
Curcumol Inhibits Growth and Induces Apoptosis of Colorectal Cancer LoVo Cell Line via IGF-1R and p38 MAPK Pathway

Abstract
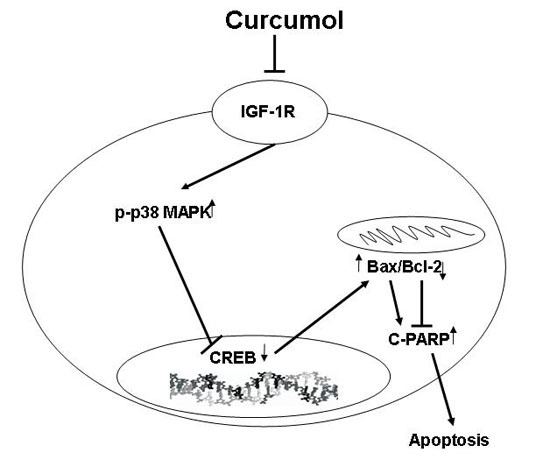
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Result
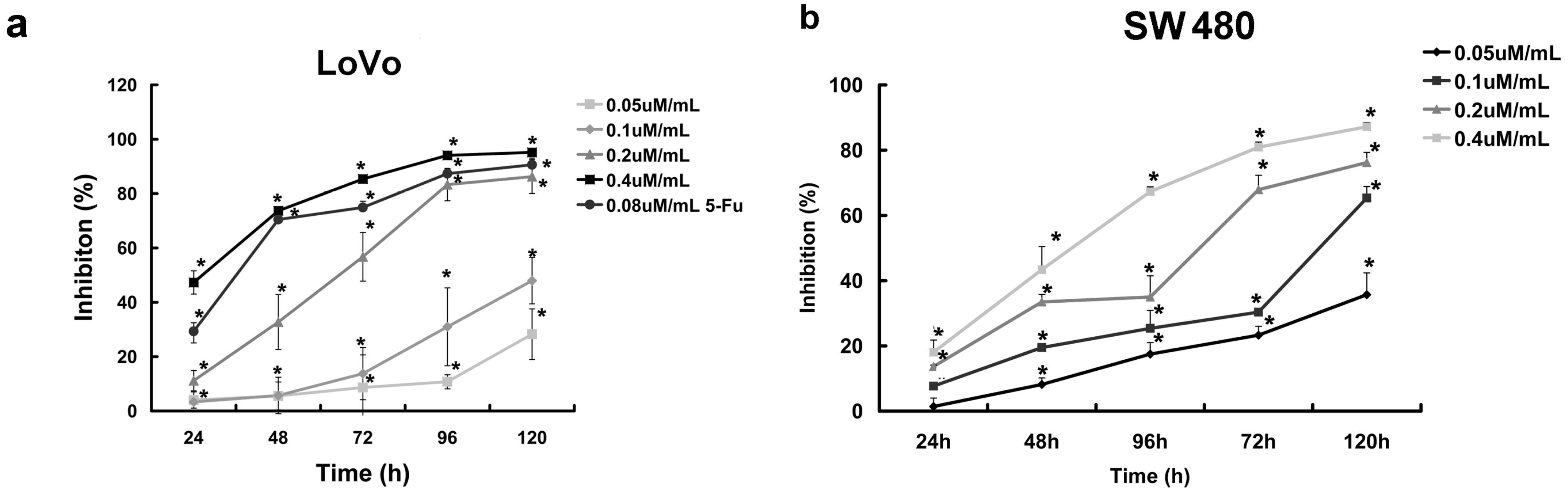
2.1. Curcumol Inhibited Cell Proliferation of Colorectal Cancer

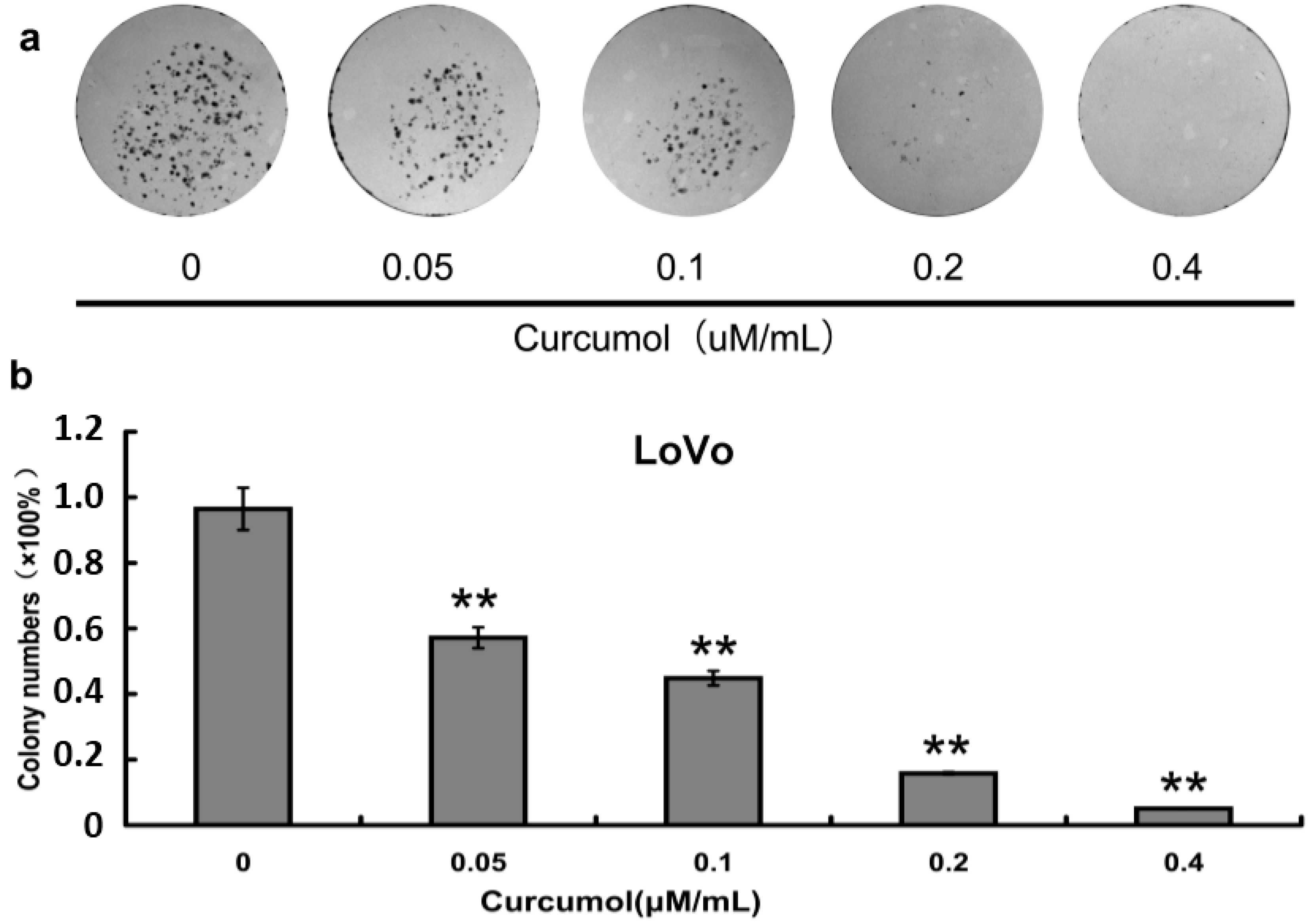
2.2. Inhibition of Cell Survival by Colony-Formation Assay

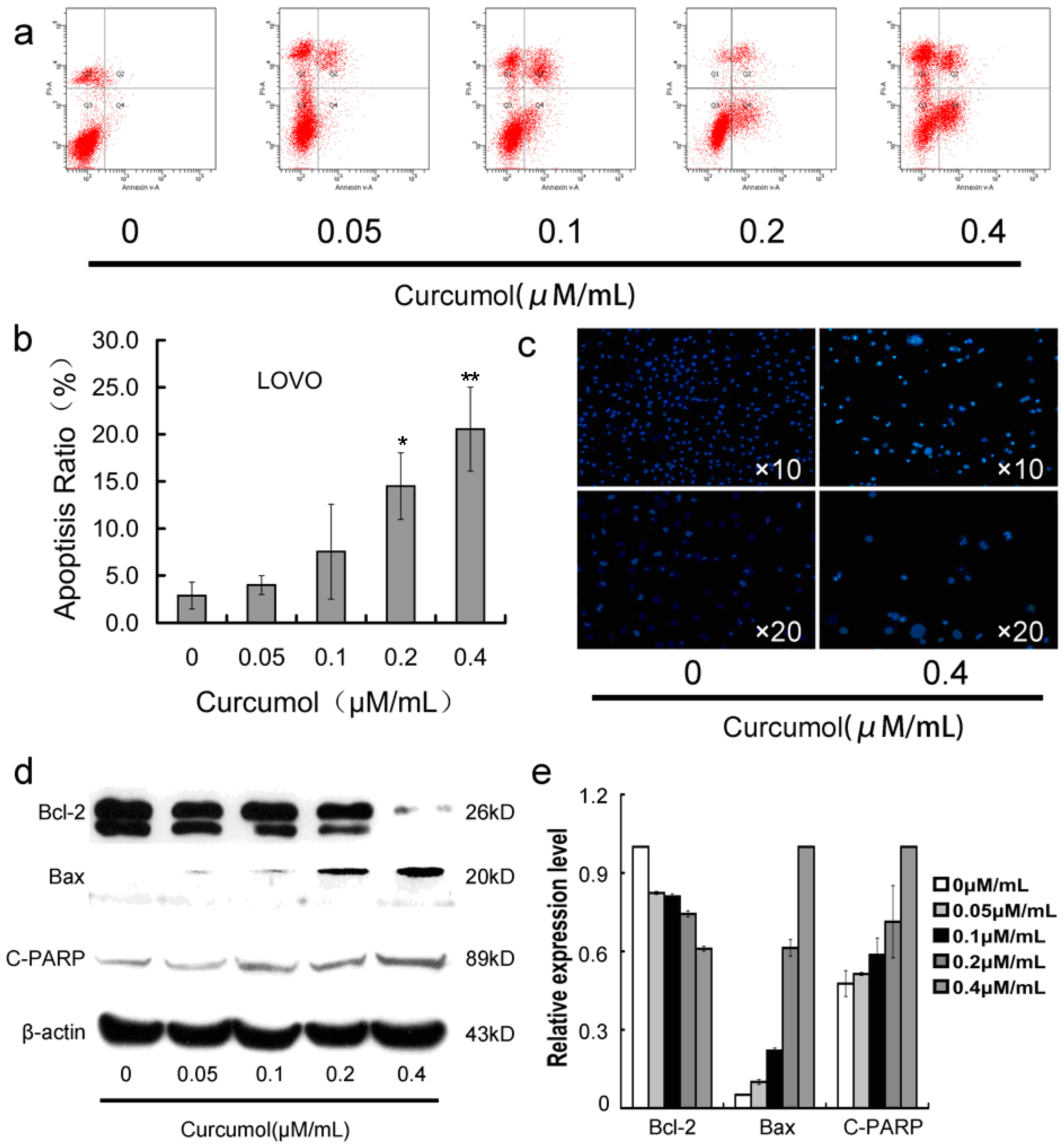
2.3. Curcumol Caused Apoptosis in Colorectal Cancer Cells

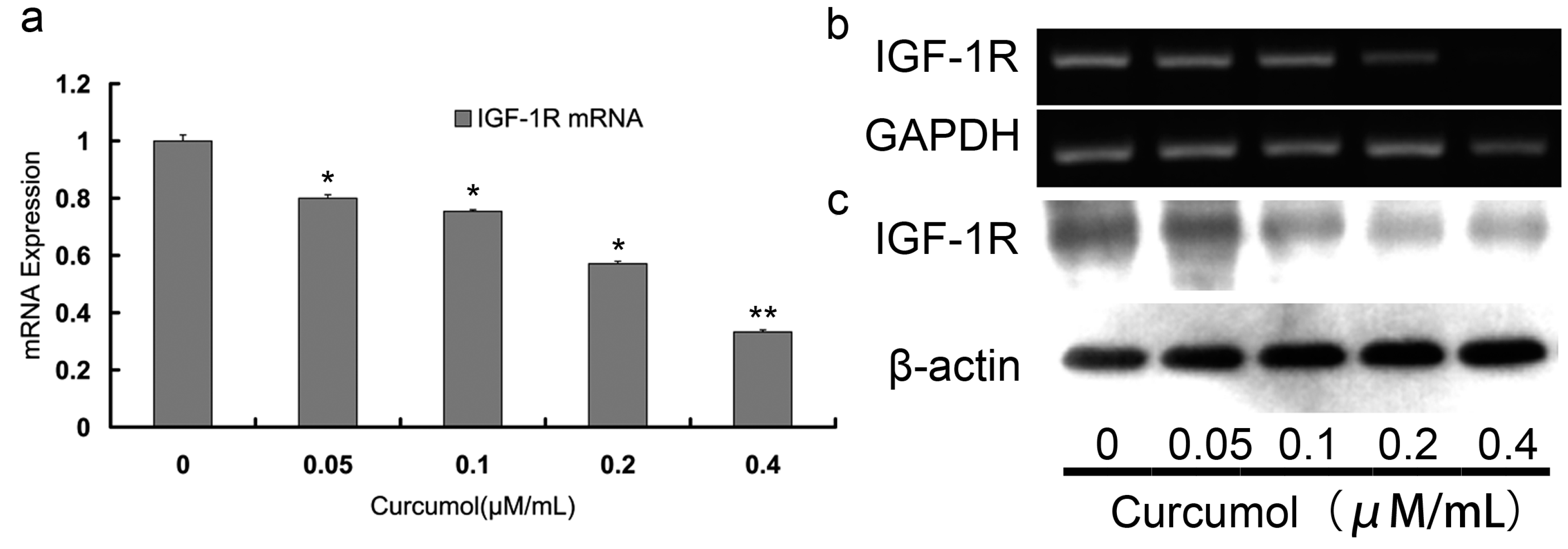
2.4. Curcumol Down-Regulated IGF-1R Levels

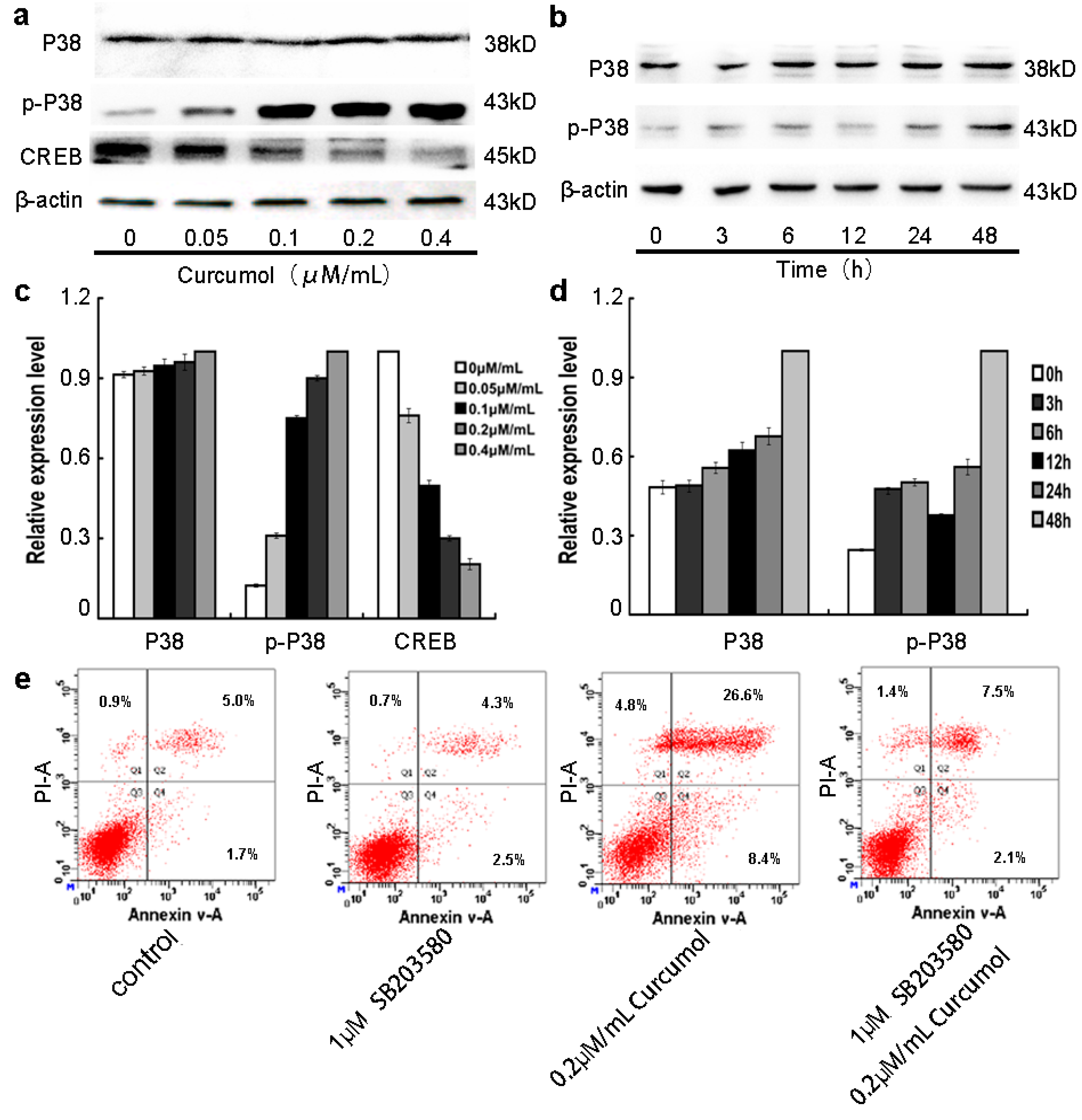
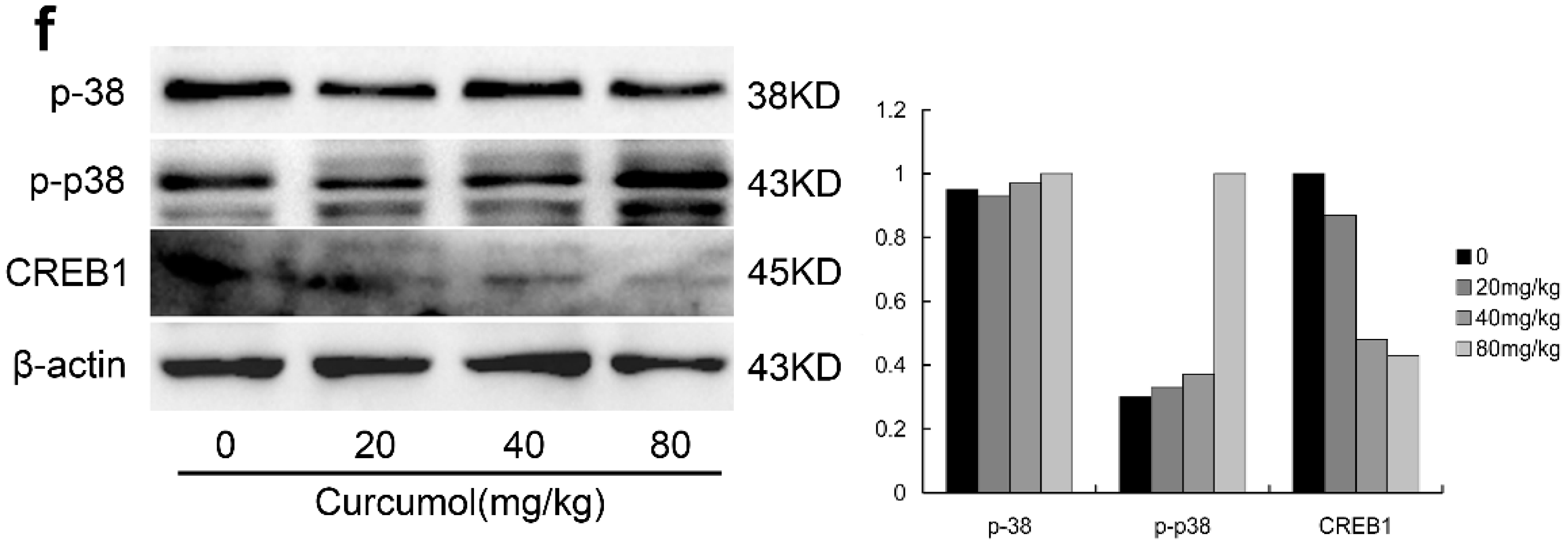
2.5. The p38 MAPK Pathways Were Involved in the Anti-Tumor Effects of Curcumol

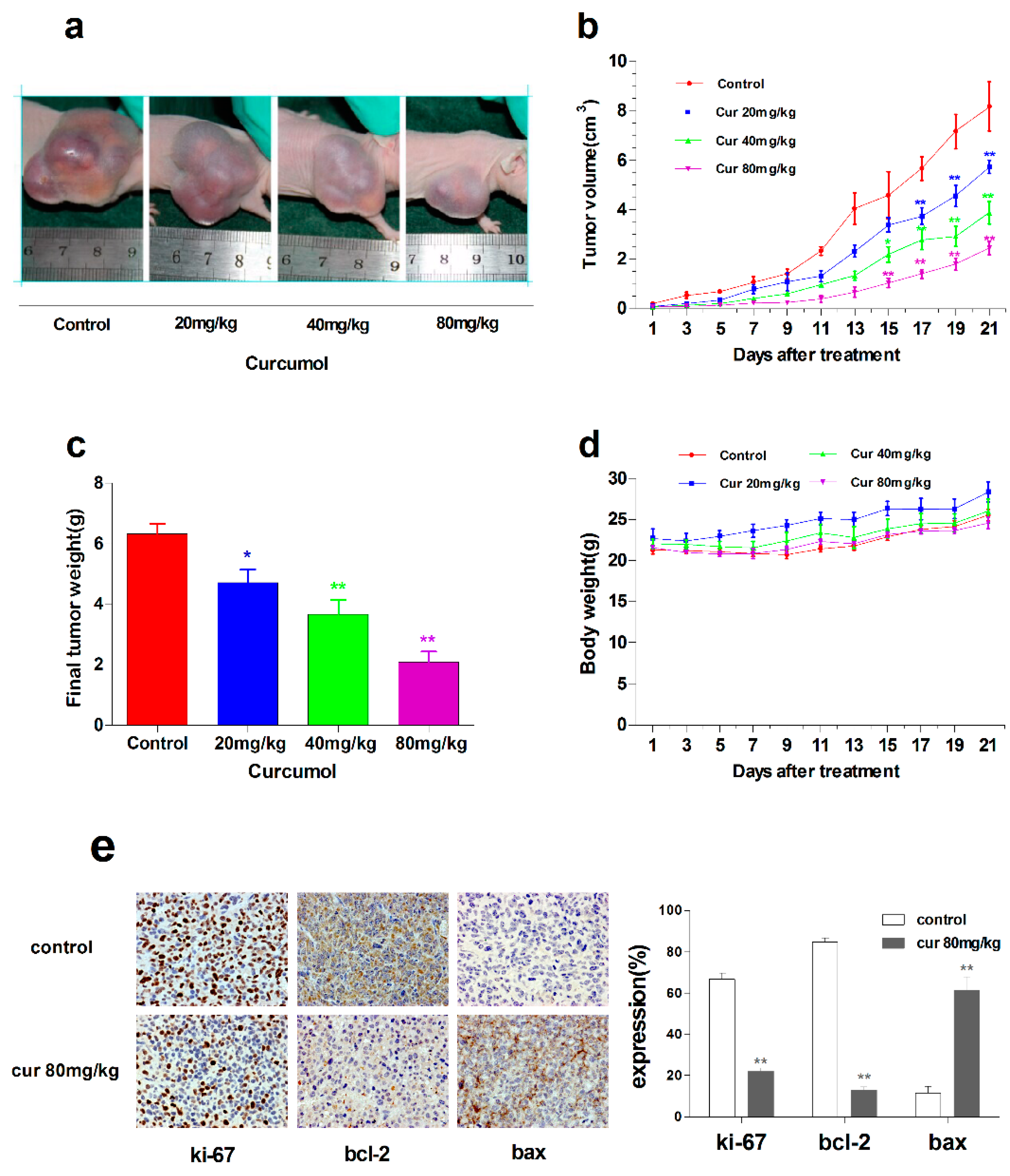
2.6. Curcumol Suppressed Tumor Growth in Vivo


3. Discussion
4. Experimental Section
4.1. Ethics Statement
4.2. Drugs and Reagents
4.3. Cell Culture
4.4. Cell Proliferation Assay
4.5. Colony Formation Assay
4.6. Morphological Analysis after Hoechst Staining
4.7. Apoptosis Analysis by Flow Cytometry (FCM)
4.8. RT-PCR and Real-Time PCR Analysis
4.9. Western Blot Analysis
4.10. Xenograft Tumor Models
4.11. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| CRC | colorectal cancer |
| IGF-1R | insulin-like factor-1 receptor |
| CREB | cAMP-response element binding protein |
| PARP-1 | Poly (ADPribose) polymerase 1 |
| MAPK | mitogen activated protein kinase |
| P13k | phosphatidylinositol 3-kinase. |
References
- Grossi, V.; Peserico, A.; Tezil, T.; Simone, C. p38α MAPK pathway: A key factor in colorectal cancer therapy and chemoresistance. World J. Gastroenterol. 2014, 20, 9744–9758. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Naishadham, D.; Jemal, A. Cancer statistics, 2013. CA Cancer J. Clin. 2013, 63, 11–30. [Google Scholar] [CrossRef] [PubMed]
