
In Vitro Expansion of CAG, CAA, and Mixed CAG/CAA Repeats


Abstract
:
1. Introduction
2. Results and Discussion
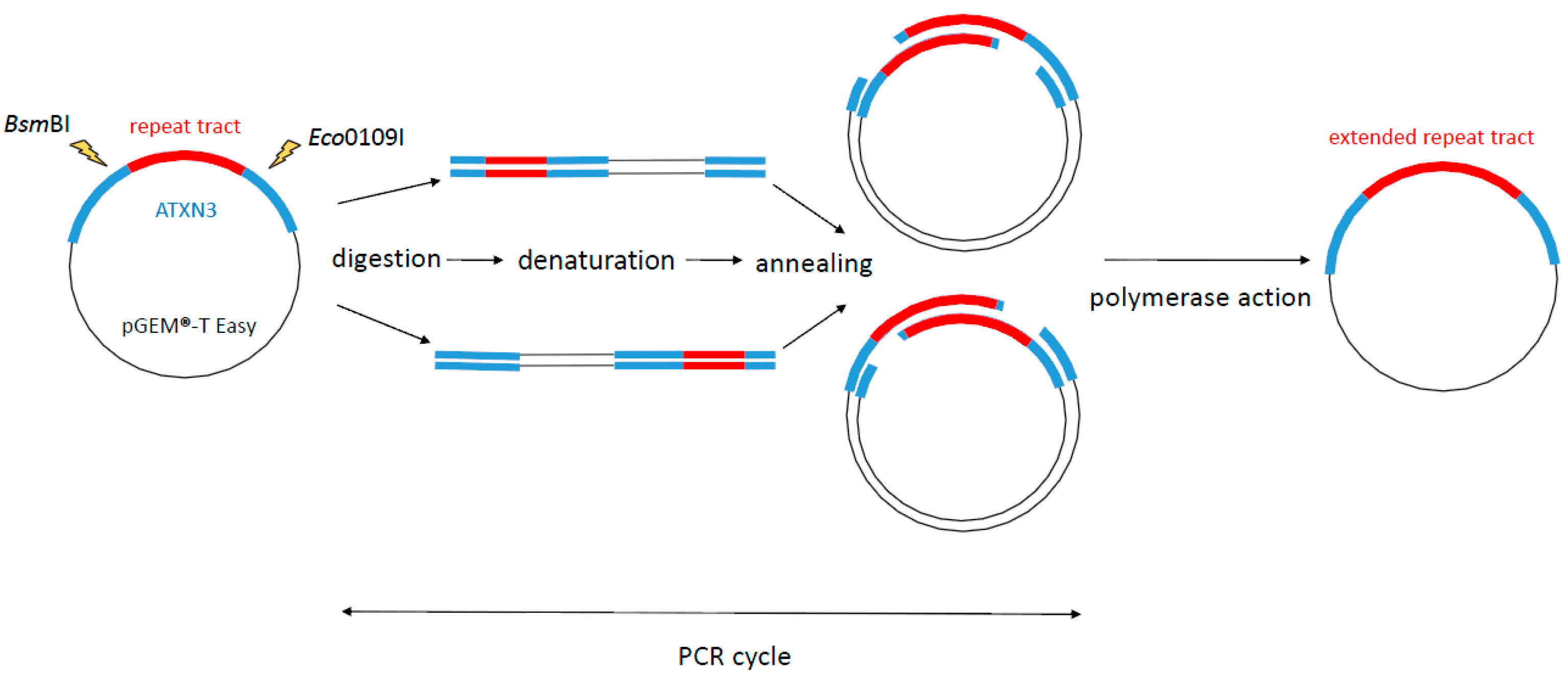
2.1. Experimental Design


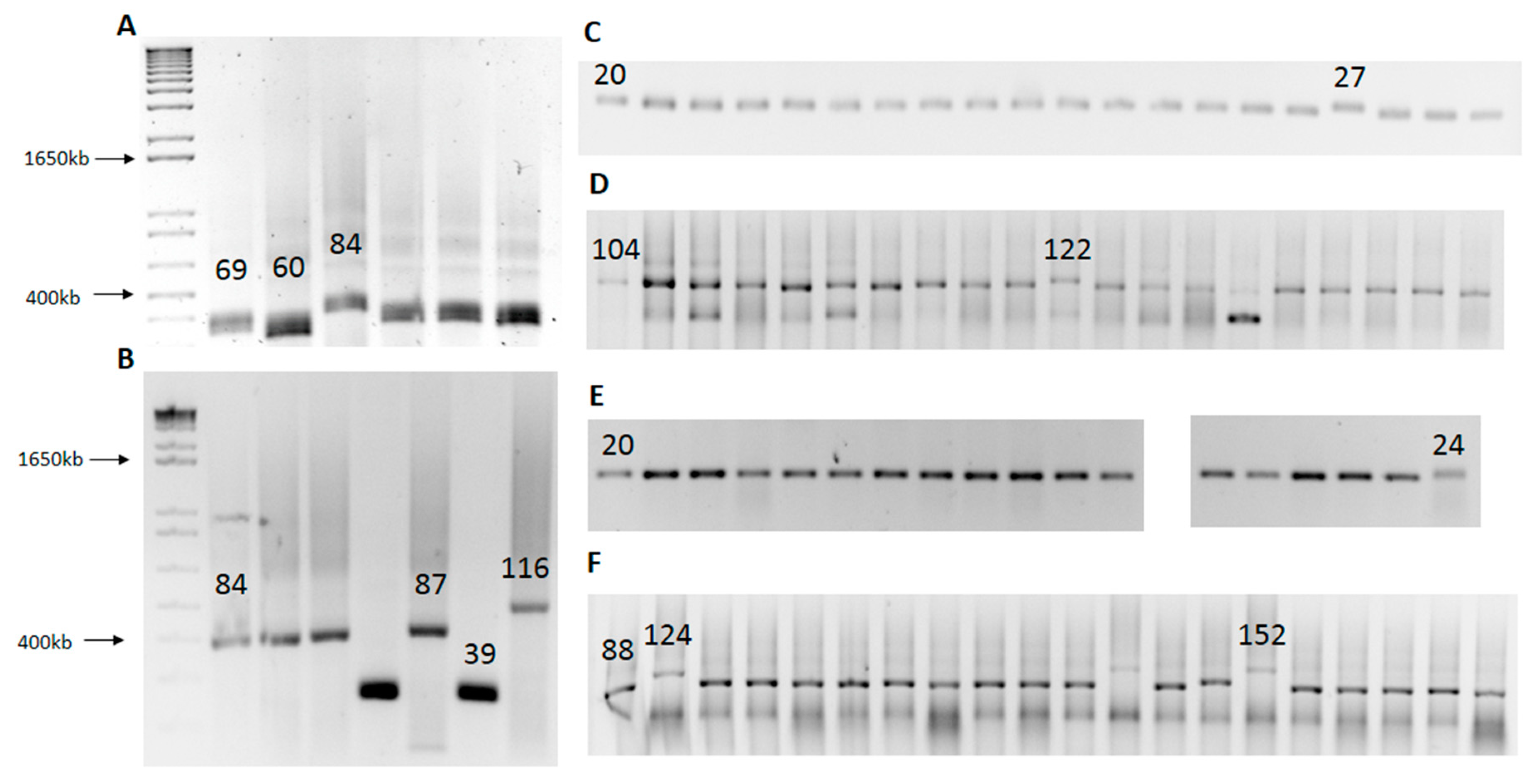
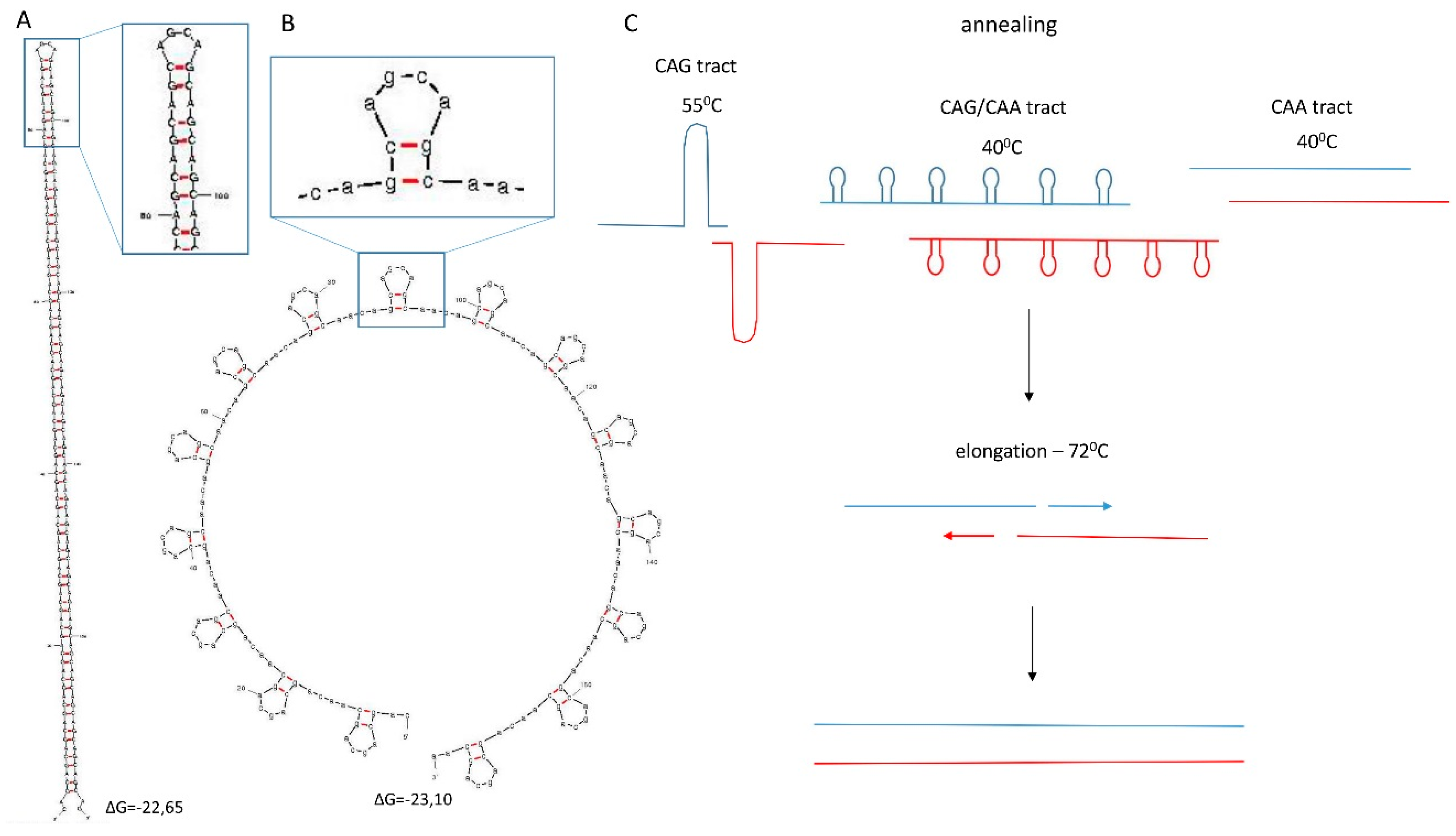
2.2. SLIP (Synthesis of Long Iterative Polynucleotide) Performance


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| CAG | L | S | Total | CAG/CAA | L | S | Total | CAA | L | S | Total |
|---|---|---|---|---|---|---|---|---|---|---|---|
| 69 | 3 | 3 | 25 | 20 | 1 | 0 | 85 | 20 | 2 | 0 | 95 |
| 84 | 2 | 2 | 18 | 24 | 1 | 1 | 95 | 25, 27 | 2 | 1 | 228 |
| 87, 116 | - | - | - | 28 | 2 | 0 | 114 | 30, 33 | 1 | 0 | 114 |
| 36 | 4 | 1 | 114 | 36 | 1 | 0 | 114 | ||||
| 7 | 2 | 114 | |||||||||
| 40, 44, 48, 52 | 2 | 3 | 114 | 53 | 2 | 2 | 228 | ||||
| 80, 88 | 7 | 2 | 114 | 58 | 1 | 2 | 114 | ||||
| 96, 108, 116, 124, 152 | - | - | - | 104 | 6 | 3 | 114 | ||||
| 1 | 5 | 114 | |||||||||
| 143, 150 | - | - | - | ||||||||
| 122 |

3. Experimental Section
3.1. SLIP (Synthesis of Long Iterative Polynucleotide)
3.2. Colony PCR
3.3. Preparation of Starting Constructs
| Repeated Motif | Oligonucleotide Sequence |
|---|---|
| (CAG)3CAA | 5ʹcggaagagacgagaagcctactttgaaaaacagcagcaaaagcagcaacgg (111 nt) |
| (CAG)3CAA | 5ʹgtcccgttgctgctgctgttgctgctgctgttgctgctgctgttgctgctgctgttgctgctgctgttgctgcttttgctgctgtttttcaaagtaggcttctcgtctct (107 nt) |
| CAA | 5ʹcggaagagacgagaagcctactttgaaaaacagcagcaaaagcagcaacgg (111 nt) |
| CAA | 5ʹgtcccgttgttgttgttgttgttgttgttgttgttgttgttgttgttgttgttgttgttgttgttgttgctgcttttgctgctgtttttcaaagtaggcttctcgtctct (107 nt) |
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- López Castel, A.; Cleary, J.D.; Pearson, C.E. Repeat instability as the basis for human diseases and as a potential target for therapy. Nat. Rev. Mol. Cell Biol. 2010, 11, 165–170. [Google Scholar] [CrossRef] [PubMed]
