]
- Siridechadilok, B.; Fraser, C.S.; Hall, R.J.; Doudna, J.A.; Nogales, E. Structural roles for human translation factor eIF3 in initiation of protein synthesis. Science 2005, 310, 1513–1515. [Google Scholar] [CrossRef] [PubMed]
- Passmore, L.A.; Schmeing, T.M.; Maag, D.; Applefield, D.J.; Acker, M.G.; Algire, M.A.; Lorsch, J.R.; Ramakrishnan, V. The eukaryotic translation initiation factors eIF1 and eIF1A induce an open conformation of the 40S ribosome. Mol. Cell 2007, 26, 41–50. [Google Scholar] [CrossRef] [PubMed]
- Rabl, J.; Leibundgut, M.; Ataide, S.F.; Haag, A.; Ban, N. Crystal structure of the eukaryotic 40S ribosomal subunit in complex with initiation factor 1. Science 2011, 331, 730–736. [Google Scholar] [CrossRef] [PubMed]
- Weisser, M.; Voigts-Hoffmann, F.; Rabl, J.; Leibundgut, M.; Ban, N. The crystal structure of the eukaryotic 40S ribosomal subunit in complex with eIF1 and eIF1A. Nat. Struct. Mol. Biol. 2013, 20, 1015–1017. [Google Scholar] [CrossRef] [PubMed]
- Klinge, S.; Voigts-Hoffmann, F.; Leibundgut, M.; Arpagaus, S.; Ban, N. Crystal structure of the eukaryotic 60S ribosomal subunit in complex with initiation factor 6. Science 2011, 334, 941–948. [Google Scholar] [CrossRef] [PubMed]
- Lomakin, I.B.; Steitz, T.A. The initiation of mammalian protein synthesis and mRNA scanning mechanism. Nature 2013, 500, 307–311. [Google Scholar] [CrossRef] [PubMed]
- Hashem, Y.; des Georges, A.; Dhote, V.; Langlois, R.; Liao, H.Y.; Grassucci, R.A.; Hellen, C.U.; Pestova, T.V.; Frank, J. Structure of the mammalian ribosomal 43S preinitiation complex bound to the scanning factor DHX29. Cell 2013, 153, 1108–1119. [Google Scholar] [CrossRef] [PubMed]
- Erzberger, J.P.; Stengel, F.; Pellarin, R.; Zhang, S.; Schaefer, T.; Aylett, C.H.; Cimermancic, P.; Boehringer, D.; Sali, A.; Aebersold, R.; et al. Molecular architecture of the 40S·eIF1·eIF3 translation initiation complex. Cell 2014, 158, 1123–1135. [Google Scholar]
- Hussain, T.; Llacer, J.L.; Fernandez, I.S.; Munoz, A.; Martin-Marcos, P.; Savva, C.G.; Lorsch, J.R.; Hinnebusch, A.G.; Ramakrishnan, V. Structural changes enable start codon recognition by the eukaryotic translation initiation complex. Cell 2014, 159, 597–607. [Google Scholar] [CrossRef] [PubMed]
- Saini, A.K.; Nanda, J.S.; Lorsch, J.R.; Hinnebusch, A.G. Regulatory elements in eIF1A control the fidelity of start codon selection by modulating tRNAiMet binding to the ribosome. Genes Dev. 2010, 24, 97–110. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, I.S.; Bai, X.C.; Hussain, T.; Kelley, A.C.; Lorsch, J.R.; Ramakrishnan, V.; Scheres, S.H. Molecular architecture of a eukaryotic translational initiation complex. Science 2013, 342, 1240585. [Google Scholar] [CrossRef] [PubMed]
- Kuhle, B.; Ficner, R. Structural insight into the recognition of amino-acylated initiator tRNA by eIF5B in the 80S initiation complex. BMC Struct. Biol. 2014, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Kuhle, B.; Ficner, R. eIF5B employs a novel domain release mechanism to catalyze ribosomal subunit joining. EMBO J. 2014, 33, 1177–1191. [Google Scholar] [CrossRef] [PubMed]
- Schmitt, E.; Panvert, M.; Lazennec-Schurdevin, C.; Coureux, P.D.; Perez, J.; Thompson, A.; Mechulam, Y. Structure of the ternary initiation complex aIF2-GDPNP-methionylated initiator tRNA. Nat. Struct. Mol. Biol. 2012, 19, 450–454. [Google Scholar] [CrossRef] [PubMed]
- Tourigny, D.S.; Fernandez, I.S.; Kelley, A.C.; Ramakrishnan, V. Elongation factor g bound to the ribosome in an intermediate state of translocation. Science 2013, 340, 1235490. [Google Scholar] [CrossRef] [PubMed]
- Pape, T.; Wintermeyer, W.; Rodnina, M. Induced fit in initial selection and proofreading of aminoacyl-tRNA on the ribosome. EMBO J. 1999, 18, 3800–3807. [Google Scholar] [CrossRef] [PubMed]
- Rodnina, M.V.; Wintermeyer, W. Fidelity of aminoacyl-tRNA selection on the ribosome: Kinetic and structural mechanisms. Annu. Rev. Biochem. 2001, 70, 415–435. [Google Scholar] [CrossRef] [PubMed]