- Song, X.J.; Xia, Y.; Wang, N.Y.; Zhang, L.D.; Shi, X.H.; Xu, Y.Z.; Ye, T.H.; Shi, Y.J.; Zhu, Y.; Yu, L.T. A Novel benzothiazole derivative YLT322 induces apoptosis via the mitochondrial apoptosis pathway in vitro with anti-tumor activity in solid malignancies. PLoS ONE 2013, 8, e63900. [Google Scholar]
- Tang, W.; Su, G.; Li, J.; Liao, J.; Chen, S.; Huang, C.; Liu, F.; Chen, Q.; Ye, Y. Enhanced anti-colorectal cancer effects of carfilzomib combined with CPT-11 via downregulation of nuclear factor-κB in vitro and in vivo. Int. J. Oncol. 2014, 45, 995–1010. [Google Scholar] [CrossRef] [PubMed]
- Seitz, S.J.; Schleithoff, E.S.; Koch, A.; Schuster, A.; Teufel, A.; Staib, F.; Stremmel, W.; Melino, G.; Krammer, P.H.; Schilling, T.; et al. Chemotherapy-induced apoptosis in hepatocellular carcinoma involves the p53 family and is mediated via the extrinsic and the intrinsic pathway. Int. J. Cancer 2010, 126, 2049–2066. [Google Scholar] [PubMed]
- Dang, Y.Y.; Li, X.C.; Zhang, Q.W.; Li, S.P.; Wang, Y. Preparative isolation and purification of six volatile compounds from essential oil of Curcuma wenyujin using high-performance centrifugal partition chromatography. J. Sep. Sci. 2010, 33, 1658–1664. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, Z.S.; Jiang, F.S.; Deng, X.; Yao, C.S.; Nie, G. Effects of different ingredients of zedoary on gene expression of HSC-T6 cells. World J. Gastroenterol. 2005, 11, 6780–6786. [Google Scholar] [PubMed]
- Xu, L.C.; Bian, K.M.; Liu, Z.M.; Zhou, J.; Wang, G. The inhibitory effect of the curcumol on women cancer cells and synthesis of RNA. Tumor 2005, 25, 570–572. [Google Scholar]
- Zhang, W.W.; Wang, Z.P.; Chen, T. Curcumol induces apoptosis via caspases-independent mitochondrial pathway in human lung adenocarcinoma ASTC-a-1 cells. Med. Oncol. 2011, 28, 307–314. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Wang, Y.; Li, M.; Xu, T.; Wang, X.; Hong, B.; Niu, Y. Curcumol induces HSC-T6 cell death through suppression of Bcl-2: Involvement of PI3K and NF-κB pathways. Eur. J. Pharm. Sci. 2014, 65, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.L.; Guo, J.Q.; Wang, Q.Y.; Lin, H.S.; Yang, Z.P.; Peng, T.; Pan, X.D.; Liu, B.; Wang, S.J.; Zang, L.Q. Curcumol induces apoptosis in SPC-A-1 human lung adenocarcinoma cells and displays anti-neoplastic effects in tumor bearing mice. Asian Pac. J. Cancer Prev. 2015, 16, 2307–2312. [Google Scholar] [PubMed]
- Larsson, O.; Girnita, A.; Girnita, L. Role of insulin-like growth factor 1 receptor signalling in cancer. Br. J. Cancer 2005, 92, 2097–2101. [Google Scholar] [CrossRef] [PubMed]
- Mitsiades, C.S.; Mitsiades, N.S.; McMullan, C.J.; Poulaki, V.; Shringarpure, R.; Akiyama, M.; Hideshima, T.; Chauhan, D.; Joseph, M.; Libermann, T.A.; et al. Inhibition of the insulin-like growth factor receptor-1 tyrosine kinase activity as a therapeutic strategy for multiple myeloma, other hematologic malignancies, and solid tumors. Cancer Cell 2004, 5, 221–230. [Google Scholar] [CrossRef]
- Weber, M.M.; Fottner, C.; Liu, S.B.; Jung, M.C.; Engelhardt, D.; Baretton, G.B. Overexpression of the insulin-like growth factor I receptor in human colon carcinomas. Cancer 2002, 95, 2086–2095. [Google Scholar] [CrossRef] [PubMed]
- Attias, Z.; Werner, H.; Vaisman, N. Folic acid and its metabolites modulate IGF-I receptor gene expression in colon cancer cells in a p53-dependent manner. Endocr. Relat. Cancer 2006, 13, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Rive, S.; Yael, F.; Zohar, A.G.; Ami, F.; Ilan, B.; Haim, W. Metformin downregulates the insulin/IGF-I signaling pathway and inhibits different Uterine Serous Carcinoma (USC) Cells proliferation and migration in p53-dependent or -independent manners. PLoS ONE 2013, 8, e61537. [Google Scholar]
- Chen, J.; Hou, R.; Zhang, X.; Ye, Y.; Wang, Y.; Tian, J. Calycosin suppresses breast cancer cell growth via ERβ-dependent regulation of IGF-1R, p38 MAPK and PI3K/Akt pathways. PLoS ONE 2014, 9, e91245. [Google Scholar] [CrossRef] [PubMed]
