
Fibroblast Growth Factor 21 Suppresses Adipogenesis in Pig Intramuscular Fat Cells

Abstract
:1. Introduction
2. Result
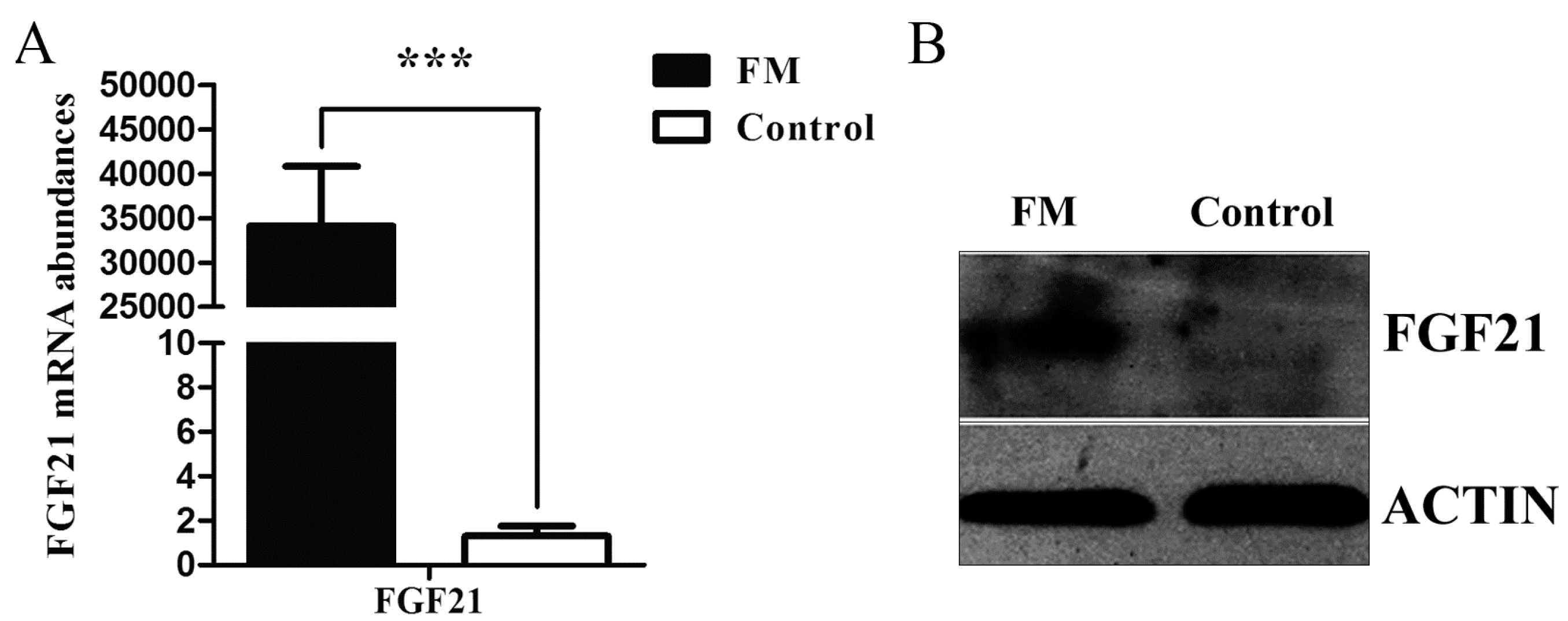
2.1. Low Expression of Fibroblast Growth Factor 21 (FGF21) in Fatty Tissue

2.2. Establishment and Identification of the Intramuscular Preadipocyte Cell Line

2.3. Cells Transfection and Stable FGF21 Cell Line Acquisition

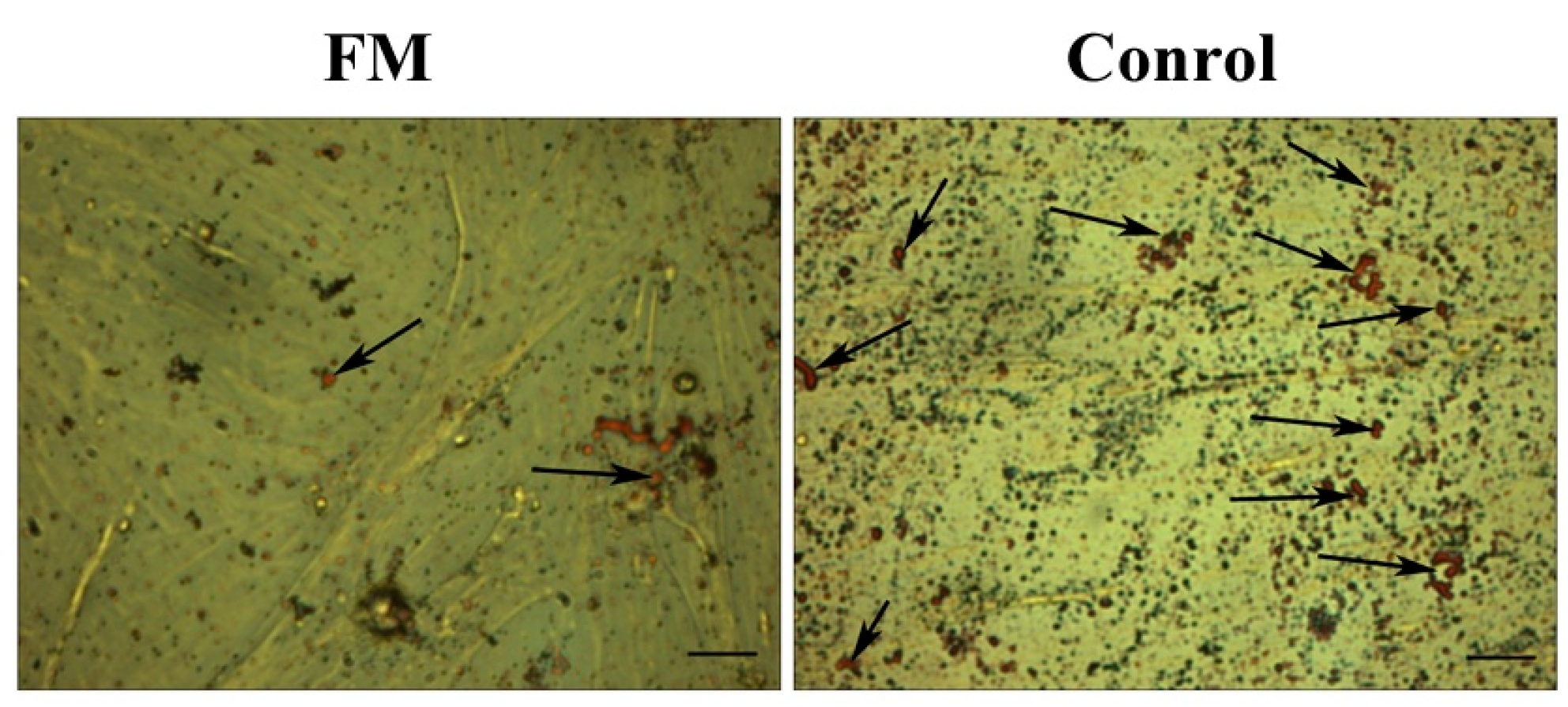
2.4. The Function of FGF21 in Adipogenesis



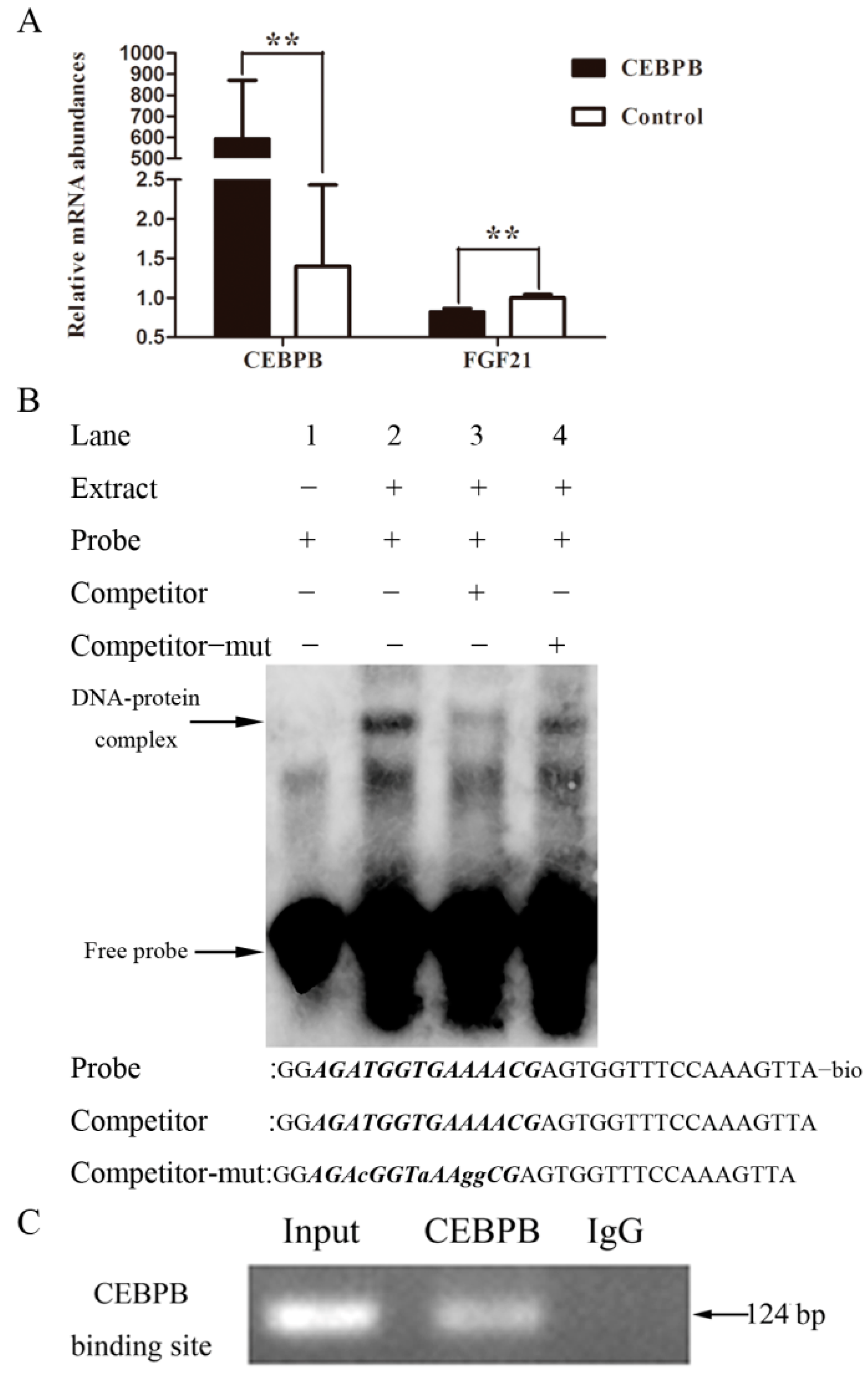
2.5. The Inhibitory Effect of the Transcription Factor CEBPB in FGF21 Expression

3. Discussion

4. Experimental Section
4.1. Ethics Statement
4.2. Expression Profile Analysis of FGF21
4.3. Isolation and Identification of Intramuscular Preadipocyte
4.4. Stable Transfection