- Gromadski, K.B.; Rodnina, M.V. Kinetic determinants of high-fidelity tRNA discrimination on the ribosome. Mol. Cell 2004, 13, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Gromadski, K.B.; Daviter, T.; Rodnina, M.V. A uniform response to mismatches in codon-anticodon complexes ensures ribosomal fidelity. Mol. Cell 2006, 21, 369–377. [Google Scholar] [CrossRef] [PubMed]
- Wohlgemuth, I.; Pohl, C.; Rodnina, M.V. Optimization of speed and accuracy of decoding in translation. EMBO J. 2010, 29, 3701–3709. [Google Scholar] [CrossRef] [PubMed]
- Johansson, M.; Zhang, J.; Ehrenberg, M. Genetic code translation displays a linear trade-off between efficiency and accuracy of tRNA selection. Proc. Natl. Acad. Sci. USA 2012, 109, 131–136. [Google Scholar] [CrossRef] [PubMed]
- Hopfield, J.J. Kinetic proofreading: A new mechanism for reducing errors in biosynthetic processes requiring high specificity. Proc. Natl. Acad. Sci. USA 1974, 71, 4135–4139. [Google Scholar] [CrossRef] [PubMed]
- Fourmy, D.; Recht, M.I.; Blanchard, S.C.; Puglisi, J.D. Structure of the A site of Escherichia coli 16S ribosomal RNA complexed with an aminoglycoside antibiotic. Science 1996, 274, 1367–1371. [Google Scholar] [CrossRef] [PubMed]
- Ogle, J.M.; Brodersen, D.E.; Clemons, W.M., Jr.; Tarry, M.J.; Carter, A.P.; Ramakrishnan, V. Recognition of cognate transfer RNA by the 30S ribosomal subunit. Science 2001, 292, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Ogle, J.M.; Murphy, F.V.; Tarry, M.J.; Ramakrishnan, V. Selection of tRNA by the ribosome requires a transition from an open to a closed form. Cell 2002, 111, 721–732. [Google Scholar] [CrossRef] [PubMed]
- Demeshkina, N.; Jenner, L.; Westhof, E.; Yusupov, M.; Yusupova, G. A new understanding of the decoding principle on the ribosome. Nature 2012, 484, 256–259. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, R.M.; Ramakrishnan, V. Structural basis of the translational elongation cycle. Annu. Rev. Biochem. 2013, 82, 203–236. [Google Scholar] [CrossRef] [PubMed]
- Khade, P.K.; Shi, X.; Joseph, S. Steric complementarity in the decoding center is important for tRNA selection by the ribosome. J. Mol. Biol. 2013, 425, 3778–3789. [Google Scholar] [CrossRef] [PubMed]
- Ogle, J.M.; Ramakrishnan, V. Structural insights into translational fidelity. Annu. Rev. Biochem. 2005, 74, 129–177. [Google Scholar] [CrossRef] [PubMed]
- Schmeing, T.M.; Ramakrishnan, V. What recent ribosome structures have revealed about the mechanism of translation. Nature 2009, 461, 1234–1242. [Google Scholar] [CrossRef] [PubMed]
- Agirrezabala, X.; Frank, J. Elongation in translation as a dynamic interaction among the ribosome, tRNA, and elongation factors EF-G and EF-Tu. Q. Rev. Biophys. 2009, 42, 159–200. [Google Scholar] [CrossRef] [PubMed]
- Zaher, H.S.; Green, R. Fidelity at the molecular level: Lessons from protein synthesis. Cell 2009, 136, 746–762. [Google Scholar] [CrossRef] [PubMed]
- Stark, H.; Rodnina, M.V.; Wieden, H.J.; Zemlin, F.; Wintermeyer, W.; van Heel, M. Ribosome interactions of aminoacyl-tRNA and elongation factor Tu in the codon-recognition complex. Nat. Struct. Biol. 2002, 9, 849–854. [Google Scholar] [PubMed]
- Valle, M.; Sengupta, J.; Swami, N.K.; Grassucci, R.A.; Burkhardt, N.; Nierhaus, K.H.; Agrawal, R.K.; Frank, J. Cryo-EM reveals an active role for aminoacyl-tRNA in the accommodation process. EMBO J. 2002, 21, 3557–3567. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.; Zavialov, A.; Li, W.; Stagg, S.M.; Sengupta, J.; Nielsen, R.C.; Nissen, P.; Harvey, S.C.; Ehrenberg, M.; Frank, J. Incorporation of aminoacyl-tRNA into the ribosome as seen by cryo-electron microscopy. Nat. Struct. Biol. 2003, 10, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Agirrezabala, X.; Lei, J.; Bouakaz, L.; Brunelle, J.L.; Ortiz-Meoz, R.F.; Green, R.; Sanyal, S.; Ehrenberg, M.; Frank, J. Recognition of aminoacyl-tRNA: A common molecular mechanism revealed by cryo-EM. EMBO J. 2008, 27, 3322–3331. [Google Scholar] [CrossRef] [PubMed]