- Heller, G.; Zöchbauer-Müller, S. Insulin-like growth factor type I receptor: A new target in the treatment of non-small cell lung cancer patients. memo-Mag. Eur. Med. Oncol. 2010, 3, 81–84. [Google Scholar] [CrossRef]
- Valenciano, A.; Henríquez-Hernández, L.A.; Moreno, M.; Lloret, M.; Lara, P.C. Role of IGF-1 receptor in radiation response. Transl. Oncol. 2012, 5, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Dhillon, A.S.; Hagan, S.; Rath, O.; Kolch, W. MAP kinase signalling pathways in cancer. Oncogene 2007, 26, 3279–3290. [Google Scholar] [CrossRef] [PubMed]
- Wagner, E.F.; Nebreda, A.R. Signal integration by JNK and p38 MAPK pathways in cancer development. Nat. Rev. Cancer 2009, 9, 537–549. [Google Scholar] [CrossRef] [PubMed]
- Hui, L.; Bakiri, L.; Mairhorfer, A.; Schweifer, N.; Haslinger, C.; Kenner, L.; Komnenovic, V.; Scheuch, H.; Beug, H.; Wagner, E.F. p38α suppresses normal and cancer cell proliferation by antagonizing the JNK-c-Jun pathway. Nat. Genet. 2007, 39, 741–749. [Google Scholar] [CrossRef] [PubMed]
- Di Conza, G.; Buttarelli, M.; Monti, O.; Pellegrino, M.; Mancini, F.; Pontecorvi, A.; Scotlandi, K.; Moretti, F. IGF-1R/MDM2 relationship confers enhanced sensitivity to RITA in ewing sarcoma cells. Mol. Cancer Ther. 2012, 11, 1247–1256. [Google Scholar] [CrossRef] [PubMed]
- Gombos, A.; Metzger-Filho, O.; Dal Lago, L.; Awada-Hussein, A. Clinical development of insulin-like growth factor receptor-1 (IGF-1R) inhibitors: At the crossroad? Investig. New Drugs 2012, 30, 2433–2442. [Google Scholar] [CrossRef] [PubMed]
- Hakam, A.; Yeatman, T.J.; Lu, L.; Mora, L.; Marcet, G.; Nicosia, S.V.; Karl, R.C.; Coppola, D. Expression of insulin-like growth factor-1 receptor in human colorectal cancer. Hum. Pathol. 1999, 30, 1128–1133. [Google Scholar] [CrossRef]
