
Transcriptome of Cultured Lung Fibroblasts in Idiopathic Pulmonary Fibrosis: Meta-Analysis of Publically Available Microarray Datasets Reveals Repression of Inflammation and Immunity Pathways

 ,
,
Abstract
:1. Introduction
2. Results
3. Discussion
4. Materials and Methods
4.1. Data Compilation
4.2. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- King, T.E.; Bradford, W.Z.; Castro-Bernardini, S.; Fagan, E.A.; Glaspole, I.; Glassberg, M.K.; Gorina, E.; Hopkins, P.M.; Kardatzke, D.; Lancaster, L.; et al. A phase 3 trial of pirfenidone in patients with idiopathic pulmonary fibrosis. N. Engl. J. Med. 2014, 370, 2083–2092. [Google Scholar] [CrossRef] [PubMed]
- Richeldi, L.; du Bois, R.M.; Raghu, G.; Azuma, A.; Brown, K.K.; Costabel, U.; Cottin, V.; Flaherty, K.R.; Hansell, D.M.; Inoue, Y.; et al. Efficacy and safety of nintedanib in idiopathic pulmonary fibrosis. N. Engl. J. Med. 2014, 370, 2071–2082. [Google Scholar] [CrossRef] [PubMed]
- Crestani, B.; Besnard, V.; Plantier, L.; Borensztajn, K.; Mailleux, A. Fibroblasts: The missing link between fibrotic lung diseases of different etiologies? Respir. Res. 2013, 14. [Google Scholar] [CrossRef] [PubMed]
- Pierce, E.M.; Carpenter, K.; Jakubzick, C.; Kunkel, S.L.; Flaherty, K.R.; Martinez, F.J.; Hogaboam, C.M. Therapeutic targeting of CC ligand 21 or CC chemokine receptor 7 abrogates pulmonary fibrosis induced by the adoptive transfer of human pulmonary fibroblasts to immunodeficient mice. Am. J. Pathol. 2007, 170, 1152–1164. [Google Scholar] [CrossRef] [PubMed]
- Renzoni, E.A.; Abraham, D.J.; Howat, S.; Shi-Wen, X.; Sestini, P.; Bou-Gharios, G.; Wells, A.U.; Veeraraghavan, S.; Nicholson, A.G.; Denton, C.P.; et al. Gene expression profiling reveals novel TGFβ targets in adult lung fibroblasts. Respir. Res. 2004, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lindahl, G.E.; Stock, C.J.; Shi-Wen, X.; Leoni, P.; Sestini, P.; Howat, S.L.; Bou-Gharios, G.; Nicholson, A.G.; Denton, C.P.; Grutters, J.C.; et al. Microarray profiling reveals suppressed interferon stimulated gene program in fibroblasts from scleroderma-associated interstitial lung disease. Respir. Res. 2013, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Peng, R.; Sridhar, S.; Tyagi, G.; Phillips, J.E.; Garrido, R.; Harris, P.; Burns, L.; Renteria, L.; Woods, J.; Chen, L.; et al. Bleomycin induces molecular changes directly relevant to idiopathic pulmonary fibrosis: A model for “active” disease. PLoS ONE 2013, 8, e59348. [Google Scholar] [CrossRef] [PubMed]
- Vuga, L.J.; Ben-Yehudah, A.; Kovkarova-Naumovski, E.; Oriss, T.; Gibson, K.F.; Feghali-Bostwick, C.; Kaminski, N. WNT5A is a regulator of fibroblast proliferation and resistance to apoptosis. Am. J. Respir. Cell Mol. Biol. 2009, 41, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Melboucy-Belkhir, S.; Pradère, P.; Tadbiri, S.; Habib, S.; Bacrot, A.; Brayer, S.; Mari, B.; Besnard, V.; Mailleux, A.; Guenther, A.; et al. Forkhead Box F1 represses cell growth and inhibits COL1 and ARPC2 expression in lung fibroblasts in vitro. Am. J. Physiol. Lung Cell. Mol. Physiol. 2014, 307, L838–L847. [Google Scholar] [CrossRef] [PubMed]
- Gene Expression Omnibus. Available online: http://www.ncbi.nlm.nih.gov/gds/ (accessed 20 November 2016).