- Villa, E.; Sengupta, J.; Trabuco, L.G.; LeBarron, J.; Baxter, W.T.; Shaikh, T.R.; Grassucci, R.A.; Nissen, P.; Ehrenberg, M.; Schulten, K.; et al. Ribosome-induced changes in elongation factor Tu conformation control GTP hydrolysis. Proc. Natl. Acad. Sci. USA 2009, 106, 1063–1068. [Google Scholar]
- Schuette, J.C.; Murphy, F.V.T.; Kelley, A.C.; Weir, J.R.; Giesebrecht, J.; Connell, S.R.; Loerke, J.; Mielke, T.; Zhang, W.; Penczek, P.A.; et al. GTPase activation of elongation factor EF-Tu by the ribosome during decoding. EMBO J. 2009, 28, 755–765. [Google Scholar]
- Agirrezabala, X.; Schreiner, E.; Trabuco, L.G.; Lei, J.; Ortiz-Meoz, R.F.; Schulten, K.; Green, R.; Frank, J. Structural insights into cognate versus near-cognate discrimination during decoding. EMBO J. 2011, 30, 1497–1507. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Neumann, P.; Konevega, A.L.; Bock, L.V.; Ficner, R.; Rodnina, M.V.; Stark, H. Structure of the E. coli ribosome–EF-Tu complex at <3 Å resolution by Cs-corrected cryo-EM. Nature 2015, 520, 567–570. [Google Scholar]
- Schmeing, T.M.; Voorhees, R.M.; Kelley, A.C.; Gao, Y.G.; Murphy, F.V.T.; Weir, J.R.; Ramakrishnan, V. The crystal structure of the ribosome bound to EF-Tu and aminoacyl-tRNA. Science 2009, 326, 688–694. [Google Scholar] [CrossRef] [PubMed]
- Schmeing, T.M.; Voorhees, R.M.; Kelley, A.C.; Ramakrishnan, V. How mutations in tRNA distant from the anticodon affect the fidelity of decoding. Nat. Struct. Mol. Biol. 2011, 18, 432–436. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, R.M.; Schmeing, T.M.; Kelley, A.C.; Ramakrishnan, V. The mechanism for activation of GTP hydrolysis on the ribosome. Science 2010, 330, 835–838. [Google Scholar] [CrossRef] [PubMed]
- Lee, T.H.; Blanchard, S.C.; Kim, H.D.; Puglisi, J.D.; Chu, S. The role of fluctuations in tRNA selection by the ribosome. Proc. Natl. Acad. Sci. USA 2007, 104, 13661–13665. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, R.M.; Weixlbaumer, A.; Loakes, D.; Kelley, A.C.; Ramakrishnan, V. Insights into substrate stabilization from snapshots of the peptidyl transferase center of the intact 70S ribosome. Nat. Struct. Mol. Biol. 2009, 16, 528–533. [Google Scholar] [CrossRef] [PubMed]
- Schultz, D.W.; Yarus, M. tRNA structure and ribosomal function. I. tRNA nucleotide 27–43 mutations enhance first position wobble. J. Mol. Biol. 1994, 235, 1381–1394. [Google Scholar]
- Favre, A.; Buchingham, R.; Thomas, G. tRNA tertiary structure in solution as probed by the photochemically induced 8–13 cross-link. Nucleic Acids Res. 1975, 2, 1421–1431. [Google Scholar] [CrossRef] [PubMed]
- Smith, D.; Yarus, M. Transfer RNA structure and coding specificity. II. A D-arm tertiary interaction that restricts coding range. J. Mol. Biol. 1989, 206, 503–511. [Google Scholar]
- Hirsh, D. Tryptophan tRNA of Escherichia coli. Nature 1970, 228, 57. [Google Scholar] [CrossRef] [PubMed]
- Cochella, L.; Green, R. An active role for tRNA in decoding beyond codon:anticodon pairing. Science 2005, 308, 1178–1180. [Google Scholar] [CrossRef] [PubMed]
- Piepenburg, O.; Pape, T.; Pleiss, J.A.; Wintermeyer, W.; Uhlenbeck, O.C.; Rodnina, M.V. Intact aminoacyl-tRNA is required to trigger GTP hydrolysis by elongation factor Tu on the ribosome. Biochemistry 2000, 39, 1734–1738. [Google Scholar] [CrossRef] [PubMed]
- Daviter, T.; Wieden, H.J.; Rodnina, M.V. Essential role of histidine 84 in elongation factor Tu for the chemical step of GTP hydrolysis on the ribosome. J. Mol. Biol. 2003, 332, 689–699. [Google Scholar] [CrossRef] [PubMed]
- Adamczyk, A.J.; Warshel, A. Converting structural information into an allosteric-energy-based picture for elongation factor Tu activation by the ribosome. Proc. Natl. Acad. Sci. USA 2011, 108, 9827–9832. [Google Scholar] [CrossRef] [PubMed]
- Liljas, A.; Ehrenberg, M.; Aqvist, J. Comment on “The mechanism for activation of GTP hydrolysis on the ribosome”. Science 2011, 333, 37, author reply 37. [Google Scholar] [PubMed]