- Hui, L.J.; Bakiri, L.; Stepniak, E.; Wagner, E. p38α: A suppressor of cell proliferation and tumorigenesis. Cell Cycle 2007, 6, 2429–2433. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Li, S.; Priceman, S.J.; Xin, H.; Zhang, W.; Deng, J.; Liu, Y.; Huang, J.; Zhu, W.; Chen, M.; et al. Icaritin inhibits JAK/STAT3 signaling and growth of renal cell carcinoma. PLoS ONE 2013, 8, e81657. [Google Scholar]
- Liu, S.Q.; Huang, J.A.; Qin, M.B.; Su, Y.J.; Lai, M.Y.; Jiang, H.X.; Tang, G.D. Sphingosine kinase 1 enhances colon cancer cell proliferation and invasion by upregulating the production of MMP-2/9 and uPA via MAPK pathways. Int. J. Colorectal Dis. 2012, 27, 1569–1578. [Google Scholar] [CrossRef] [PubMed]
- Hui, K.; Yang, Y.; Shi, K.; Luo, H.; Duan, J.; An, J.; Wu, P.; Ci, Y.; Shi, L.; Xu, C. The p38 MAPK-regulated PKD1/CREB/Bcl-2 pathway contributes to selenite-induced colorectal cancer cell apoptosis in vitro and in vivo. Cancer Lett. 2014, 354, 189–199. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Chen, X.; Zeng, J.H. Effect of curcumolon proliferation and apoptosis of nasopharyngeal carcinoma cellline CNE-2. Chin. J. Cell. Mol. Immunol. 2011, 27, 790–792. [Google Scholar]
- Evan, G.I.; Vousden, K. Proliferation, cell cycleand apoptosis in cancer. Nature 2001, 411, 342–348. [Google Scholar] [CrossRef] [PubMed]
- Yin, X.; Zhang, J.; Li, X.; Liu, D.; Feng, C.; Liang, R.; Zhuang, K.; Cai, C.; Xue, X.; Jing, F.; et al. DADS suppresses human esophageal xenograft tumors through RAF/MEK/ERK and mitochondria-dependent pathways. Int. J. Mol. Sci. 2014, 15, 12422–12441. [Google Scholar] [CrossRef] [PubMed]
- Mayeenuddin, L.H.; Yu, Y.; Kang, Z.; Helman, L.J.; Cao, L. Insulin-like growth factor 1 receptor antibody induces rhabdomyosarcoma cell death via a process involving AKT and Bcl-xL. Oncogene 2010, 29, 6367–6377. [Google Scholar] [CrossRef] [PubMed]
