
Synergistic and Antagonistic Action of Phytochrome (Phy) A and PhyB during Seedling De-Etiolation in Arabidopsis thaliana

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results and Discussion
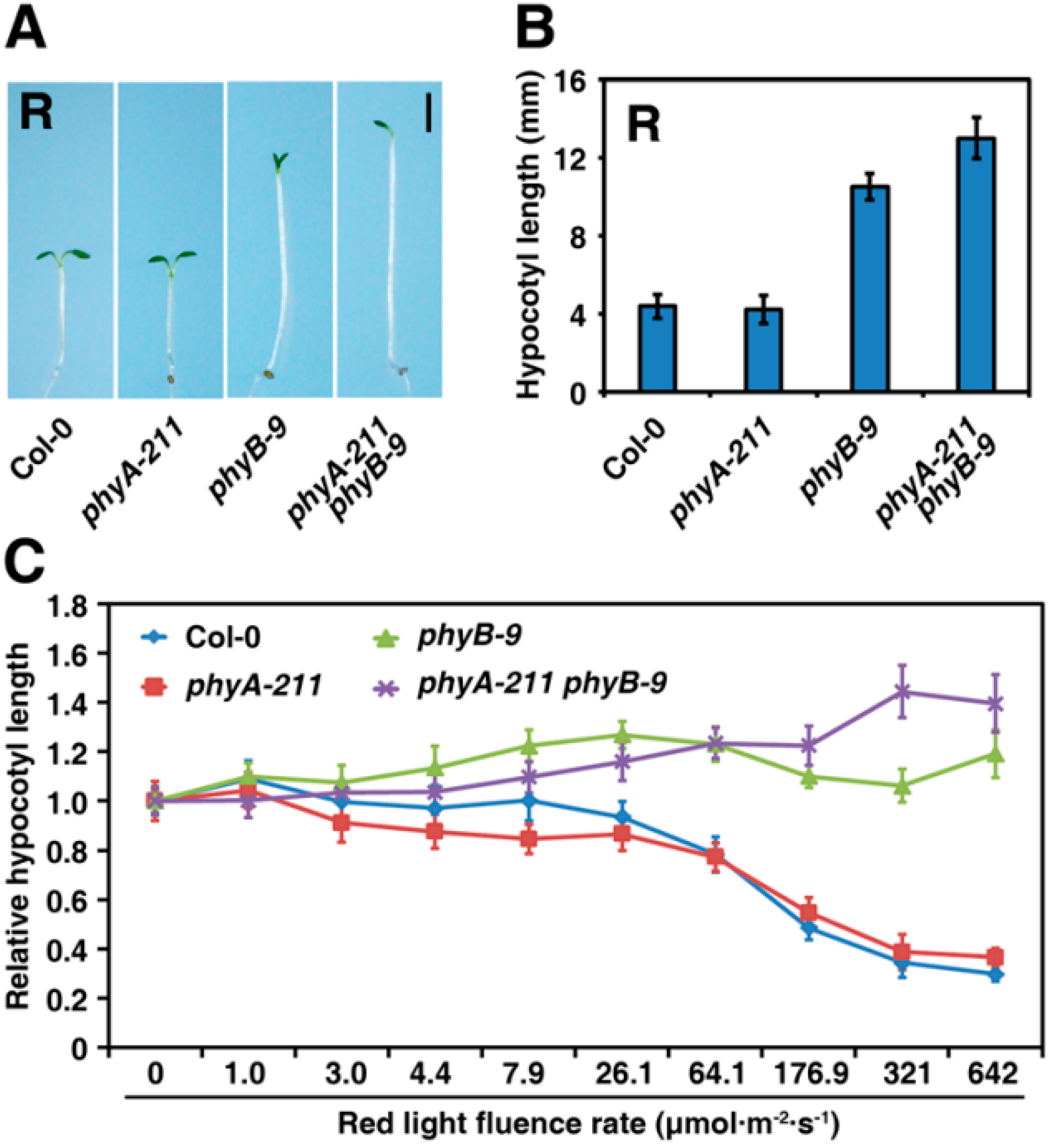
2.1. PhyA and PhyB Function Coordinately to Repress Hypocotyl Elongation under R Light

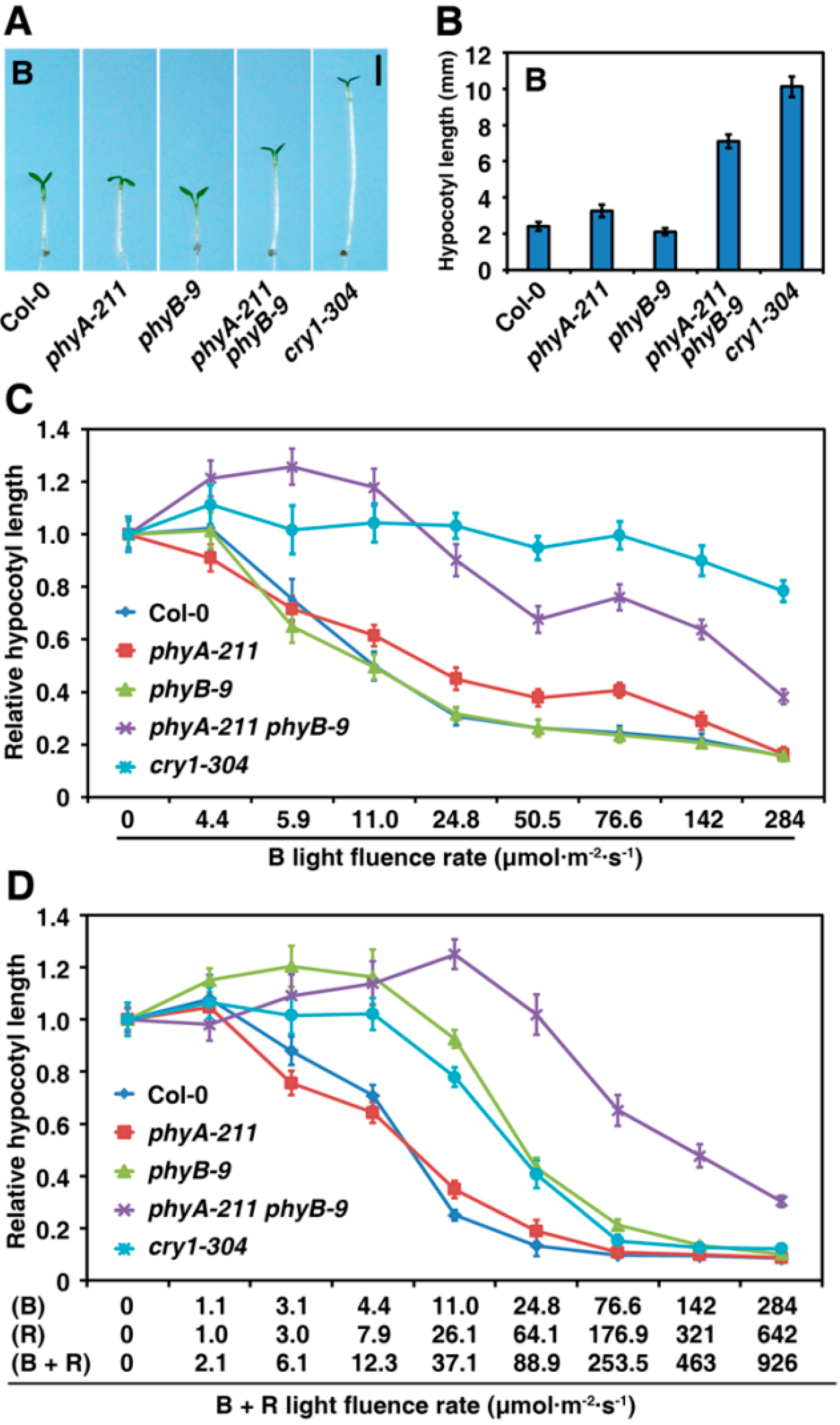
2.2. PhyA and PhyB Synergistically Inhibit Hypocotyl Elongation under B Light

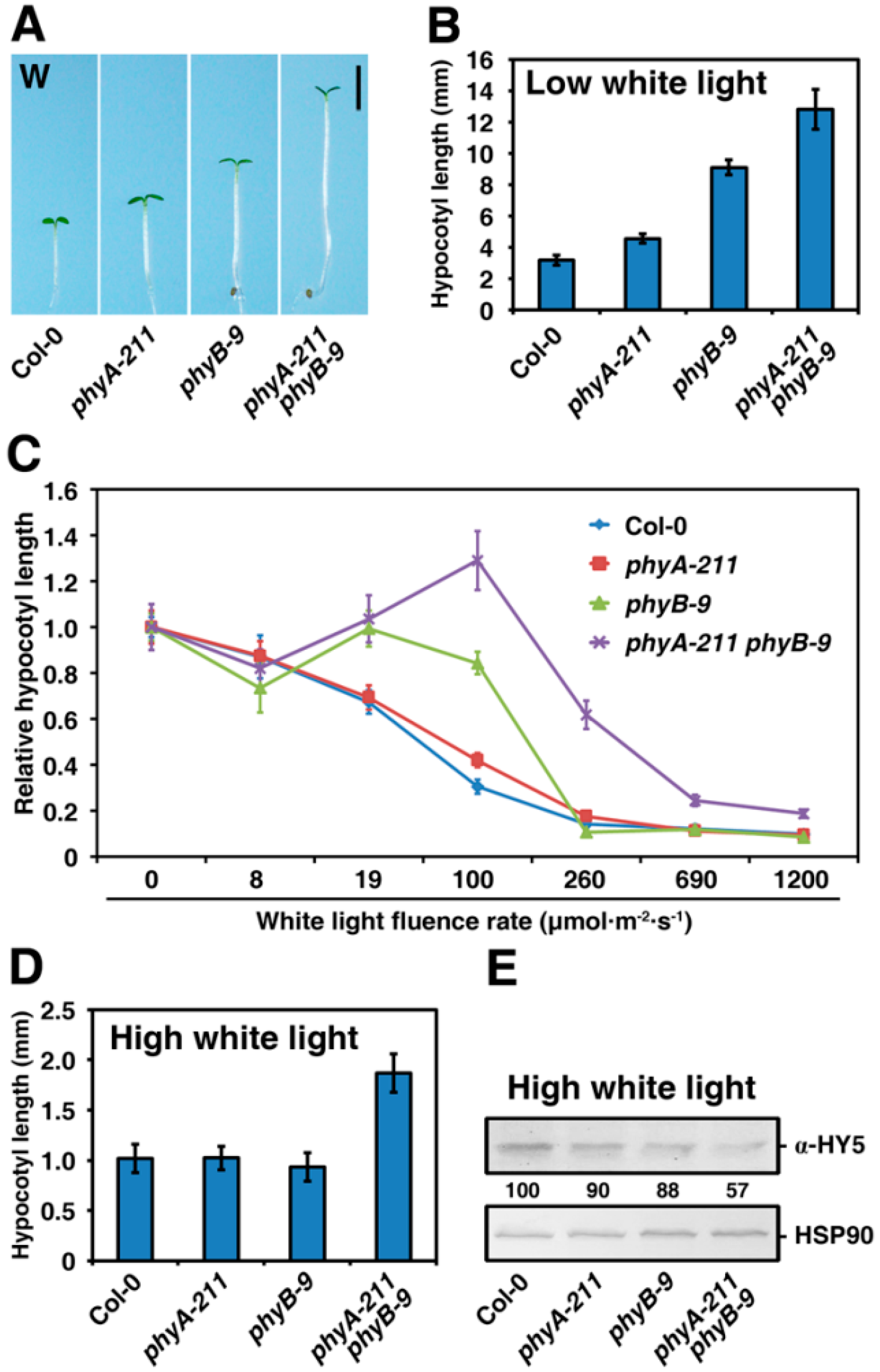
2.3. PhyA and PhyB Synergistically Promote De-Etiolation under W Light Condition

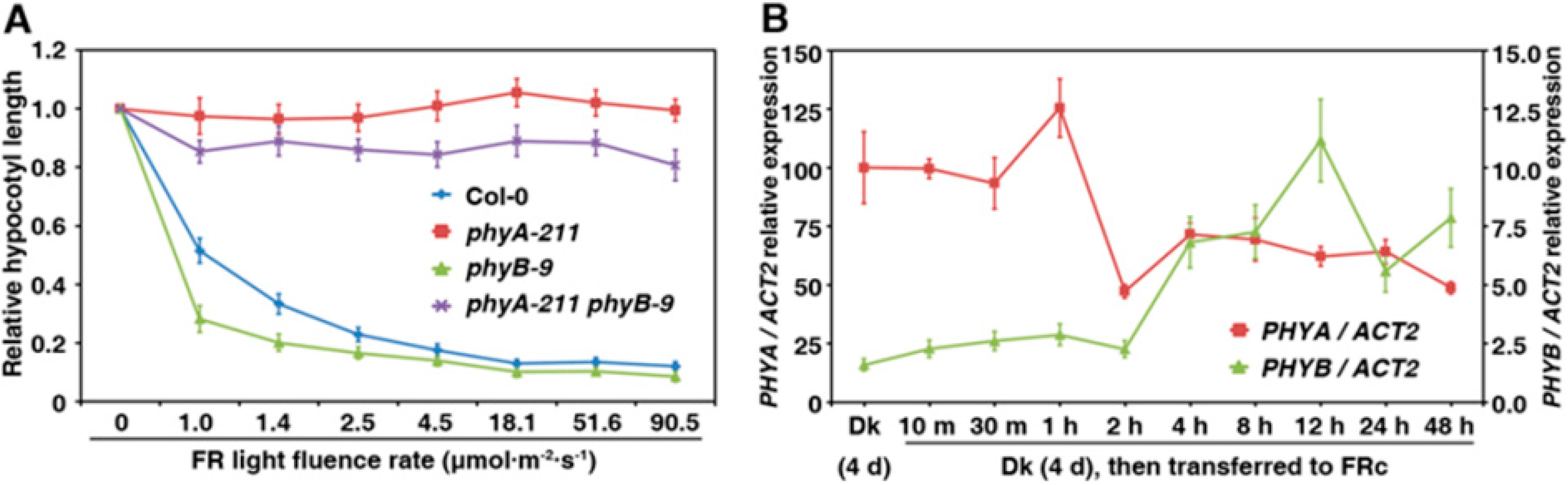
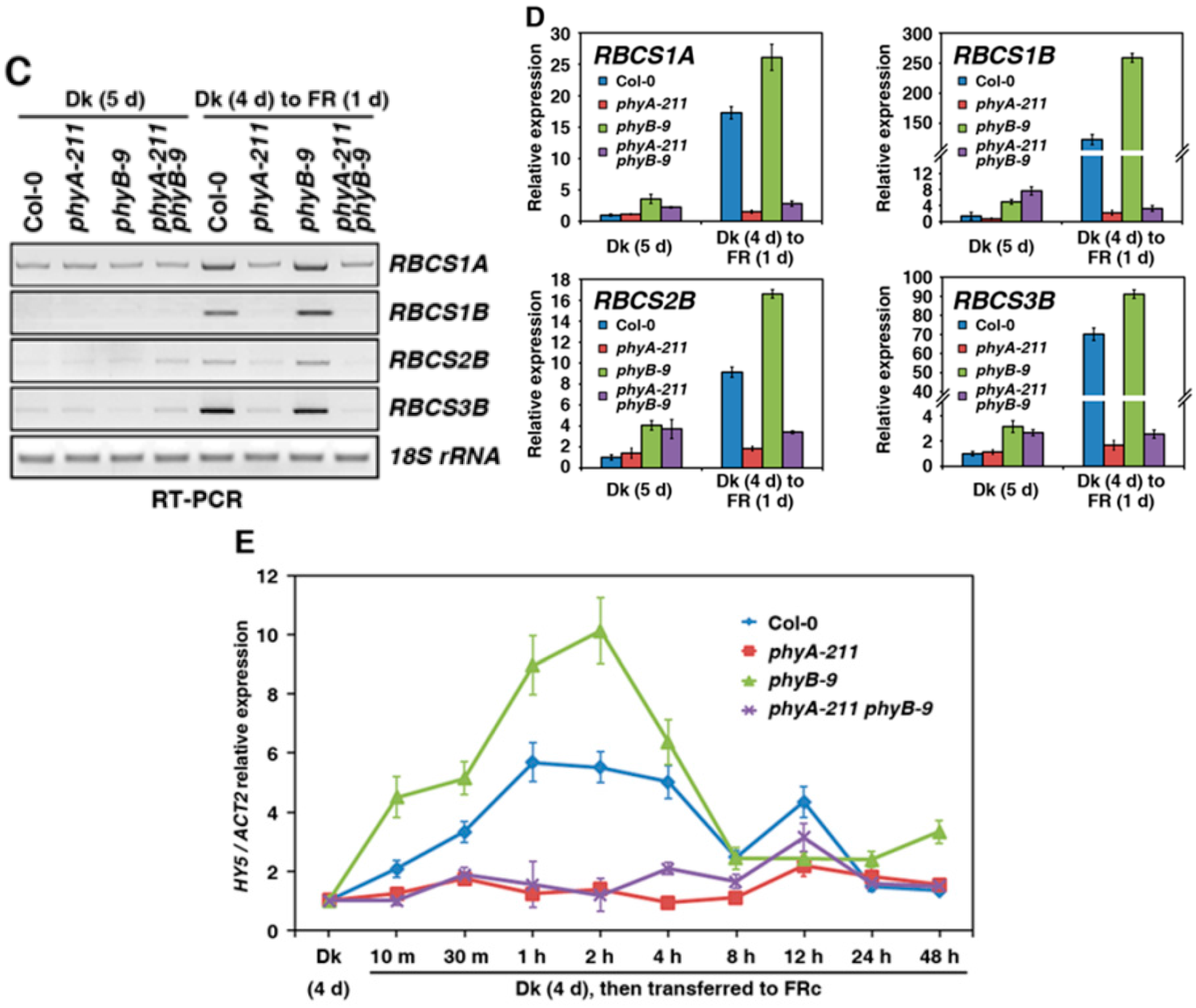
2.4. PhyB Acts in Opposition to PhyA on Seedling Development under FR Light


3. Experimental Section
3.1. Plant Materials and Growth Conditions
3.2. Construction of Double Mutant
3.3. Measurement of Hypocotyl Length and Data Analysis
3.4. Immunoblot Analysis
3.5. RT-PCR and Real Time Quantitative RT-PCR Analyses
3.6. Accession Numbers
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- McNellis, T.W.; Deng, X.W. Light control of seedling morphogenetic pattern. Plant Cell 1995, 7, 1749–1761. [Google Scholar] [CrossRef] [PubMed]
- Zhong, S.; Shi, H.; Xue, C.; Wang, L.; Xi, Y.; Li, J.; Quail, P.H.; Deng, X.W.; Guo, H. A molecular framework of light-controlled phytohormone action in Arabidopsis. Curr. Biol. 2012, 22, 1530–1535. [Google Scholar] [CrossRef] [PubMed]