- Ashizawa, T.; Wells, R.D. Overview of the field. In Genetic Instabilities and Neurological Diseases, 2nd ed.; Wells, R.D., Ashizawa, T., Eds.; Elsevier: Burlington, MA, USA, 2006; pp. 3–17. [Google Scholar]
- Ranum, L.P.; Cooper, T.A. RNA-mediated neuromuscular disorders. Annu. Rev. Neurosci. 2006, 29, 259–277. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, T.M.; Thornton, C.A. Myotonic dystrophy: RNA-mediated muscle disease. Curr. Opin. Neurol. 2007, 20, 572–576. [Google Scholar] [CrossRef] [PubMed]
- Fiszer, A.; Krzyzosiak, W.J. RNA toxicity in polyglutamine disorders: Concepts, models, and progress of research. J. Mol. Med. 2013, 91, 683–691. [Google Scholar] [CrossRef] [PubMed]
- Ho, T.H.; Savkur, R.S.; Poulos, M.G.; Mancini, M.A.; Swanson, M.S.; Cooper, T.A. Colocalization of muscleblind with RNA foci is separable from mis-regulation of alternative splicing in myotonic dystrophy. J. Cell Sci. 2005, 118, 2923–2933. [Google Scholar] [CrossRef] [PubMed]
- Mykowska, A.; Sobczak, K.; Wojciechowska, M.; Kozlowski, P.; Krzyzosiak, W.J. CAG repeats mimic CUG repeats in the misregulation of alternative splicing. Nucleic Acids Res. 2011, 39, 8938–8951. [Google Scholar] [CrossRef] [PubMed]
- Bañez-Coronel, M.; Porta, S.; Kagerbauer, B.; Mateu-Huertas, E.; Pantano, L.; Ferrer, I.; Guzmán, M.; Estivill, X.; Martí, E. A pathogenic mechanism in huntington’s disease involves small CAG-repeated RNAs with neurotoxic activity. PLoS Genet. 2012, 8, e1002481. [Google Scholar] [CrossRef] [PubMed]