- Peiro, G.; Lohse, P.; Mayr, D.; Diebold, J. Insulin-like growth factor-i receptor and PTEN protein expression in endometrial carcinoma: correlation with Bax and Bcl-2 expression, microsatellite instability status, and outcome. Am. J. Clin. Pathol. 2003, 120, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Michael, Höpfner; Andreas P., Sutter; Alexander, Huether; Viola, Baradari; Scherübl, H. Tyrosine kinase of insulin-like growth factor receptor as target for novel treatment and prevention strategies of colorectal cancer. World J. Gastroenterol. 2006, 12, 5635–5643. [Google Scholar]
- Sophie, G.; Robert, B.; Alinel, K.-C.; Sophie, T.; Emmanuel, V.O. The insulin and insulin-like growth factor-I receptor substrate IRS-1 associates with and activates phosphatidylinositol 3-kinase in vitro. J. Biol. Chem. 1993, 268, 7358–7364. [Google Scholar]
- Koul, H.K.; Pal, M.; Koul, S. Role of p38 MAP kinase signal transduction in solid tumors. Genes Cancer 2013, 4, 342–359. [Google Scholar] [CrossRef] [PubMed]
- Porras, A.; Zuluaga, S.; Black, E.; Valladares, A.; Alvarez, A.M.; Ambrosino, C.; Benito, M.; Nebreda, A.R. p38α mitogen-activated protein kinase sensitizes cells to apoptosis induced by different stimuli. Mol. Biol. Cell 2004, 15, 922–933. [Google Scholar] [CrossRef] [PubMed]
- Gupta, J.; del Barco Barrantes, I.; Igea, A.; Sakellariou, S.; Pateras, I.S.; Gorgoulis, V.G.; Nebreda, A.R. Dual function of p38α MAPK in colon cancer: Suppression of colitis-associated tumor initiation but requirement for cancer cell survival. Cancer Cell 2014, 25, 484–500. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Li, H.Y.; Zhang, Z.H.; Bian, H.L.; Lin, G. Garlic-derived compound S-allylmercaptocysteine inhibits cell growth and induces apoptosis via the JNK and p38 pathways in human colorectal carcinoma cells. Oncol. Lett. 2014, 8, 2591–2596. [Google Scholar] [CrossRef] [PubMed]
- Dolado, I.; Swat, A.; Ajenjo, N.; de Vita, G.; Cuadrado, A.; Nebreda, A.R. p38α MAP kinase as a sensor of reactive oxygen species in tumorigenesis. Cancer Cell 2007, 11, 191–205. [Google Scholar] [CrossRef] [PubMed]