- Kothe, U.; Rodnina, M.V. Delayed release of inorganic phosphate from elongation factor Tu following GTP hydrolysis on the ribosome. Biochemistry 2006, 45, 12767–12774. [Google Scholar] [CrossRef] [PubMed]
- Fahlman, R.P.; Dale, T.; Uhlenbeck, O.C. Uniform binding of aminoacylated transfer RNAs to the ribosomal A and P sites. Mol. Cell 2004, 16, 799–805. [Google Scholar] [CrossRef] [PubMed]
- Ledoux, S.; Uhlenbeck, O.C. Different aa-tRNAs are selected uniformly on the ribosome. Mol. Cell 2008, 31, 114–123. [Google Scholar] [CrossRef] [PubMed]
- Budkevich, T.V.; Giesebrecht, J.; Behrmann, E.; Loerke, J.; Ramrath, D.J.; Mielke, T.; Ismer, J.; Hildebrand, P.W.; Tung, C.S.; Nierhaus, K.H.; et al. Regulation of the mammalian elongation cycle by subunit rolling: A eukaryotic-specific ribosome rearrangement. Cell 2014, 158, 121–131. [Google Scholar]
- Moazed, D.; Robertson, J.M.; Noller, H.F. Interaction of elongation factors EF-G and EF-Tu with a conserved loop in 23S RNA. Nature 1988, 334, 362–364. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Saito, K.; Ishitani, R.; Ito, K.; Nureki, O. Structural basis for translation termination by archaeal RF1 and GTP-bound EF1α complex. Nucleic Acids Res. 2012, 40, 9319–9328. [Google Scholar] [CrossRef] [PubMed]
- Beringer, M.; Rodnina, M.V. The ribosomal peptidyl transferase. Mol. Cell 2007, 26, 311–321. [Google Scholar] [CrossRef] [PubMed]
- Simonovic, M.; Steitz, T.A. A structural view on the mechanism of the ribosome-catalyzed peptide bond formation. Biochim. Biophys. Acta 2009, 1789, 612–623. [Google Scholar] [CrossRef] [PubMed]
- Kuhlenkoetter, S.; Wintermeyer, W.; Rodnina, M.V. Different substrate-dependent transition states in the active site of the ribosome. Nature 2011, 476, 351–354. [Google Scholar] [CrossRef] [PubMed]
- Polikanov, Y.S.; Steitz, T.A.; Innis, C.A. A proton wire to couple aminoacyl-tRNA accommodation and peptide-bond formation on the ribosome. Nat. Struct. Mol. Biol. 2014, 21, 787–793. [Google Scholar] [CrossRef] [PubMed]
- Zavialov, A.V.; Ehrenberg, M. Peptidyl-tRNA regulates the GTPase activity of translation factors. Cell 2003, 114, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Aitken, C.E.; Puglisi, J.D. Following the intersubunit conformation of the ribosome during translation in real time. Nat. Struct. Mol. Biol. 2010, 17, 793–800. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.; Agrawal, R.K. A ratchet-like inter-subunit reorganization of the ribosome during translocation. Nature 2000, 406, 318–322. [Google Scholar] [CrossRef] [PubMed]
- Valle, M.; Zavialov, A.; Sengupta, J.; Rawat, U.; Ehrenberg, M.; Frank, J. Locking and unlocking of ribosomal motions. Cell 2003, 114, 123–134. [Google Scholar] [CrossRef] [PubMed]
- Sharma, D.; Southworth, D.R.; Green, R. EF-G-independent reactivity of a pre-translocation-state ribosome complex with the aminoacyl trna substrate puromycin supports an intermediate (hybrid) state of tRNA binding. RNA 2004, 10, 102–113. [Google Scholar] [CrossRef] [PubMed]
- Ermolenko, D.N.; Majumdar, Z.K.; Hickerson, R.P.; Spiegel, P.C.; Clegg, R.M.; Noller, H.F. Observation of intersubunit movement of the ribosome in solution using fret. J. Mol. Biol. 2007, 370, 530–540. [Google Scholar] [CrossRef] [PubMed]
- Marshall, R.A.; Dorywalska, M.; Puglisi, J.D. Irreversible chemical steps control intersubunit dynamics during translation. Proc. Natl. Acad. Sci. USA 2008, 105, 15364–15369. [Google Scholar] [CrossRef] [PubMed]
- Agirrezabala, X.; Lei, J.; Brunelle, J.L.; Ortiz-Meoz, R.F.; Green, R.; Frank, J. Visualization of the hybrid state of tRNA binding promoted by spontaneous ratcheting of the ribosome. Mol. Cell 2008, 32, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Julian, P.; Konevega, A.L.; Scheres, S.H.; Lazaro, M.; Gil, D.; Wintermeyer, W.; Rodnina, M.V.; Valle, M. Structure of ratcheted ribosomes with tRNAs in hybrid states. Proc. Natl. Acad. Sci. USA 2008, 105, 16924–16927. [Google Scholar] [CrossRef] [PubMed]