- Usdin, K. Bending the rules: Unusual nucleic acid structures and disease pathology in the repeat expansion diseases. In Genetic Instabilities and Neurological Diseases, 2nd ed.; Wells, R.D., Ashizawa, T., Eds.; Elsevier: Burlington, MA, USA, 2006; pp. 617–635. [Google Scholar]
- Mirkin, S.M. Replication of expandable DNA repeats. In Genetic Instabilities and Neurological Diseases, 2nd ed.; Wells, R.D., Ashizawa, T., Eds.; Elsevier: Burlington, MA, USA, 2006; pp. 637–644. [Google Scholar]
- Lin, Y.; Dion, V.; Wilson, J.H. Transcription and triplet repeat instability. In Genetic Instabilities and Neurological Diseases, 2nd ed.; Wells, R.D., Ashizawa, T., Eds.; Elsevier: Burlington, MA, USA, 2006; pp. 691–704. [Google Scholar]
- Ordway, J.M.; Detloff, P.J. In vitro synthesis and cloning of long CAG repeats. Biotechniques 1996, 21, 609–610, 612. [Google Scholar] [PubMed]
- Philips, A.V.; Timchenko, L.T.; Cooper, T.A. Disruption of splicing regulated by a CUG-binding protein in myotonic dystrophy. Science 1998, 280, 737–741. [Google Scholar] [CrossRef] [PubMed]
- Pujana, M.A.; Volpini, V.; Estivill, X. Large CAG/CTG repeat templates produced by PCR, usefulness for the DIRECT method of cloning genes with CAG/CTG repeat expansions. Nucleic Acids Res. 1998, 26, 1352–1353. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Sasagawa, N.; Suzuki, K.; Ishiura, S. Synthesis of long trinucleotide repeats in vitro. Neurosci. Lett. 1999, 262, 45–48. [Google Scholar] [CrossRef]
- Hsu, R.J.; Hsiao, K.M.; Lin, M.J.; Li, C.Y.; Wang, L.C.; Chen, L.K.; Pan, H. Long tract of untranslated CAG repeats is deleterious in transgenic mice. PLoS ONE 2011, 6, e16417. [Google Scholar] [CrossRef] [PubMed]
- Peters, M.F.; Ross, C.A. Preparation of human cDNAs encoding expanded polyglutamine repeats. Neurosci. Lett. 1999, 275, 129–132. [Google Scholar] [CrossRef]