- De Zutter, G.S.; Davis, R.J. Pro-apoptotic gene expression mediated by the p38 mitogen-activated protein kinase signal transduction pathway. Proc. Natl. Acad. Sci. USA 2001, 98, 6168–6173. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Wang, K.; Ren, Y.; Zhang, L.; Tang, X.J.; Zhang, H.M.; Zhao, C.Q.; Liu, P.J.; Zhang, J.M.; He, J.J. MAPK signaling mediates sinomenine hydrochloride-induced human breast cancer cell death via both reactive oxygen species-dependent and -independent pathways: An in vitro and in vivo study. Cell Death Dis. 2014, 5. [Google Scholar] [CrossRef] [PubMed]
- Matesic, D.F.; Sidorova, T.S.; Burns, T.J.; Bell, A.M.; Tran, P.L.; Ruch, R.J.; May, S.W. p38 MAPK activation, JNK inhibition, neoplastic growth inhibition, and increased gap junction communication in human lung carcinoma and Ras-transformed cells by 4-phenyl-3-butenoic acid. J. Cell. Biochem. 2012, 113, 269–281. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Wu, N.; Ma, L.N.; Zhong, J.T.; Liu, G.; Zheng, L.H.; Lin, X.K. p38 MAPK signaling mediates mitochondrial apoptosis in cancer cells induced by oleanolic acid. Asian Pac. J. Cancer Prev. 2014, 15, 4519–4525. [Google Scholar] [CrossRef] [PubMed]
- Guo, G.C.; Yang, W.C.; Zhang, Q.Q.; Bo, Y.L. Oleanolic acid suppresses migration and invasion of malignant glioma cells by inactivating MAPK/ERK signaling pathway. PLoS ONE 2013, 8, e72079. [Google Scholar] [CrossRef] [PubMed]
- Hsu, W.H.; Hsieh, Y.S.; Kuo, H.C.; Teng, C.Y.; Huang, H.I.; Wang, C.J.; Yang, S.F.; Liou, Y.S.; Kuo, W.H. Berberine induces apoptosis in SW620 human colonic carcinoma cells through generation of reactive oxygen species and activation of JNK/p38 MAPK and FasL. Arch. Toxicol. 2007, 81, 719–728. [Google Scholar] [CrossRef] [PubMed]