4.5. RNA Extraction and qRT-PCR

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Genes | Primer Sequences (5′–3′) | Length of Amplicon (bp) |
|---|---|---|
| FGF21 | F: ACTGTGGGTCCCTGTGCTG R: ATCCGTGTAGAGGTATCGTTGG | 118 |
| PPARG | F: TCCCGCTGACCAAAGCAAAGGC R: CCACGGAGCGAAACTGACACCC | 195 |
| FABP4 | F: GAAAGTCAAGAGCACGATAACC R: CAAGATACATTCCACCACCAAC | 124 |
| FAS | F: TTACACCTCCCTCAACTTCCG R: GGCACCATTCCCATCACG | 153 |
| HSL | F: GTCTTTGCGGGTATTCGG R: GAGTTCGGCCAGGTTGTG | 228 |
| Visfatin | F: TGCCTTTGGTTCTGGTGG R: GCGTAATGAGGTGCTGCTTC | 354 |
| LPL | F: TAACGAACCCGACCAGCATC R: TACACCACCGCCACAGCAA | 140 |
| FGFR1 | F: GAATCGGAGGCTACAAGGTC R: GATAGAGTTACCCGCCAAGC | 305 |
| FGFR2 | F: CTGCCGCCAACTCTGTCA R: CGGATGGAACCACGCTTT | 165 |