- Scior, A.; Preissler, S.; Koch, M.; Deuerling, E. Directed PCR-free engineering of highly repetitive DNA sequences. BMC Biotechnol. 2011, 11, 87. [Google Scholar] [CrossRef] [PubMed]
- Michalik, A.; Kazantsev, A.; van Broeckhoven, C. Method to introduce stable, expanded, polyglutamine-encoding CAG/CAA trinucleotide repeats into CAG repeat-containing genes. Biotechniques 2001, 31, 250–252, 254. [Google Scholar] [PubMed]
- Nakamura, Y.; Gojobori, T.; Ikemura, T. Codon usage tabulated from international DNA sequence databases: Status for the year 2000. Nucleic Acids Res. 2000, 28, 292. [Google Scholar] [CrossRef] [PubMed]
- Imbert, G.; Saudou, F.; Yvert, G.; Devys, D.; Trottier, Y.; Garnier, J.M.; Weber, C.; Mandel, J.L.; Cancel, G.; Abbas, N.; et al. Cloning of the gene for spinocerebellar ataxia 2 reveals a locus with high sensitivity to expanded CAG/glutamine repeats. Nat. Genet. 1996, 14, 285–291. [Google Scholar] [CrossRef] [PubMed]
- Pulst, S.M.; Nechiporuk, A.; Nechiporuk, T.; Gispert, S.; Chen, X.N.; Lopes-Cendes, I.; Pearlman, S.; Starkman, S.; Orozco-Diaz, G.; Lunkes, A.; et al. Moderate expansion of a normally biallelic trinucleotide repeat in spinocerebellar ataxia type 2. Nat. Genet. 1996, 14, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Sanpei, K.; Takano, H.; Igarashi, S.; Sato, T.; Oyake, M.; Sasaki, H.; Wakisaka, A.; Tashiro, K.; Ishida, Y.; Ikeuchi, T.; et al. Identification of the spinocerebellar ataxia type 2 gene using a direct identification of repeat expansion and cloning technique, DIRECT. Nat. Genet. 1996, 14, 277–284. [Google Scholar] [CrossRef] [PubMed]