- Liang, X.; Sun, Y.; Ye, M.; Scimia, M.C.; Cheng, H.; Martin, J.; Wang, G.; Rearden, A.; Wu, C.; Peterson, K.L.; et al. Targeted ablation of PINCH1 and PINCH2 from murine myocardium results in dilated cardiomyopathy and early postnatal lethality. Circulation 2009, 120, 568–576. [Google Scholar] [CrossRef] [PubMed]
- Pan, D.; Zhe, X.; Jakkaraju, S.; Taylor, G.A.; Schuger, L. P311 induces a TGF-β1-independent, nonfibrogenic myofibroblast phenotype. J. Clin. Investig. 2002, 110, 1349–1358. [Google Scholar] [CrossRef] [PubMed]
- Ihn, H. The role of TGF-β signaling in the pathogenesis of fibrosis in scleroderma. Arch. Immunol. Ther. Exp. 2002, 50, 325–331. [Google Scholar]
- Liu, W.; Ding, I.; Chen, K.; Olschowka, J.; Xu, J.; Hu, D.; Morrow, G.R.; Okunieff, P. Interleukin 1β (IL1B) signaling is a critical component of radiation-induced skin fibrosis. Radiat. Res. 2006, 165, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Christmann, R.B.; Sampaio-Barros, P.; Stifano, G.; Borges, C.L.; de Carvalho, C.R.; Kairalla, R.; Parra, E.R.; Spira, A.; Simms, R.; Capellozzi, V.L.; et al. Association of Interferon- and transforming growth factor β-regulated genes and macrophage activation with systemic sclerosis-related progressive lung fibrosis. Arthritis Rheumatol. 2014, 66, 714–725. [Google Scholar] [CrossRef] [PubMed]
- Park, J.H.; Chung, H.Y.; Kim, M.; Lee, J.H.; Jung, M.; Ha, H. Daumone fed late in life improves survival and reduces hepatic inflammation and fibrosis in mice. Aging Cell 2014, 13, 709–718. [Google Scholar] [CrossRef] [PubMed]
- Attia, M.; Rachez, C.; de Pauw, A.; Avner, P.; Rogner, U.C. Nap1l2 promotes histone acetylation activity during neuronal differentiation. Mol. Cell. Biol. 2007, 27, 6093–6102. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Chen, K.; Guo, L.; Wu, C. Characterization of PINCH-2, a new focal adhesion protein that regulates the PINCH-1-ILK interaction, cell spreading, and migration. J. Biol. Chem. 2002, 277, 38328–38338. [Google Scholar] [CrossRef] [PubMed]
- Tan, J.; Peng, X.; Luo, G.; Ma, B.; Cao, C.; He, W.; Yuan, S.; Li, S.; Wilkins, J.A.; Wu, J. Investigating the role of P311 in the hypertrophic scar. PLoS ONE 2010, 5, e9995. [Google Scholar] [CrossRef] [PubMed]
- Ponticos, M.; Holmes, A.M.; Shi-Wen, X.; Leoni, P.; Khan, K.; Rajkumar, V.S.; Hoyles, R.K.; Bou-Gharios, G.; Black, C.M.; Denton, C.P.; et al. Pivotal role of connective tissue growth factor in lung fibrosis: MAPK-dependent transcriptional activation of type I collagen. Arthritis Rheum. 2009, 60, 2142–2155. [Google Scholar] [CrossRef] [PubMed]
- Sonnylal, S.; Shi-Wen, X.; Leoni, P.; Naff, K.; van Pelt, C.S.; Nakamura, H.; Leask, A.; Abraham, D.; Bou-Gharios, G.; de Crombrugghe, B. Selective expression of connective tissue growth factor in fibroblasts in vivo promotes systemic tissue fibrosis. Arthritis Rheum. 2010, 62, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Allen, J.T.; Knight, R.A.; Bloor, C.A.; Spiteri, M.A. Enhanced insulin-like growth factor binding protein-related protein 2 (Connective tissue growth factor) expression in patients with idiopathic pulmonary fibrosis and pulmonary sarcoidosis. Am. J. Respir. Cell Mol. Biol. 1999, 21, 693–700. [Google Scholar] [CrossRef] [PubMed]
- Raghu, G.; Scholand, M.B.; de Andrade, J.; Lancaster, L.; Mageto, Y.; Goldin, J.; Brown, K.K.; Flaherty, K.R.; Wencel, M.; Wanger, J.; et al. FG-3019 anti-connective tissue growth factor monoclonal antibody: Results of an open-label clinical trial in idiopathic pulmonary fibrosis. Eur. Respir. J. 2016, 47, 1481–1491. [Google Scholar] [CrossRef] [PubMed]
- Bébien, M.; Salinas, S.; Becamel, C.; Richard, V.; Linares, L.; Hipskind, R.A. Immediate-early gene induction by the stresses anisomycin and arsenite in human osteosarcoma cells involves MAPK cascade signaling to Elk-1, CREB and SRF. Oncogene 2003, 22, 1836–1847. [Google Scholar] [CrossRef] [PubMed]
- Sharili, A.S.; Connelly, J.T. Nucleocytoplasmic shuttling: A common theme in mechanotransduction. Biochem. Soc. Trans. 2014, 42, 645–649. [Google Scholar] [CrossRef] [PubMed]
- Luchsinger, L.L.; Patenaude, C.A.; Smith, B.D.; Layne, M.D. Myocardin-related transcription factor-A complexes activate type I collagen expression in lung fibroblasts. J. Biol. Chem. 2011, 286, 44116–44125. [Google Scholar] [CrossRef] [PubMed]
- Sisson, T.H.; Ajayi, I.O.; Subbotina, N.; Dodi, A.E.; Rodansky, E.S.; Chibucos, L.N.; Kim, K.K.; Keshamouni, V.G.; White, E.S.; Zhou, Y.; et al. Inhibition of myocardin-related transcription factor/serum response factor signaling decreases lung fibrosis and promotes mesenchymal cell apoptosis. Am. J. Pathol. 2015, 185, 969–986. [Google Scholar] [CrossRef] [PubMed]