- Voorhees, R.M.; Fernandez, I.S.; Scheres, S.H.; Hegde, R.S. Structure of the mammalian ribosome-Sec61 complex to 3.4 Å resolution. Cell 2014, 157, 1632–1643. [Google Scholar]
- Svidritskiy, E.; Brilot, A.F.; Koh, C.S.; Grigorieff, N.; Korostelev, A.A. Structures of yeast 80S ribosome-tRNA complexes in the rotated and nonrotated conformations. Structure 2014, 22, 1210–1218. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Dunkle, J.A.; Cate, J.H. Structures of the ribosome in intermediate states of ratcheting. Science 2009, 325, 1014–1017. [Google Scholar] [CrossRef] [PubMed]
- Fischer, N.; Konevega, A.L.; Wintermeyer, W.; Rodnina, M.V.; Stark, H. Ribosome dynamics and trna movement by time-resolved electron cryomicroscopy. Nature 2010, 466, 329–333. [Google Scholar] [CrossRef] [PubMed]
- Budkevich, T.; Giesebrecht, J.; Altman, R.B.; Munro, J.B.; Mielke, T.; Nierhaus, K.H.; Blanchard, S.C.; Spahn, C.M. Structure and dynamics of the mammalian ribosomal pretranslocation complex. Mol. Cell 2011, 44, 214–224. [Google Scholar] [CrossRef] [PubMed]
- Fu, J.; Munro, J.B.; Blanchard, S.C.; Frank, J. Cryoelectron microscopy structures of the ribosome complex in intermediate states during tRNA translocation. Proc. Natl. Acad. Sci. USA 2011, 108, 4817–4821. [Google Scholar] [CrossRef] [PubMed]
- Agirrezabala, X.; Liao, H.Y.; Schreiner, E.; Fu, J.; Ortiz-Meoz, R.F.; Schulten, K.; Green, R.; Frank, J. Structural characterization of mRNA-tRNA translocation intermediates. Proc. Natl. Acad. Sci. USA 2012, 109, 6094–6099. [Google Scholar] [CrossRef] [PubMed]
- Taylor, D.J.; Nilsson, J.; Merrill, A.R.; Andersen, G.R.; Nissen, P.; Frank, J. Structures of modified eEF2 80S ribosome complexes reveal the role of GTP hydrolysis in translocation. EMBO J. 2007, 26, 2421–2431. [Google Scholar] [CrossRef] [PubMed]
- Anger, A.M.; Armache, J.P.; Berninghausen, O.; Habeck, M.; Subklewe, M.; Wilson, D.N.; Beckmann, R. Structures of the human and Drosophila 80S ribosome. Nature 2013, 497, 80–85. [Google Scholar] [CrossRef] [PubMed]
- Schuwirth, B.S.; Borovinskaya, M.A.; Hau, C.W.; Zhang, W.; Vila-Sanjurjo, A.; Holton, J.M.; Cate, J.H. Structures of the bacterial ribosome at 3.5 Å resolution. Science 2005, 310, 827–834. [Google Scholar]
- Ratje, A.H.; Loerke, J.; Mikolajka, A.; Brunner, M.; Hildebrand, P.W.; Starosta, A.L.; Donhofer, A.; Connell, S.R.; Fucini, P.; Mielke, T.; et al. Head swivel on the ribosome facilitates translocation by means of intra-subunit tRNA hybrid sites. Nature 2010, 468, 713–716. [Google Scholar]
- Guo, Z.; Noller, H.F. Rotation of the head of the 30S ribosomal subunit during mRNA translocation. Proc. Natl. Acad. Sci. USA 2012, 109, 20391–20394. [Google Scholar] [CrossRef] [PubMed]
- Mohan, S.; Donohue, J.P.; Noller, H.F. Molecular mechanics of 30S subunit head rotation. Proc. Natl. Acad. Sci. USA 2014, 111, 13325–13330. [Google Scholar] [CrossRef] [PubMed]
- Lin, J.; Gagnon, M.G.; Bulkley, D.; Steitz, T.A. Conformational changes of elongation factor G on the ribosome during tRNA translocation. Cell 2015, 160, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Salsi, E.; Farah, E.; Netter, Z.; Dann, J.; Ermolenko, D.N. Movement of elongation factor G between compact and extended conformations. J. Mol. Biol. 2015, 427, 454–467. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.G.; Selmer, M.; Dunham, C.M.; Weixlbaumer, A.; Kelley, A.C.; Ramakrishnan, V. The structure of the ribosome with elongation factor G trapped in the posttranslocational state. Science 2009, 326, 694–699. [Google Scholar] [CrossRef] [PubMed]
- Khade, P.K.; Joseph, S. Messenger RNA interactions in the decoding center control the rate of translocation. Nat. Struct. Mol. Biol. 2011, 18, 1300–1302. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; Song, G.; Zhang, D.; Li, Z.; Lyu, Z.; Dong, J.; Achenbach, J.; Gong, W.; Zhao, X.S.; Nierhaus, K.H.; et al. EF-G catalyzes tRNA translocation by disrupting interactions between decoding center and codon-anticodon duplex. Nat. Struct. Mol. Biol. 2014, 21, 817–824. [Google Scholar]