- Kendrick, R.E.; Kronenberg, G.H.M. Photomorphogenesis in Plants; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1994. [Google Scholar]
- Briggs, W.R.; Olney, M.A. Photoreceptors in plant photomorphogenesis to date: Five phytochromes, two cryptochromes, one phototropin, and one superchrome. Plant Physiol. 2001, 125, 85–88. [Google Scholar] [CrossRef] [PubMed]
- Lin, C. Blue light receptors and signal transduction. Plant Cell 2002, 14, S207–S225. [Google Scholar] [PubMed]
- Neff, M.M.; Fankhauser, C.; Chory, J. Light: An indicator of time and place. Genes Dev. 2000, 14, 257–271. [Google Scholar] [PubMed]
- Nagatani, A.; Reed, J.W.; Chory, J. Isolation and initial characterization of Arabidopsis mutants that are deficient in phytochrome A. Plant Physiol. 1993, 102, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Whitelam, G.C.; Johnson, E.; Peng, J.; Carol, P.; Anderson, M.L.; Cowl, J.S.; Harberd, N.P. Phytochrome A null mutants of Arabidopsis display a wild-type phenotype in white light. Plant Cell 1993, 5, 757–768. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.W.; Nagatani, A.; Elich, T.D.; Fagan, M.; Chory, J. Phytochrome A and phytochrome B have overlapping but distinct functions in Arabidopsis development. Plant Physiol. 1994, 104, 1139–1149. [Google Scholar] [PubMed]