- Cancel, G.; Dürr, A.; Didierjean, O.; Imbert, G.; Bürk, K.; Lezin, A.; Belal, S.; Benomar, A.; Abada-Bendib, M.; Vial, C.; et al. Molecular and clinical correlations in spinocerebellar ataxia 2: A study of 32 families. Hum. Mol. Genet. 1997, 6, 709–715. [Google Scholar] [CrossRef] [PubMed]
- Giunti, P.; Sabbadini, G.; Sweeney, M.G.; Davis, M.B.; Veneziano, L.; Mantuano, E.; Federico, A.; Plasmati, R.; Frontali, M.; Wood, N.W. The role of the SCA2 trinucleotide repeat expansion in 89 autosomal dominant cerebellar ataxia families. Frequency, clinical and genetic correlates. Brain 1998, 121 Pt 3, 459–467. [Google Scholar] [CrossRef] [PubMed]
- Mizushima, K.; Watanabe, M.; Kondo, I.; Okamoto, K.; Shizuka, M.; Abe, K.; Aoki, M.; Shoji, M. Analysis of spinocerebellar ataxia type 2 gene and haplotype analysis: (CCG)1–2 polymorphism and contribution to founder effect. J. Med. Genet. 1999, 36, 112–114. [Google Scholar] [PubMed]
- Pujana, M.A.; Corral, J.; Gratacòs, M.; Combarros, O.; Berciano, J.; Genís, D.; Banchs, I.; Estivill, X.; Volpini, V. Spinocerebellar ataxias in spanish patients: Genetic analysis of familial and sporadic cases. The ataxia study group. Hum. Genet. 1999, 104, 516–522. [Google Scholar] [CrossRef] [PubMed]
- Choudhry, S.; Mukerji, M.; Srivastava, A.K.; Jain, S.; Brahmachari, S.K. CAG repeat instability at SCA2 locus: Anchoring CAA interruptions and linked single nucleotide polymorphisms. Hum. Mol. Genet. 2001, 10, 2437–2446. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, K.; Krzyzosiak, W.J. CAG repeats containing CAA interruptions form branched hairpin structures in spinocerebellar ataxia type 2 transcripts. J. Biol. Chem. 2005, 280, 3898–3910. [Google Scholar] [CrossRef] [PubMed]
- Sobczak, K.; Michlewski, G.; de Mezer, M.; Kierzek, E.; Krol, J.; Olejniczak, M.; Kierzek, R.; Krzyzosiak, W.J. Structural diversity of triplet repeat RNAs. J. Biol. Chem. 2010, 285, 12755–12764. [Google Scholar] [CrossRef] [PubMed]