- Rodnina, M.V.; Savelsbergh, A.; Katunin, V.I.; Wintermeyer, W. Hydrolysis of GTP by elongation factor G drives tRNA movement on the ribosome. Nature 1997, 385, 37–41. [Google Scholar] [CrossRef] [PubMed]
- Peske, F.; Matassova, N.B.; Savelsbergh, A.; Rodnina, M.V.; Wintermeyer, W. Conformationally restricted elongation factor G retains GTPase activity but is inactive in translocation on the ribosome. Mol. Cell 2000, 6, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Savelsbergh, A.; Katunin, V.I.; Mohr, D.; Peske, F.; Rodnina, M.V.; Wintermeyer, W. An elongation factor G-induced ribosome rearrangement precedes tRNA-mRNA translocation. Mol. Cell 2003, 11, 1517–1523. [Google Scholar] [CrossRef] [PubMed]
- Wilden, B.; Savelsbergh, A.; Rodnina, M.V.; Wintermeyer, W. Role and timing of GTP binding and hydrolysis during EF-G-dependent tRNA translocation on the ribosome. Proc. Natl. Acad. Sci. USA 2006, 103, 13670–13675. [Google Scholar] [CrossRef] [PubMed]
- Caliskan, N.; Katunin, V.I.; Belardinelli, R.; Peske, F.; Rodnina, M.V. Programmed–1 frameshifting by kinetic partitioning during impeded translocation. Cell 2014, 157, 1619–1631. [Google Scholar] [CrossRef] [PubMed]
- Spiegel, P.C.; Ermolenko, D.N.; Noller, H.F. Elongation factor G stabilizes the hybrid-state conformation of the 70s ribosome. RNA 2007, 13, 1473–1482. [Google Scholar] [CrossRef] [PubMed]
- Munro, J.B.; Wasserman, M.R.; Altman, R.B.; Wang, L.; Blanchard, S.C. Correlated conformational events in EF-G and the ribosome regulate translocation. Nat. Struct. Mol. Biol. 2010, 17, 1470–1477. [Google Scholar] [CrossRef] [PubMed]
- Ermolenko, D.N.; Noller, H.F. mRNA translocation occurs during the second step of ribosomal intersubunit rotation. Nat. Struct. Mol. Biol. 2011, 18, 457–462. [Google Scholar] [CrossRef] [PubMed]
- Jenner, L.B.; Demeshkina, N.; Yusupova, G.; Yusupov, M. Structural aspects of messenger RNA reading frame maintenance by the ribosome. Nat. Struct. Mol. Biol. 2010, 17, 555–560. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lancaster, L.; Donohue, J.P.; Noller, H.F. How the ribosome hands the A-site tRNA to the P site during EF-G-catalyzed translocation. Science 2014, 345, 1188–1191. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Lancaster, L.; Donohue, J.P.; Noller, H.F. Crystal structures of EF-G-ribosome complexes trapped in intermediate states of translocation. Science 2013, 340, 1236086. [Google Scholar] [CrossRef] [PubMed]
- Ramrath, D.J.; Lancaster, L.; Sprink, T.; Mielke, T.; Loerke, J.; Noller, H.F.; Spahn, C.M. Visualization of two transfer RNAs trapped in transit during elongation factor G-mediated translocation. Proc. Natl. Acad. Sci. USA 2013, 110, 20964–20969. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Feng, S.; Kumar, V.; Ero, R.; Gao, Y.G. Structure of EF-G-ribosome complex in a pretranslocation state. Nat. Struct. Mol. Biol. 2013, 20, 1077–1084. [Google Scholar] [CrossRef] [PubMed]
- Pulk, A.; Cate, J.H. Control of ribosomal subunit rotation by elongation factor G. Science 2013, 340, 1235970. [Google Scholar] [CrossRef] [PubMed]
- Brilot, A.F.; Korostelev, A.A.; Ermolenko, D.N.; Grigorieff, N. Structure of the ribosome with elongation factor G trapped in the pretranslocation state. Proc. Natl Acad. Sci. USA 2013, 110, 20994–20999. [Google Scholar] [CrossRef] [PubMed]
- Samaha, R.R.; Green, R.; Noller, H.F. A base pair between tRNA and 23S rRNA in the peptidyl transferase centre of the ribosome. Nature 1995, 377, 309–314. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.F.; Green, R. Base-pairing between 23s rRNA and tRNA in the ribosomal A site. Mol. Cell 1999, 4, 859–864. [Google Scholar] [CrossRef] [PubMed]
- Hopper, A.K.; Phizicky, E.M. tRNA transfers to the limelight. Genes Dev. 2003, 17, 162–180. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, R.; Chen, S.; Dare, K.; Gilreath, M.; Praetorius-Ibba, M.; Raina, M.; Reynolds, N.M.; Rogers, T.; Roy, H.; Yadavalli, S.S.; et al. tRNAs: Cellular barcodes for amino acids. FEBS Lett. 2010, 584, 387–395. [Google Scholar]