- Botto, J.F.; Sanchez, R.A.; Whitelam, G.C.; Casal, J.J. Phytochrome A mediates the promotion of seed germination by very low fluences of light and canopy shade light in Arabidopsis. Plant Physiol. 1996, 110, 439–444. [Google Scholar] [PubMed]
- Shinomura, T.; Nagatani, A.; Hanzawa, H.; Kubota, M.; Watanabe, M.; Furuya, M. Action spectra for phytochrome A- and B-specific photoinduction of seed germination in Arabidopsis thaliana. Proc. Natl. Acad. Sci. USA 1996, 93, 8129–8133. [Google Scholar] [CrossRef] [PubMed]
- Neff, M.M.; Chory, J. Genetic interactions between phytochrome A, phytochrome B, and cryptochrome 1 during Arabidopsis development. Plant Physiol. 1998, 118, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Koornneef, M.; Rolff, E.; Spruit, C.J.P. Genetic control of light-inhibited hypocotyl elongation in Arabidopsis thaliana (L.). Heynh. Z. Pflanzenphysiol. 1980, 100, 147–160. [Google Scholar] [CrossRef]
- Oyama, T.; Shimura, Y.; Okada, K. The Arabidopsis HY5 gene encodes a bZIP protein that regulates stimulus-induced development of root and hypocotyl. Genes Dev. 1997, 11, 2983–2995. [Google Scholar] [CrossRef] [PubMed]
- Osterlund, M.T.; Hardtke, C.; Wei, N.; Deng, X.W. Targeted destabilization of HY5 during light regulated development of Arabidopsis. Nature 2000, 405, 462–466. [Google Scholar] [PubMed]