| Klotho | F: TCATCCTGTCACCGTTTATTC R: CCTCCACCTGAAATGCTCC | 169 |
| CEBPA | F: CGGTGCGTCTAAGATGAGG R: AGCGGTGAGTTTGCGTTT | 120 |
| CEBPB | F: AGCCTGTCCACATCCTCG R: CACGGTCTTCTTGGTCTTACTC | 135 |
| CEBPD | F: TCAAACACGCCGAACTACAC R: GAGCAAAGGGAAAGCAAATC | 207 |
| GLUT4 | F: TGTTGCGGATGCTATGGG R: GGGTTTCACCTCCTGCTCTAA | 169 |
| ADIPOQ | F: CTGGCGAGAAGAGTGAGA R: TGCTGAACGGTAGACATAGGC | 158 |
| AGPAT2 | F: GTTCGTCCGCTCCTACAA R: CAGGATGCTCTGGTGGTTG | 112 |
| PLIN1 | F: GTGCCAGGAACAGCAACAG R: GGGCTCTACCACCTTCTCATC | 194 |
| FOXO1 | F: AAGACCGCTTTACAAGTGCC R: TCTCCATCCATGAGGTCGTT | 200 |
| LSD1 | F: AGCCAGTTGACAGTGAGGAAT R: CTGAAGCGGTGTAGCGAAC | 109 |
| LRP6 | F: GCAGGGTGGAATGAATGTG R: GAGCAGGAAAGTAGTTGGAGC | 150 |
| β -catenin | F: AAGCAGGTGGATCTATTTCATG R: AGCATTGTATCACAGCAGGTTA | 159 |
| GAPDH | F: GGGCATGAACCATGAGAAGT R: AAGCAGGGATGATGTTCTGG124 | 230 |
| KLF2 | F: GGCGAGAAGCCCTAACACT R: CGCACAGATGGCATTGGA | 121 |
| KLF3 | F: CGCACAGATGGCATTGGA R: CAGCACATTCCTCCCAGTTA | 184 |
| KLF4 | F: CGGAGGGAGACGGAGGAGTT R: TGAAGCCGACGAGGACACG | 108 |
| KLF5 | F: GAAGGGTGCGACTGGAGGTT R: TGTGCTGGGCGAGGTGAT | 133 |
| COUP-TF ii | F: CAAGGCCATAGTCCTGTTCACC R: CGTACTCTTCCAAAGCACACTGG | 101 |
4.6. CEBPA Promoter Methylation Analysis
4.7. Western Blot
4.8. Electrophoretic Mobility Shift Assay (EMSA)
4.9. Chromatin Immunoprecipitation (CHIP) Assay
4.10. Statistical Analysis
5. Conclusions
- FGF21 suppressed adipogenesis by reducing LSD1 expression.
- The transcription factor CEBPB can bind to the promoter of FGF21 directly, and attenuates the expression of FGF21.