- Haak, A.J.; Tsou, P.-S.; Amin, M.A.; Ruth, J.H.; Campbell, P.; Fox, D.A.; Khanna, D.; Larsen, S.D.; Neubig, R.R. Targeting the myofibroblast genetic switch: Inhibitors of myocardin-related transcription factor/serum response factor-regulated gene transcription prevent fibrosis in a murine model of skin injury. J. Pharmacol. Exp. Ther. 2014, 349, 480–486. [Google Scholar] [CrossRef] [PubMed]
- Chapman, H.A. Disorders of lung matrix remodeling. J. Clin. Investig. 2004, 113, 148–157. [Google Scholar] [CrossRef] [PubMed]
- Elias, J.A.; Jimenez, S.A.; Freundlich, B. Recombinant γ, α, and β interferon regulation of human lung fibroblast proliferation. Am. Rev. Respir. Dis. 1987, 135, 62–65. [Google Scholar] [PubMed]
- Shoda, H.; Yokoyama, A.; Nishino, R.; Nakashima, T.; Ishikawa, N.; Haruta, Y.; Hattori, N.; Naka, T.; Kohno, N. Overproduction of collagen and diminished SOCS1 expression are causally linked in fibroblasts from idiopathic pulmonary fibrosis. Biochem. Biophys. Res. Commun. 2007, 353, 1004–1010. [Google Scholar] [CrossRef] [PubMed]
- Elias, J.A.; Freundlich, B.; Kern, J.A.; Rosenbloom, J. Cytokine networks in the regulation of inflammation and fibrosis in the lung. Chest 1990, 97, 1439–1445. [Google Scholar] [CrossRef] [PubMed]
- Eickelberg, O.; Pansky, A.; Koehler, E.; Bihl, M.; Tamm, M.; Hildebrand, P.; Perruchoud, A.P.; Kashgarian, M.; Roth, M. Molecular mechanisms of TGF-(β) antagonism by interferon (γ) and cyclosporine A in lung fibroblasts. FASEB J. 2001, 15, 797–806. [Google Scholar] [CrossRef] [PubMed]
- O’Dwyer, D.N.; Armstrong, M.E.; Trujillo, G.; Cooke, G.; Keane, M.P.; Fallon, P.G.; Simpson, A.J.; Millar, A.B.; McGrath, E.E.; Whyte, M.K.; et al. The Toll-like receptor 3 L412F polymorphism and disease progression in idiopathic pulmonary fibrosis. Am. J. Respir. Crit. Care Med. 2013, 188, 1442–1450. [Google Scholar] [CrossRef] [PubMed]
- The IPF Clinical Research Network. Prednisone, Azathioprine, and N-Acetylcysteine for Pulmonary Fibrosis. N. Engl. J. Med. 2012, 366, 1968–1977.
- King, T.E.; Albera, C.; Bradford, W.Z.; Costabel, U.; Hormel, P.; Lancaster, L.; Noble, P.W.; Sahn, S.A.; Szwarcberg, J.; Thomeer, M.; et al. Effect of interferon γ-1b on survival in patients with idiopathic pulmonary fibrosis (INSPIRE): A multicentre, randomised, placebo-controlled trial. Lancet 2009, 374, 222–228. [Google Scholar] [CrossRef]
- Raghu, G.; Brown, K.K.; Bradford, W.Z.; Starko, K.; Noble, P.W.; Schwartz, D.A.; King, T.E. Idiopathic Pulmonary Fibrosis Study Group. A placebo-controlled trial of interferon γ-1b in patients with idiopathic pulmonary fibrosis. N. Engl. J. Med. 2004, 350, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Li, X.; Zhang, L.; Cui, D.; Quan, X.; Yang, W. The anti-fibrotic effects of microRNA-153 by targeting TGFBR-2 in pulmonary fibrosis. Exp. Mol. Pathol. 2015, 99, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Pandit, K.V.; Milosevic, J. MicroRNA regulatory networks in idiopathic pulmonary fibrosis. Biochem. Cell Biol. Biochim. Biol. Cell. 2015, 93, 129–137. [Google Scholar] [CrossRef] [PubMed]
- Deng, N.; Sanchez, C.G.; Lasky, J.A.; Zhu, D. Detecting splicing variants in idiopathic pulmonary fibrosis from non-differentially expressed genes. PLoS ONE 2013, 8, e68352. [Google Scholar] [CrossRef] [PubMed]
- Larsson, O.; Diebold, D.; Fan, D.; Peterson, M.; Nho, R.S.; Bitterman, P.B.; Henke, C.A. Fibrotic myofibroblasts manifest genome-wide derangements of translational control. PLoS ONE 2008, 3, e3220. [Google Scholar] [CrossRef] [PubMed]
- Emblom-Callahan, M.C.; Chhina, M.K.; Shlobin, O.A.; Ahmad, S.; Reese, E.S.; Iyer, E.P.R.; Cox, D.N.; Brenner, R.; Burton, N.A.; Grant, G.M.; et al. Genomic phenotype of non-cultured pulmonary fibroblasts in idiopathic pulmonary fibrosis. Genomics 2010, 96, 134–145. [Google Scholar] [CrossRef] [PubMed]
- Bagnato, G.; Harari, S. Cellular interactions in the pathogenesis of interstitial lung diseases. Eur. Respir. Rev. 2015, 24, 102–114. [Google Scholar] [CrossRef] [PubMed]
- Campain, A.; Yang, Y.H. Comparison study of microarray meta-analysis methods. BMC Bioinform. 2010, 11, 408. [Google Scholar] [CrossRef] [PubMed]
- Tusher, V.G.; Tibshirani, R.; Chu, G. Significance analysis of microarrays applied to the ionizing radiation response. Proc. Natl. Acad. Sci. USA 2001, 98, 5116–5121. [Google Scholar] [CrossRef] [PubMed]
- Yasrebi, H. Comparative study of joint analysis of microarray gene expression data in survival prediction and risk assessment of breast cancer patients. Brief. Bioinform. 2016, 17, 771–785. [Google Scholar] [CrossRef] [PubMed]
- Tseng, G.C.; Ghosh, D.; Feingold, E. Comprehensive literature review and statistical considerations for microarray meta-analysis. Nucleic Acids Res. 2012, 40, 3785–3799. [Google Scholar] [CrossRef] [PubMed]
- Cheadle, C.; Vawter, M.P.; Freed, W.J.; Becker, K.G. Analysis of microarray data using z score transformation. J. Mol. Diagn. 2003, 5, 73–81. [Google Scholar] [CrossRef]
- PubMed-NCBI. Available online: http://www.ncbi.nlm.nih.gov/pubmed (accessed on 29 August 2016).