- Switonski, P.M.; Fiszer, A.; Kazmierska, K.; Kurpisz, M.; Krzyzosiak, W.J.; Figiel, M. Mouse ataxin-3 functional knock-out model. Neuromol. Med. 2011, 13, 54–65. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M. Mfold web server for nucleic acid folding and hybridization prediction. Nucleic Acids Res. 2003, 31, 3406–3415. [Google Scholar] [CrossRef] [PubMed]
- Zuker, M.; Jacobson, A.B. Using reliability information to annotate RNA secondary structures. RNA 1998, 4, 669–679. [Google Scholar] [CrossRef] [PubMed]
- Waugh, A.; Gendron, P.; Altman, R.; Brown, J.W.; Case, D.; Gautheret, D.; Harvey, S.C.; Leontis, N.; Westbrook, J.; Westhof, E.; et al. RNAML: A standard syntax for exchanging RNA information. RNA 2002, 8, 707–717. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Figura, G.; Koscianska, E.; Krzyzosiak, W.J. In Vitro Expansion of CAG, CAA, and Mixed CAG/CAA Repeats. Int. J. Mol. Sci. 2015, 16, 18741-18751. https://doi.org/10.3390/ijms160818741
Figura G, Koscianska E, Krzyzosiak WJ. In Vitro Expansion of CAG, CAA, and Mixed CAG/CAA Repeats. International Journal of Molecular Sciences. 2015; 16(8):18741-18751. https://doi.org/10.3390/ijms160818741
Chicago/Turabian StyleFigura, Grzegorz, Edyta Koscianska, and Wlodzimierz J. Krzyzosiak. 2015. "In Vitro Expansion of CAG, CAA, and Mixed CAG/CAA Repeats" International Journal of Molecular Sciences 16, no. 8: 18741-18751. https://doi.org/10.3390/ijms160818741
APA StyleFigura, G., Koscianska, E., & Krzyzosiak, W. J. (2015). In Vitro Expansion of CAG, CAA, and Mixed CAG/CAA Repeats. International Journal of Molecular Sciences, 16(8), 18741-18751. https://doi.org/10.3390/ijms160818741