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Muoio, D.M. Revisiting the connection between intramyocellular lipids and insulin resistance: A long and winding road. Diabetologia 2012, 55, 2551–2554. [Google Scholar] [CrossRef] [PubMed]
- Guo, Z. Intramyocellular lipids: Maker vs. marker of insulin resistance. Med. Hypotheses 2008, 70, 625–629. [Google Scholar] [CrossRef] [PubMed]
- Cesar, A.S.; Regitano, L.C.; Koltes, J.E.; Fritz-Waters, E.R.; Lanna, D.P.; Gasparin, G.; Mourao, G.B.; Oliveira, P.S.; Reecy, J.M.; Coutinho, L.L. Putative regulatory factors associated with intramuscular fat content. PLoS ONE 2015, 10, e0128350. [Google Scholar] [CrossRef] [PubMed]
- Larson-Meyer, D.E.; Newcomer, B.R.; Ravussin, E.; Volaufova, J.; Bennett, B.; Chalew, S.; Cefalu, W.T.; Sothern, M. Intrahepatic and intramyocellular lipids are determinants of insulin resistance in prepubertal children. Diabetologia 2011, 54, 869–875. [Google Scholar] [CrossRef] [PubMed]
- Christodoulides, C.; Lagathu, C.; Sethi, J.K.; Vidal-Puig, A. Adipogenesis and WNT signalling. Trends Endocrinol. Metab. 2009, 20, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Ross, S.E. Inhibition of Adipogenesis by Wnt Signaling. Science 2000, 289, 950–953. [Google Scholar] [CrossRef] [PubMed]
- Bennett, C.N.; Ross, S.E.; Longo, K.A.; Bajnok, L.; Hemati, N.; Johnson, K.W.; Harrison, S.D.; MacDougald, O.A. Regulation of Wnt signaling during adipogenesis. J. Biol. Chem. 2002, 277, 30998–31004. [Google Scholar] [CrossRef] [PubMed]
- Waki, H.; Park, K.W.; Mitro, N.; Pei, L.; Damoiseaux, R.; Wilpitz, D.C.; Reue, K.; Saez, E.; Tontonoz, P. The small molecule harmine is an antidiabetic cell-type-specific regulator of PPARγ expression. Cell Metab. 2007, 5, 357–370. [Google Scholar] [CrossRef] [PubMed]
- Nakae, J.; Kitamura, T.; Kitamura, Y.; Biggs, W.H., III; Arden, K.C.; Accili, D. The forkhead transcription factor Foxo1 regulates adipocyte differentiation. Dev. Cell 2003, 4, 119–129. [Google Scholar] [CrossRef]
- Liu, W.; Singh, R.; Choi, C.S.; Lee, H.Y.; Keramati, A.R.; Samuel, V.T.; Lifton, R.P.; Shulman, G.I.; Mani, A. Low density lipoprotein (LDL) receptor-related protein 6 (LRP6) regulates body fat and glucose homeostasis by modulating nutrient sensing pathways and mitochondrial energy expenditure. J. Biol. Chem. 2012, 287, 7213–7223. [Google Scholar] [CrossRef] [PubMed]
- Oates, A.C. The zebrafish klf gene family. 2001, 98, 1792–1801. [Google Scholar] [CrossRef] [PubMed]
- De Ruijter, A.J.M.; van Gennip, A.H.; Caron, H.N.; Kemp, S.; van Kuilenburg, A.B.P. Histone deacetylases (HDACs): Characterization of the classical HDAC family. Biochem. J. 2003, 370, 737–749. [Google Scholar] [PubMed]
- Lowe, C.E.; O’Rahilly, S.; Rochford, J.J. Adipogenesis at a glance. J. Cell Sci. 2011, 124, 2681–2686. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Miller, R.A.; Patel, R.T.; Chen, J.; Dhir, R.; Wang, H.; Zhang, D.; Graham, M.J.; Unterman, T.G.; Shulman, G.I.; et al. Hepatic Hdac3 promotes gluconeogenesis by repressing lipid synthesis and sequestration. Nat. Med. 2012, 18, 934–942. [Google Scholar] [CrossRef] [PubMed]
- Inagaki, T.; Dutchak, P.; Zhao, G.; Ding, X.; Gautron, L.; Parameswara, V.; Li, Y.; Goetz, R.; Mohammadi, M.; Esser, V.; et al. Endocrine regulation of the fasting response by PPARα-mediated induction of fibroblast growth factor 21. Cell Metab. 2007, 5, 415–425. [Google Scholar] [CrossRef] [PubMed]