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Database for Annotation, Visualization, and Integrated Discovery (DAVID). Available online: https://david.ncifcrf.gov/ (accessed on 13 August 2016).
- Chen, J.; Xu, H.; Aronow, B.J.; Jegga, A.G. Improved human disease candidate gene prioritization using mouse phenotype. BMC Bioinform. 2007, 8. [Google Scholar] [CrossRef] [PubMed]
- TOPPFUN. Available online: https://toppgene.cchmc.org/enrichment.jsp (accessed on 13 August 2016).
- Hierarchical Clustering Explorer v3.5. Available online: http://www.cs.umd.edu/hcil/hce/ (accessed on 13 October 2015).
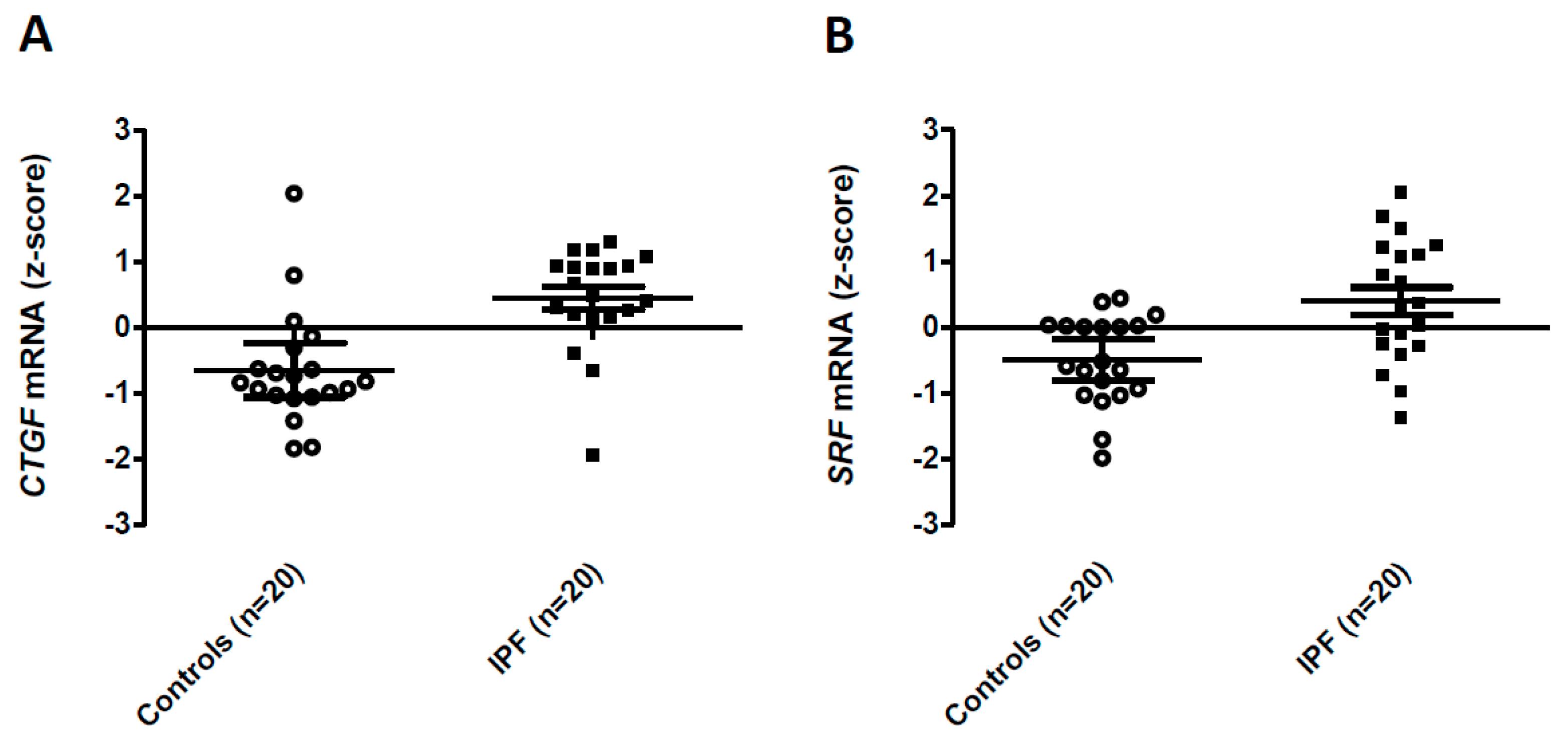
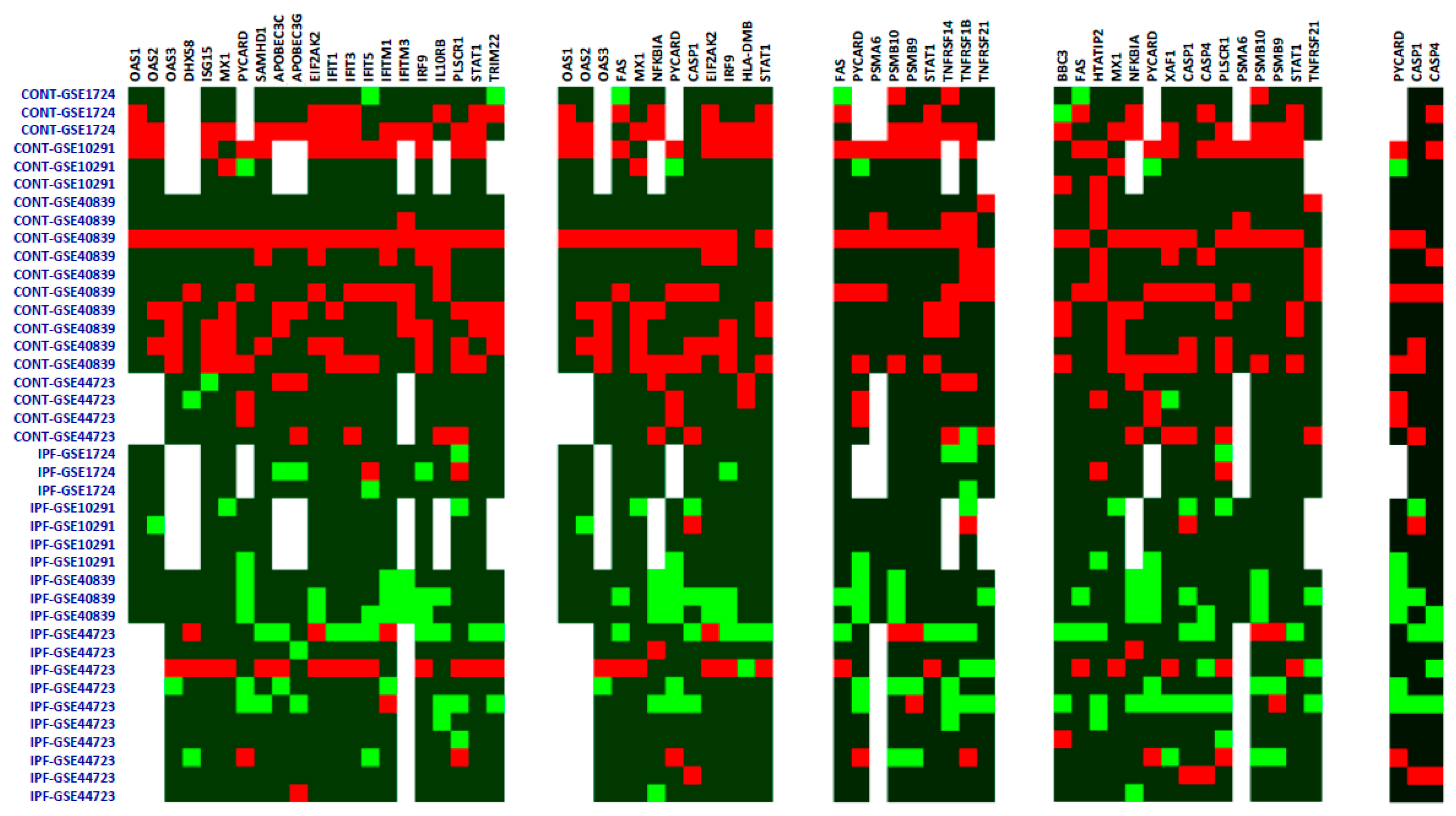
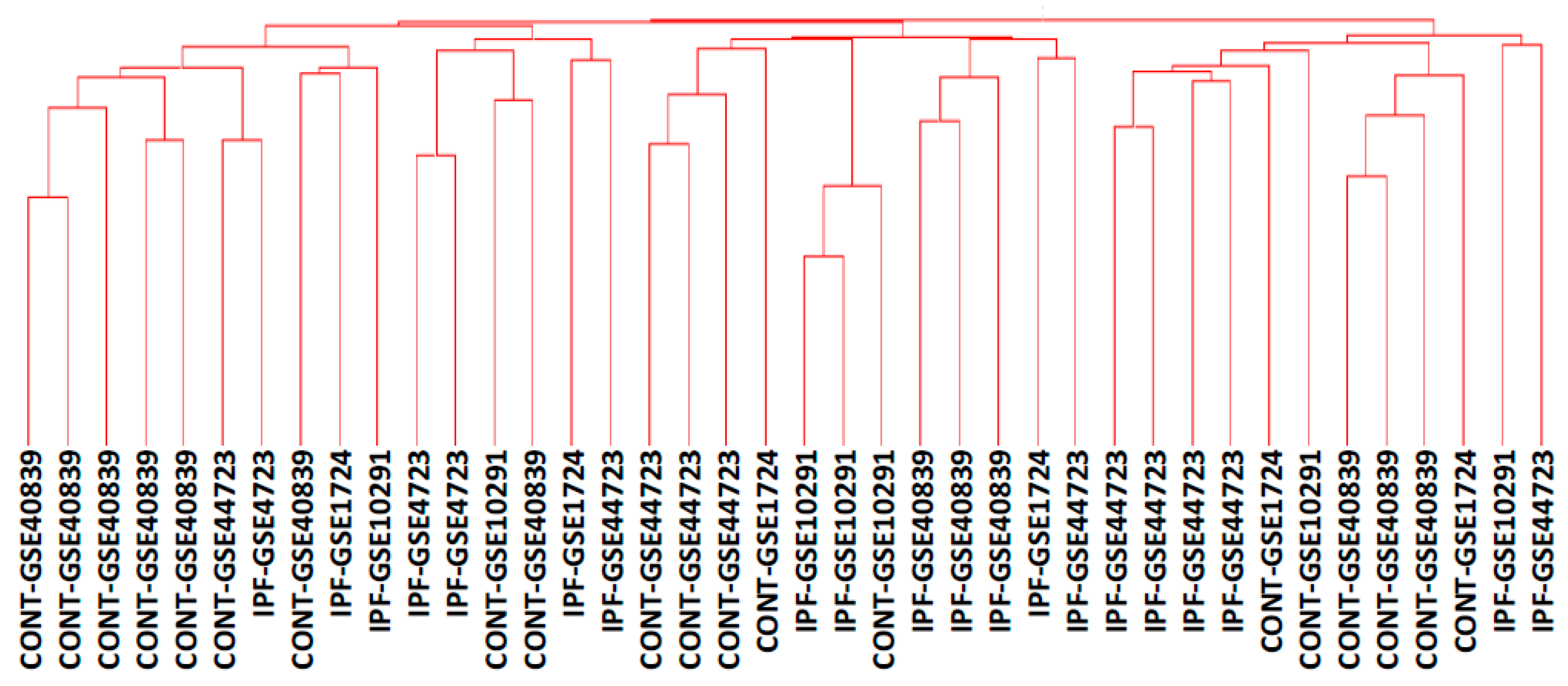

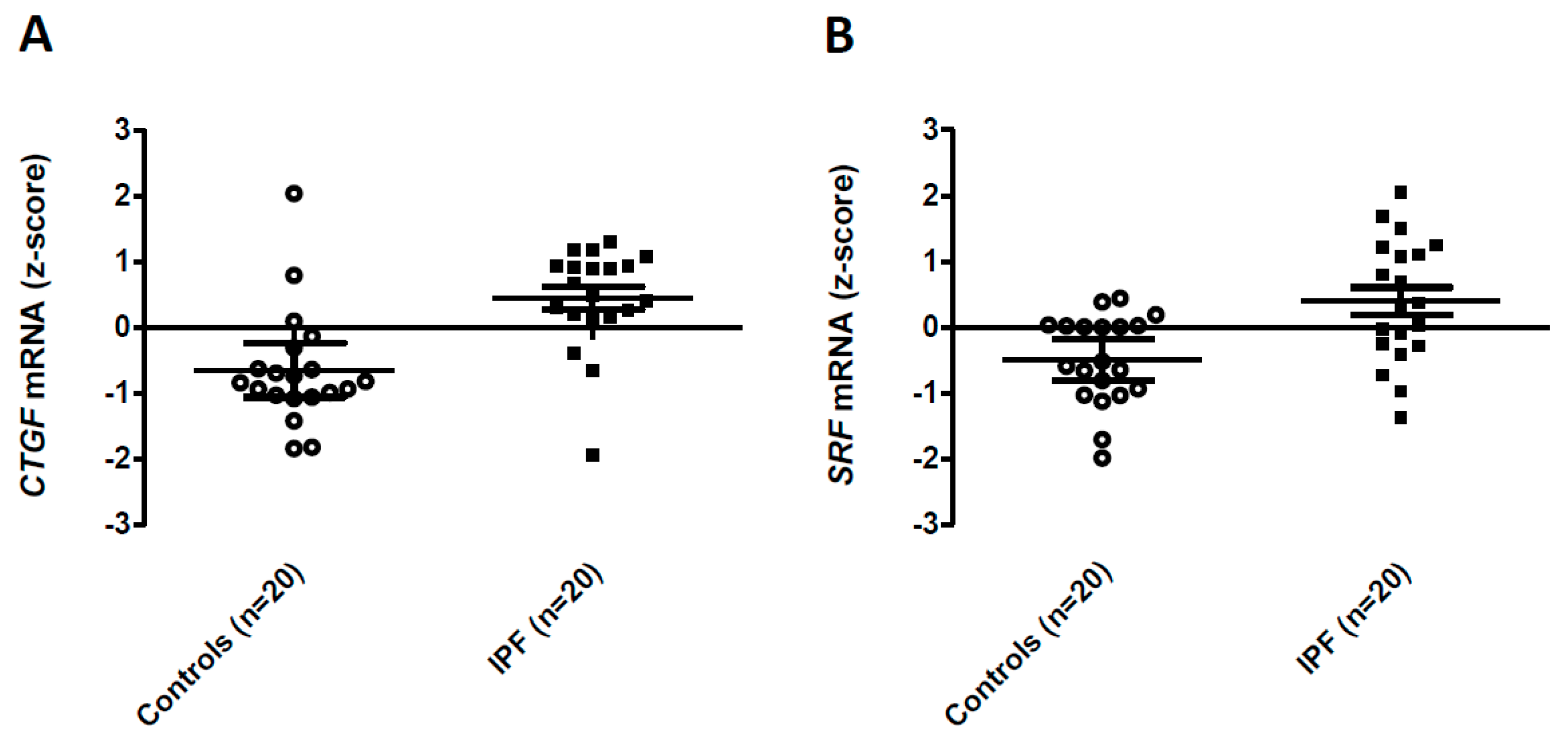{kind=link}
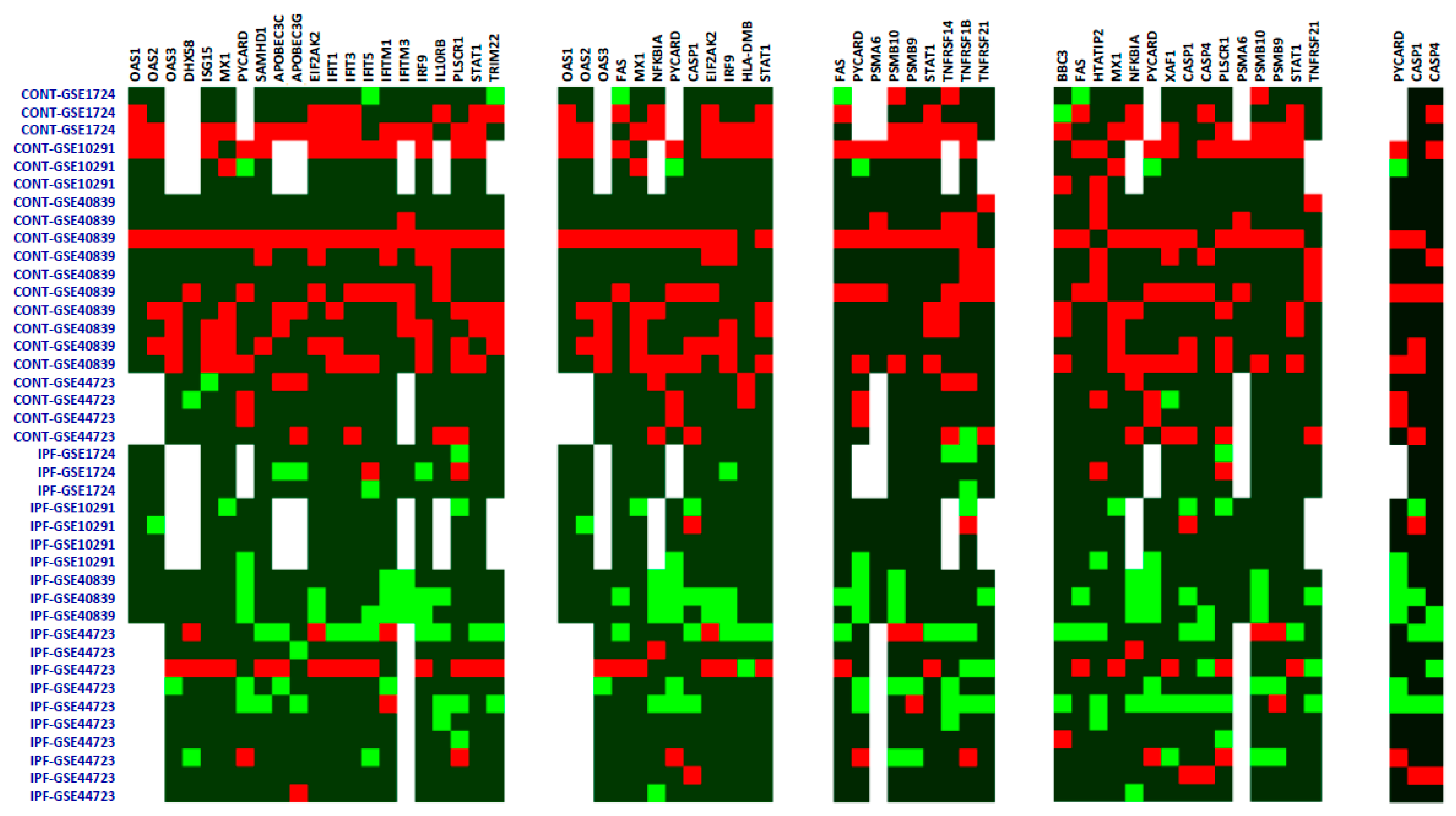{kind=link}
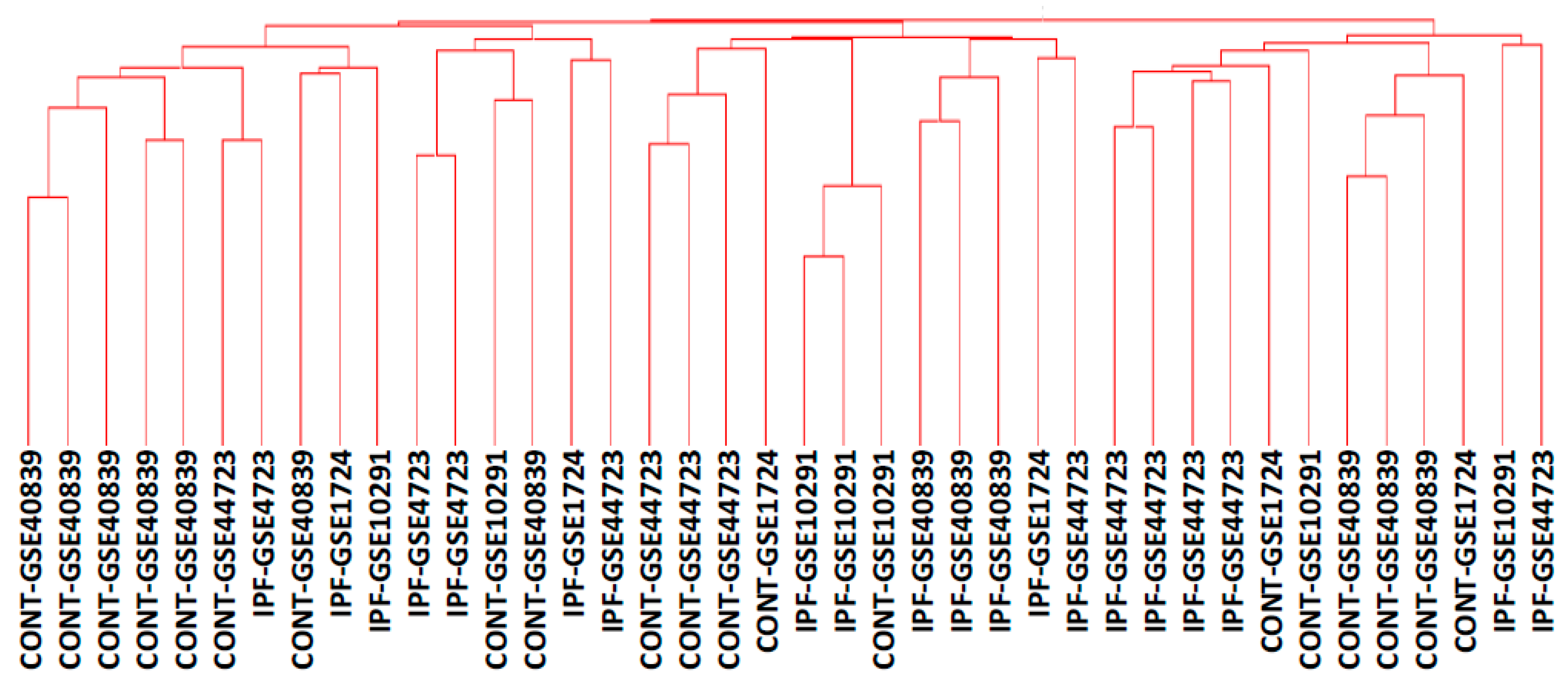{kind=link}
| Dataset | Number of Culture Passages | Number of Samples | Microarray Platform | Number of Probes | Normalization Method |
|---|---|---|---|---|---|
| GSE1724 | 4–5 | 3 controls, 3 IPF | Affymetrix human U95Av2 | 12,625 | N/A |