- Ulm, R.; Baumann, A.; Oravecz, A.; Mate, Z.; Adam, E.; Oakeley, E.J.; Schafer, E.; Nagy, F. Genome-wide analysis of gene expression reveals function of the bZIP transcription factor HY5 in the UV-B response of Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 1397–1402. [Google Scholar]
- Spreitzer, R.J. Role of the small subunit in ribulose-1,5-bisphosphate carboxylase/oxygenase. Arch. Biochem. Biophys. 2003, 414, 141–149. [Google Scholar] [CrossRef]
- Masanori, I.; Honami, T.; Yuji, S.; Amane, M.; Hiroyuki, I. RBCS1A and RBCS3B, two major members within the Arabidopsis RBCS multigene family, function to yield sufficient Rubisco content for leaf photosynthetic capacity. J. Exp. Bot. 2012, 63, 2159–2170. [Google Scholar]
- Casal, J.J. Coupling of phytochrome B to the control of hypocotyl growth in Arabidopsis. Planta 1995, 196, 23–29. [Google Scholar] [CrossRef] [PubMed]
- Parks, B.M.; Spalding, E.P. Sequential and coordinated action of phytochromes A and B during Arabidopsis stem growth revealed by kinetic analysis. Proc. Natl. Acad. Sci. USA 1999, 96, 14142–14146. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A.; Allen, T.; Whitelam, G.C. Phytochrome A is an irradiance-dependent red light sensor. Plant J. 2007, 50, 108–117. [Google Scholar] [CrossRef] [PubMed]
- Casal, J.J.; Boccalandro, H. Co-action between phytochrome B and HY4 in Arabidopsis thaliana. Planta 1995, 197, 213–218. [Google Scholar] [CrossRef] [PubMed]