- Arner, P.; Pettersson, A.; Mitchell, P.J.; Dunbar, J.D.; Kharitonenkov, A.; Ryden, M. FGF21 attenuates lipolysis in human adipocytes—A possible link to improved insulin sensitivity. FEBS Lett. 2008, 582, 1725–1730. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ge, H.; Weiszmann, J.; Hecht, R.; Li, Y.S.; Veniant, M.M.; Xu, J.; Wu, X.; Lindberg, R.; Li, Y. Inhibition of lipolysis may contribute to the acute regulation of plasma FFA and glucose by FGF21 in ob/ob mice. FEBS Lett. 2009, 583, 3230–3234. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.; Hoo, R.L.; Konishi, M.; Itoh, N.; Lee, P.C.; Ye, H.Y.; Lam, K.S.; Xu, A. Growth hormone induces hepatic production of fibroblast growth factor 21 through a mechanism dependent on lipolysis in adipocytes. J. Biol. Chem. 2011, 286, 34559–34566. [Google Scholar] [CrossRef] [PubMed]
- Murata, Y.; Konishi, M.; Itoh, N. FGF21 as an endocrine regulator in lipid metabolism: From molecular evolution to physiology and pathophysiology. J. Nutr. Metab. 2011, 2011. [Google Scholar] [CrossRef] [PubMed]
- Canto, C.; Auwerx, J. Cell biology. FGF21 takes a fat bite. Science 2012, 336, 675–676. [Google Scholar] [CrossRef] [PubMed]
- Coskun, T.; Bina, H.A.; Schneider, M.A.; Dunbar, J.D.; Hu, C.C.; Chen, Y.; Moller, D.E.; Kharitonenkov, A. Fibroblast growth factor 21 corrects obesity in mice. Endocrinology 2008, 149, 6018–6027. [Google Scholar] [CrossRef] [PubMed]
- Chau, M.D.; Gao, J.; Yang, Q.; Wu, Z.; Gromada, J. Fibroblast growth factor 21 regulates energy metabolism by activating the AMPK-SIRT1-PGC-1α pathway. Proc. Natl. Acad. Sci. USA 2010, 107, 12553–12558. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Lloyd, D.J.; Hale, C.; Stanislaus, S.; Chen, M.; Sivits, G.; Vonderfecht, S.; Hecht, R.; Li, Y.S.; Lindberg, R.A.; et al. Fibroblast growth factor 21 reverses hepatic steatosis, increases energy expenditure, and improves insulin sensitivity in diet-induced obese mice. Diabetes 2009, 58, 250–259. [Google Scholar] [CrossRef] [PubMed]
- Hotta, Y.; Nakamura, H.; Konishi, M.; Murata, Y.; Takagi, H.; Matsumura, S.; Inoue, K.; Fushiki, T.; Itoh, N. Fibroblast growth factor 21 regulates lipolysis in white adipose tissue but is not required for ketogenesis and triglyceride clearance in liver. Endocrinology 2009, 150, 4625–4633. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Shiyanova, T.L.; Koester, A.; Ford, A.M.; Micanovic, R.; Galbreath, E.J.; Sandusky, G.E.; Hammond, L.J.; Moyers, J.S.; Owens, R.A.; et al. FGF-21 as a novel metabolic regulator. J. Clin. Investig. 2005, 115, 1627–1635. [Google Scholar] [CrossRef] [PubMed]
- Huang, Z.; Wang, H.; Lu, M.; Sun, C.; Wu, X.; Tan, Y.; Ye, C.; Zhu, G.; Wang, X.; Cai, L.; et al. A better anti-diabetic recombinant human fibroblast growth factor 21 (rhFGF21) modified with polyethylene glycol. PLoS ONE 2011, 6, e20669. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Wroblewski, V.J.; Koester, A.; Chen, Y.F.; Clutinger, C.K.; Tigno, X.T.; Hansen, B.C.; Shanafelt, A.B.; Etgen, G.J. The metabolic state of diabetic monkeys is regulated by fibroblast growth factor-21. Endocrinology 2007, 148, 774–781. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M. Intramuscular triacylglycerol and insulin resistance: Guilty as charged or wrongly accused? Biochim. Biophys. Acta 2010, 1801, 281–288. [Google Scholar] [CrossRef] [PubMed]