- Chiacchiera, F.; Matrone, A.; Ferrari, E.; Ingravallo, G.; Lo Sasso, G.; Murzilli, S.; Petruzzelli, M.; Salvatore, L.; Moschetta, A.; Simone, C. p38α blockade inhibits colorectal cancer growth in vivo by inducing a switch from HIF1α- to FoxO-dependent transcription. Cell Death Differ. 2009, 16, 1203–11214. [Google Scholar] [CrossRef] [PubMed]
- Vanamala, J.; Reddivari, L.; Radhakrishnan, S.; Tarver, C. Resveratrol suppresses IGF-1 induced human colon cancer cell proliferation and elevates apoptosis via suppression of IGF-1R/Wnt and activation of p53 signaling pathways. BMC Cancer 2010, 10. [Google Scholar] [CrossRef] [PubMed]
- Chen, X.; Wang, J.; Jiang, X.S.; Zeng, J.H.; Huang, F.X. Effects of curcumol on apoptosis induce factor, poly ADP-ribose polymerase and caspase-3 in lung cancer cell line A549. J. Exp. Tradit. Med. Formulae 2011, 19, 157–159. [Google Scholar]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, J.; Huang, F.; Bai, Z.; Chi, B.; Wu, J.; Chen, X. Curcumol Inhibits Growth and Induces Apoptosis of Colorectal Cancer LoVo Cell Line via IGF-1R and p38 MAPK Pathway. Int. J. Mol. Sci. 2015, 16, 19851-19867. https://doi.org/10.3390/ijms160819851
Wang J, Huang F, Bai Z, Chi B, Wu J, Chen X. Curcumol Inhibits Growth and Induces Apoptosis of Colorectal Cancer LoVo Cell Line via IGF-1R and p38 MAPK Pathway. International Journal of Molecular Sciences. 2015; 16(8):19851-19867. https://doi.org/10.3390/ijms160819851
Chicago/Turabian StyleWang, Juan, Fengxiang Huang, Zhun Bai, Bixia Chi, Jiacai Wu, and Xu Chen. 2015. "Curcumol Inhibits Growth and Induces Apoptosis of Colorectal Cancer LoVo Cell Line via IGF-1R and p38 MAPK Pathway" International Journal of Molecular Sciences 16, no. 8: 19851-19867. https://doi.org/10.3390/ijms160819851
APA StyleWang, J., Huang, F., Bai, Z., Chi, B., Wu, J., & Chen, X. (2015). Curcumol Inhibits Growth and Induces Apoptosis of Colorectal Cancer LoVo Cell Line via IGF-1R and p38 MAPK Pathway. International Journal of Molecular Sciences, 16(8), 19851-19867. https://doi.org/10.3390/ijms160819851