- Casal, J.J.; Mazzella, M.A. Conditional synergism between cryptochrome 1 and phytochrome B is shown by the analysis of phyA, phyB, and hy4 simple, double, and triple mutants in Arabidopsis. Plant Physiol. 1998, 118, 19–25. [Google Scholar] [CrossRef] [PubMed]
- Franklin, K.A.; Davis, S.J.; Stoddart, W.M.; Vierstra, R.D.; Whitelam, G.C. Mutant analyses define multiple roles for phytochrome C in Arabidopsis photomorphogenesis. Plant Cell 2003, 15, 1981–1989. [Google Scholar] [CrossRef] [PubMed]
- Hennig, L.; Funk, M.; Whitelam, G.C.; Schafer, E. Functional interaction of cryptochrome 1 and phytochrome D. Plant J. 1999, 20, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Devlin, P.F.; Patel, S.R.; Whitelam, G.C. Phytochrome E influences internode elongation and flowering time in Arabidopsis. Plant Cell 1998, 10, 1479–1487. [Google Scholar] [PubMed]
- Poppe, C.; Schäfer, E. Seed germination of Arabidopsis thaliana phyA/phyB double mutants is under phytochrome control. Plant Physiol. 1997, 114, 1487–1492. [Google Scholar] [CrossRef] [PubMed]
- Hennig, L.; Poppe, C.; Sweere, U.; Martin, A.; Schäfer, E. Negative interference of endogenous phytochrome B with phytochrome A function in Arabidopsis. Plant Physiol. 2001, 125, 1036–1044. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Koloszvari, M.; Quail, P.H. Two small spatially distinct regions of phytochrome B are required for efficient signaling rates. Plant Cell. 1996, 8, 859–871. [Google Scholar] [CrossRef] [PubMed]