- Muoio, D.M.; Newgard, C.B. Molecular and metabolic mechanisms of insulin resistance and β-cell failure in type 2 diabetes. Nat. Rev. Mol. Cell Biol. 2008, 9, 193–205. [Google Scholar] [CrossRef] [PubMed]
- McGarry, J. Dysregulation of fatty acid metabolism in the etiology of type 2 diabetes. Diabetes 2002, 51, 17–18. [Google Scholar] [CrossRef]
- Siersbaek, R.; Nielsen, R.; Mandrup, S. PPARγ in adipocyte differentiation and metabolism—Novel insights from genome-wide studies. FEBS Lett. 2010, 584, 3242–3249. [Google Scholar] [CrossRef] [PubMed]
- Lehrke, M.; Lazar, M.A. The many faces of PPARγ. Cell 2005, 123, 993–999. [Google Scholar] [CrossRef] [PubMed]
- Picard, F.; Kurtev, M.; Chung, N.; Topark-Ngarm, A.; Senawong, T.; de Machado Oliveira, R.; Leid, M.; McBurney, M.W.; Guarente, L. Sirt1 promotes fat mobilization in white adipocytes by repressing PPAR-(gamma). Nature 2004, 429, 771–776. [Google Scholar] [CrossRef] [PubMed]
- Jing, E.; Gesta, S.; Kahn, C.R. SIRT2 Regulates Adipocyte differentiation through FoxO1 acetylation/deacetylation. Cell Metabol. 2007, 6, 105–114. [Google Scholar] [CrossRef]
- Okamura, M.; Kudo, H.; Wakabayashi, K.; Tanaka, T.; Nonaka, A.; Uchida, A.; Tsutsumi, S.; Sakakibara, I.; Naito, M.; Osborne, T.F.; et al. COUP-TFII acts downstream of Wnt/β-catenin signal to silence PPARγ gene expression and repress adipogenesis. Proc. Natl. Acad. Sci. USA 2009, 106, 5819–5824. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.S.; Feinberg, M.W.; Watanabe, M.; Gray, S.; Haspel, R.L.; Denkinger, D.J.; Kawahara, R.; Hauner, H.; Jain, M.K. The Kruppel-like factor KLF2 inhibits peroxisome proliferator-activated receptor-gamma expression and adipogenesis. J. Biol. Chem. 2003, 278, 2581–2584. [Google Scholar] [CrossRef] [PubMed]
- Sue, N.; Jack, B.H.; Eaton, S.A.; Pearson, R.C.; Funnell, A.P.; Turner, J.; Czolij, R.; Denyer, G.; Bao, S.; Molero-Navajas, J.C.; et al. Targeted disruption of the basic Kruppel-like factor gene (Klf3) reveals a role in adipogenesis. Mol. Cell. Biol. 2008, 28, 3967–3978. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Birsoy, K.; Chen, Z.; Friedman, J. Transcriptional regulation of adipogenesis by KLF4. Cell Metab. 2008, 7, 339–347. [Google Scholar] [CrossRef] [PubMed]
- Kinoshita, M.; Ono, K.; Horie, T.; Nagao, K.; Nishi, H.; Kuwabara, Y.; Takanabe-Mori, R.; Hasegawa, K.; Kita, T.; Kimura, T. Regulation of adipocyte differentiation by activation of serotonin (5-HT) receptors 5-HT2AR and 5-HT2CR and involvement of microRNA-448-mediated repression of KLF5. Mol. Endocrinol. 2010, 24, 1978–1987. [Google Scholar] [CrossRef] [PubMed]
- Musri, M.M.; Carmona, M.C.; Hanzu, F.A.; Kaliman, P.; Gomis, R.; Parrizas, M. Histone demethylase LSD1 regulates adipogenesis. J. Biol. Chem. 2010, 285, 30034–30041. [Google Scholar] [CrossRef] [PubMed]
- Esteve, E.; Ricart, W.; Fernandez-Real, J.M. Adipocytokines and Insulin Resistance: The possible role of lipocalin-2, retinol binding protein-4, and adiponectin. Diabetes Care 2009, 32, S362–S367. [Google Scholar] [CrossRef] [PubMed]