- Greber, B.J.; Boehringer, D.; Leibundgut, M.; Bieri, P.; Leitner, A.; Schmitz, N.; Aebersold, R.; Ban, N. The complete structure of the large subunit of the mammalian mitochondrial ribosome. Nature 2014, 515, 283–286. [Google Scholar] [PubMed]
- Pesavento, C.; Hengge, R. Bacterial nucleotide-based second messengers. Curr. Opin. Microbiol. 2009, 12, 170–176. [Google Scholar] [CrossRef] [PubMed]
- Stent, G.S.; Brenner, S. A genetic locus for the regulation of ribonucleic acid synthesis. Proc. Natl. Acad. Sci. USA 1961, 47, 2005–2014. [Google Scholar] [CrossRef] [PubMed]
- Potrykus, K.; Cashel, M. (p)ppGpp: Still magical? Annu. Rev. Microbiol. 2008, 62, 35–51. [Google Scholar] [CrossRef] [PubMed]
- Srivatsan, A.; Wang, J.D. Control of bacterial transcription, translation and replication by (p)ppGpp. Curr. Opin. Microbiol. 2008, 11, 100–105. [Google Scholar] [CrossRef] [PubMed]
- Starosta, A.L.; Lassak, J.; Jung, K.; Wilson, D.N. The bacterial translation stress response. FEMS Microbiol. Rev. 2014, 38, 1172–1201. [Google Scholar] [CrossRef] [PubMed]
- Yegian, C.D.; Stent, G.S.; Martin, E.M. Intracellular condition of Escherichia coli transfer RNA. Proc. Natl. Acad. Sci. USA 1966, 55, 839–846. [Google Scholar] [CrossRef] [PubMed]
- Haseltine, W.A.; Block, R. Synthesis of guanosine tetra- and pentaphosphate requires the presence of a codon-specific, uncharged transfer ribonucleic acid in the acceptor site of ribosomes. Proc. Natl. Acad. Sci. USA 1973, 70, 1564–1568. [Google Scholar] [CrossRef] [PubMed]
- Agirrezabala, X.; Fernandez, I.S.; Kelley, A.C.; Carton, D.G.; Ramakrishnan, V.; Valle, M. The ribosome triggers the stringent response by RelA via a highly distorted tRNA. EMBO Rep. 2013, 14, 811–816. [Google Scholar] [CrossRef] [PubMed]
- Gropp, M.; Strausz, Y.; Gross, M.; Glaser, G. Regulation of Escherichia coli RelA requires oligomerization of the C-terminal domain. J. Bacteriol. 2001, 183, 570–579. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Ishiguro, E.E. Involvement of the N-terminus of ribosomal protein L11 in regulation of the RelA protein of Escherichia coli. J. Bacteriol. 2001, 183, 6532–6537. [Google Scholar] [CrossRef] [PubMed]
- Wendrich, T.M.; Blaha, G.; Wilson, D.N.; Marahiel, M.A.; Nierhaus, K.H. Dissection of the mechanism for the stringent factor RelA. Mol. Cell 2002, 10, 779–788. [Google Scholar] [CrossRef] [PubMed]
- Avarbock, D.; Avarbock, A.; Rubin, H. Differential regulation of opposing RelMtb activities by the aminoacylation state of a tRNA·Ribosome·mRNA·RelMtb complex. Biochemistry 2000, 39, 11640–11648. [Google Scholar] [CrossRef] [PubMed]
- Avarbock, A.; Avarbock, D.; Teh, J.S.; Buckstein, M.; Wang, Z.M.; Rubin, H. Functional regulation of the opposing (p)ppGpp synthetase/hydrolase activities of RelMtb from Mycobacterium tuberculosis. Biochemistry 2005, 44, 9913–9923. [Google Scholar] [CrossRef] [PubMed]
- Jain, V.; Saleem-Batcha, R.; China, A.; Chatterji, D. Molecular dissection of the mycobacterial stringent response protein Rel. Protein Sci. 2006, 15, 1449–1464. [Google Scholar] [CrossRef] [PubMed]
- Schreiber, G.; Metzger, S.; Aizenman, E.; Roza, S.; Cashel, M.; Glaser, G. Overexpression of the relA gene in Escherichia coli. J. Biol. Chem. 1991, 266, 3760–3767. [Google Scholar] [PubMed]
- Hashem, Y.; des Georges, A.; Dhote, V.; Langlois, R.; Liao, H.Y.; Grassucci, R.A.; Pestova, T.V.; Hellen, C.U.; Frank, J. Hepatitis-C-virus-like internal ribosome entry sites displace eIF3 to gain access to the 40S subunit. Nature 2013, 503, 539–543. [Google Scholar] [CrossRef] [PubMed]
- Yamamoto, H.; Unbehaun, A.; Loerke, J.; Behrmann, E.; Collier, M.; Burger, J.; Mielke, T.; Spahn, C.M. Structure of the mammalian 80S initiation complex with initiation factor 5B on HCV-IRES RNA. Nat. Struct. Mol. Biol. 2014, 21, 721–727. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Korostelev, A.; Lancaster, L.; Noller, H.F. Crystal structures of 70s ribosomes bound to release factors RF1, RF2 and RF3. Curr. Opin. Struct. Biol. 2012, 22, 733–742. [Google Scholar] [CrossRef] [PubMed]