- Short, T.W. Overexpression of Arabidopsis phytochrome B inhibits phytochrome A function in the presence of sucrose. Plant Physiol. 1999, 119, 1497–1506. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.; Wu, S.; Zhai, H.; Zhou, P.; Song, M.; Su, L.; Xi, Y.; Li, Z.; Cai, Y.; Meng, F.; et al. Arabidopsis phytochrome B promotes SPA1 nuclear accumulation to repress photomorphogenesis under far-red light. Plant Cell 2013, 25, 115–133. [Google Scholar] [CrossRef] [PubMed]
- Ahmad, M.; Cashmore, A.R. The blue-light receptor cryptochrome 1 shows functional dependence on phytochrome A or phytochrome B in Arabidopsis thaliana. Plant J. 1997, 11, 421–427. [Google Scholar] [CrossRef] [PubMed]
- Poppe, C.; Sweere, U.; Drumm-Herrel, H.; Schäfer, E. The blue light receptor cryptochrome 1 can act independently of phytochrome A and B in Arabidopsis thaliana. Plant J. 1998, 16, 465–471. [Google Scholar] [CrossRef] [PubMed]
- Shinomura, T.; Uchida, K.; Furuya, M. Elementary processes of photoperception by phytochrome A for high-irradiance response of hypocotyl elongation in Arabidopsis. Plant Physiol. 2000, 122, 147–156. [Google Scholar] [CrossRef] [PubMed]
- Bae, G.; Choi, G. Decoding of light signals by plant phytochromes and their interacting proteins. Annu. Rev. Plant Biol. 2008, 59, 281–311. [Google Scholar] [CrossRef] [PubMed]