- Gerbens, F.; Jansen, A.; van Erp, A.J.M.; Harders, F.; Meuwissen, T.H.E.; Rettenberger, G.; Veerkamp, J.H.; te Pas, M.F.W. The adipocyte fatty acid-binding protein locus: Characterization and association with intramuscular fat content in pigs. Mamm. Genome 1998, 9, 1022–1026. [Google Scholar] [CrossRef]
- Jenkins, N.T.; McKenzie, J.A.; Damcott, C.M.; Witkowski, S.; Hagberg, J.M. Endurance exercise training effects on body fatness, VO2max, HDL-C subfractions, and glucose tolerance are influenced by a PLIN haplotype in older Caucasians. J. Appl. Physiol. 2010, 108, 498–506. [Google Scholar] [CrossRef] [PubMed]
- Chen, Y.; Mu, P.; He, S.; Tang, X.; Guo, X.; Li, H.; Xu, H.; Woo, S.L.; Qian, X.; Zeng, L.; et al. Gly482Ser mutation impairs the effects of peroxisome proliferator—Activated receptor γ coactivator-1α on decreasing fat deposition and stimulating phosphoenolpyruvate carboxykinase expression in hepatocytes. Nutr. Res. 2013, 33, 332–339. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, J.A.; Ben Ali, Y.; Abdelkafi, S.; Mendoza, L.D.; Leclaire, J.; Fotiadu, F.; Buono, G.; Carrière, F.; Abousalham, A. In vitro stereoselective hydrolysis of diacylglycerols by hormone-sensitive lipase. Biochim. Biophys. Acta (BBA) Mol. Cell Biol. Lipids 2010, 1801, 77–83. [Google Scholar] [CrossRef] [PubMed]
- Yan, X.; Chen, J.; Zhang, C.; Zhou, S.; Zhang, Z.; Chen, J.; Feng, W.; Li, X.; Tan, Y. FGF21 deletion exacerbates diabetic cardiomyopathy by aggravating cardiac lipid accumulation. J. Cell. Mol. Med. 2015, 19, 1557–1568. [Google Scholar] [CrossRef] [PubMed]
- Cortés, V.A.; Cautivo, K.M.; Rong, S.; Garg, A.; Horton, J.D.; Agarwal, A.K. Leptin ameliorates insulin resistance and hepatic steatosis in Agpat2−/− lipodystrophic mice independent of hepatocyte leptin receptors. J. Lipid Res. 2014, 55, 276–288. [Google Scholar] [CrossRef] [PubMed]
- Kharitonenkov, A.; Larsen, P. FGF21 reloaded: Challenges of a rapidly growing field. Trends Endocrinol. Metabol. 2011, 22, 81–86. [Google Scholar] [CrossRef] [PubMed]
- Jin, D.; Tan, H.J.; Lei, T.; Chen, X.D.; Long, Q.Q.; Feng, B.; Yang, Z.Q. Molecular cloning and characterization of porcine sirtuin genes. Comp. Biochem. Phys. B 2009, 153, 348–358. [Google Scholar] [CrossRef] [PubMed]
- Hou, L.; Ma, F.H.; Yang, J.Z.; Riaz, H.; Wang, Y.L.; Wu, W.J.; Xia, X.L.; Ma, Z.Y.; Zhou, Y.; Zhang, L.; et al. Effects of histone deacetylase inhibitor oxamflatinon in vitro porcine somatic cell nuclear transfer embryos. Cell. Reprogram. 2014, 16, 253–265. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons by Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Y.; Liu, X.; Hou, L.; Wu, W.; Zhao, S.; Xiong, Y. Fibroblast Growth Factor 21 Suppresses Adipogenesis in Pig Intramuscular Fat Cells. Int. J. Mol. Sci. 2016, 17, 11. https://doi.org/10.3390/ijms17010011
Wang Y, Liu X, Hou L, Wu W, Zhao S, Xiong Y. Fibroblast Growth Factor 21 Suppresses Adipogenesis in Pig Intramuscular Fat Cells. International Journal of Molecular Sciences. 2016; 17(1):11. https://doi.org/10.3390/ijms17010011
Chicago/Turabian StyleWang, Yongliang, Xinyi Liu, Liming Hou, Wangjun Wu, Shuhong Zhao, and Yuanzhu Xiong. 2016. "Fibroblast Growth Factor 21 Suppresses Adipogenesis in Pig Intramuscular Fat Cells" International Journal of Molecular Sciences 17, no. 1: 11. https://doi.org/10.3390/ijms17010011