- Dunkle, J.A.; Wang, L.; Feldman, M.B.; Pulk, A.; Chen, V.B.; Kapral, G.J.; Noeske, J.; Richardson, J.S.; Blanchard, S.C.; Cate, J.H. Structures of the bacterial ribosome in classical and hybrid states of tRNA binding. Science 2011, 332, 981–984. [Google Scholar] [CrossRef] [PubMed]
- Ramrath, D.J.; Yamamoto, H.; Rother, K.; Wittek, D.; Pech, M.; Mielke, T.; Loerke, J.; Scheerer, P.; Ivanov, P.; Teraoka, Y.; et al. The complex of tmRNA–SmpB and EF-G on translocating ribosomes. Nature 2012, 485, 526–529. [Google Scholar]
- Munro, J.B.; Sanbonmatsu, K.Y.; Spahn, C.M.; Blanchard, S.C. Navigating the ribosomeʼs metastable energy landscape. Trends Biochem. Sci. 2009, 34, 390–400. [Google Scholar] [CrossRef] [PubMed]
- Frank, J.; Gonzalez, R.L., Jr. Structure and dynamics of a processive Brownian motor: The translating ribosome. Annu. Rev. Biochem. 2010, 79, 381–412. [Google Scholar] [CrossRef] [PubMed]
- Moore, P.B. How should we think about the ribosome? Annu. Rev. Biophys. 2012, 41, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Melnikov, S.; Ben-Shem, A.; de Loubresse, N.G.; Jenner, L.; Yusupova, G.; Yusupov, M. One core, two shells: Bacterial and eukaryotic ribosomes. Nat. Struct. Mol. Biol. 2012, 19, 560–567. [Google Scholar] [CrossRef] [PubMed]
- Klinge, S.; Voigts-Hoffmann, F.; Leibundgut, M.; Ban, N. Atomic structures of the eukaryotic ribosome. Trends Biochem. Sci. 2012, 37, 189–198. [Google Scholar] [CrossRef] [PubMed]
- Yusupova, G.; Yusupov, M. High-resolution structure of the eukaryotic 80S ribosome. Annu. Rev. Biochem. 2014, 83, 467–486. [Google Scholar] [CrossRef] [PubMed]
- Garreau de Loubresse, N.; Prokhorova, I.; Holtkamp, W.; Rodnina, M.V.; Yusupova, G.; Yusupov, M. Structural basis for the inhibition of the eukaryotic ribosome. Nature 2014, 513, 517–522. [Google Scholar] [CrossRef] [PubMed]
- Ramakrishnan, V. The ribosome emerges from a black box. Cell 2014, 159, 979–984. [Google Scholar] [CrossRef] [PubMed]
- Bai, X.C.; McMullan, G.; Scheres, S.H. How cryo-EM is revolutionizing structural biology. Trends Biochem. Sci. 2015, 40, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Shaikh, T.R.; Yassin, A.S.; Lu, Z.; Barnard, D.; Meng, X.; Lu, T.M.; Wagenknecht, T.; Agrawal, R.K. Initial bridges between two ribosomal subunits are formed within 9.4 milliseconds, as studied by time-resolved cryo-EM. Proc. Natl. Acad. Sci. USA 2014, 111, 9822–9827. [Google Scholar] [CrossRef]
- Liu, T.; Kaplan, A.; Alexander, L.; Yan, S.; Wen, J.D.; Lancaster, L.; Wickersham, C.E.; Fredrik, K.; Noller, H.; Tinoco, I.; et al. Direct measurement of the mechanical work during translocation by the ribosome. Elife 2014, 3, e03406. [Google Scholar]
- Yao, L.; Li, Y.; Tsai, T.W.; Xu, S.; Wang, Y. Noninvasive measurement of the mechanical force generated by motor protein EF-G during ribosome translocation. Angew. Chem. Int. Ed. 2013, 52, 14041–14044. [Google Scholar] [CrossRef]
- Dashti, A.; Schwander, P.; Langlois, R.; Fung, R.; Li, W.; Hosseinizadeh, A.; Liao, H.Y.; Pallesen, J.; Sharma, G.; Stupina, V.A.; et al. Trajectories of the ribosome as a Brownian nanomachine. Proc. Natl. Acad. Sci. USA 2014, 111, 17492–17497. [Google Scholar] [CrossRef]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Agirrezabala, X.; Valle, M. Structural Insights into tRNA Dynamics on the Ribosome. Int. J. Mol. Sci. 2015, 16, 9866-9895. https://doi.org/10.3390/ijms16059866
Agirrezabala X, Valle M. Structural Insights into tRNA Dynamics on the Ribosome. International Journal of Molecular Sciences. 2015; 16(5):9866-9895. https://doi.org/10.3390/ijms16059866
Chicago/Turabian StyleAgirrezabala, Xabier, and Mikel Valle. 2015. "Structural Insights into tRNA Dynamics on the Ribosome" International Journal of Molecular Sciences 16, no. 5: 9866-9895. https://doi.org/10.3390/ijms16059866