- Borthwick, H.A.; Hendricks, S.B.; Parker, M.W.; Toole, E.H.; Toole, V.K. A reversible photoreaction controlling seed germination. Proc. Natl. Acad. Sci. USA 1952, 38, 662–666. [Google Scholar] [CrossRef] [PubMed]
- Sharrock, R.A.; Clack, T. Heterodimerization of type II phytochromes in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 11500–11505. [Google Scholar] [CrossRef] [PubMed]
- Reed, J.W.; Nagpal, P.; Poole, D.S.; Furuya, M.; Chory, J. Mutations in the gene for the red/far-red light receptor phytochrome B alter cell elongation and physiological responses throughout Arabidopsis development. Plant Cell 1993, 5, 147–157. [Google Scholar] [CrossRef] [PubMed]
- Mockler, T.C.; Guo, H.; Yang, H.; Duong, H.; Lin, C. Antagonistic action of Arabidopsis cryptochromes and phytochrome B in the regulation of floral induction. Development 1999, 126, 2073–2082. [Google Scholar] [PubMed]
- Zhou, P.; Song, M.; Yang, Q.; Su, L.; Hou, P.; Guo, L.; Zheng, X.; Xi, Y.; Meng, F.; Xiao, Y.; et al. Both PHYTOCHROME RAPIDLY REGULATED1 (PAR1) and PAR2 promote seedling photomorphogenesis in multiple light signaling pathways. Plant Physiol. 2014, 164, 841–852. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.; Xu, G.; Tang, W.; Jing, Y.; Ji, Q.; Fei, Z.; Lin, R. Antagonistic basic helix-loop-helix/bZIP transcription factors form transcriptional modules that integrate light and reactive oxygen species signaling in Arabidopsis. Plant Cell 2013, 25, 1657–1673. [Google Scholar] [CrossRef] [PubMed]
- Saijo, Y.; Zhu, D.; Li, J.; Rubio, V.; Zhou, Z.; Shen, Y.; Hoecker, U.; Wang, H.; Deng, X.W. Arabidopsis COP1/SPA1 complexand FHY1/FHY3 associate with distinct phosphorylated forms of phytochrome A in balancing light signaling. Mol. Cell 2008, 31, 607–613. [Google Scholar] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Su, L.; Hou, P.; Song, M.; Zheng, X.; Guo, L.; Xiao, Y.; Yan, L.; Li, W.; Yang, J. Synergistic and Antagonistic Action of Phytochrome (Phy) A and PhyB during Seedling De-Etiolation in Arabidopsis thaliana. Int. J. Mol. Sci. 2015, 16, 12199-12212. https://doi.org/10.3390/ijms160612199
Su L, Hou P, Song M, Zheng X, Guo L, Xiao Y, Yan L, Li W, Yang J. Synergistic and Antagonistic Action of Phytochrome (Phy) A and PhyB during Seedling De-Etiolation in Arabidopsis thaliana. International Journal of Molecular Sciences. 2015; 16(6):12199-12212. https://doi.org/10.3390/ijms160612199
Chicago/Turabian StyleSu, Liang, Pei Hou, Meifang Song, Xu Zheng, Lin Guo, Yang Xiao, Lei Yan, Wanchen Li, and Jianping Yang. 2015. "Synergistic and Antagonistic Action of Phytochrome (Phy) A and PhyB during Seedling De-Etiolation in Arabidopsis thaliana" International Journal of Molecular Sciences 16, no. 6: 12199-12212. https://doi.org/10.3390/ijms160612199