| GSE10921 | 4–7 | 3 controls, 4 IPF | Codelink Human Uniset I | 10,353 | Median |
| GSE44723 | Up to 11 | 4 controls, 10 IPF | Affymetrix HG-U133 plus 2.0 | 21,095 | RMA |
| GSE40839 | 2–5 | 10 controls, 3 IPF | Affymetrix human U133Av2 | 22,215 | Invariant set |
| Gene Symbol | Full Gene Name | Expression Level (z-Scores) | Implication in Fibrogenesis |
|---|---|---|---|
| Upregulated Genes | |||
| NAP1L3 | nucleosome assembly protein 1 like 3 | 1.49 | |
| PTHLH | parathyroid hormone like hormone | 1.10 | |
| KIAA0355 | N/A | 1.12 | |
| LIMS2 | LIM zinc finger domain containing 2 | 1.24 | Heart [11] |
| ASB1 | ankyrin repeat and SOCS box containing 1 | 1.16 | |
| HHAT | hedgehog acyltransferase | 1.32 | |
| EIF1 | eukaryotic translation initiation factor 1 | 1.04 | |
| PAWR | pro-apoptotic WT1 regulator | 1.15 | |
| NREP | neuronal regeneration related protein | 1.20 | Lung [12] |
| CTGF | connective tissue growth factor | 1.09 | Multiple [13] |
| Downregulated Genes | |||
| TRANK1 | tetratricopeptide repeat and ankyrin repeat containing 1 | −1.23 | |
| IFIT1 | interferon induced protein with tetratricopeptide repeats 1 | −1.20 | |
| SLC15A3 | solute carrier family 15 member 3 | −1.36 | |
| CPED1 | cadherin like and PC−esterase domain containing 1 | −1.38 | |
| PLSCR1 | phospholipid scramblase 1 | −1.13 | |
| IL1R1 | interleukin 1 receptor. type I | −1.12 | Skin [14] |
| IFI44 | interferon induced protein 44 | −1.11 | Lung [15] |
| HTATIP2 | HIV−1 Tat interactive protein 2 | −0.99 | |
| PLEKHA4 | pleckstrin homology domain containing A4 | −1.20 | |
| NFKBIA | NFκB inhibitor α | −1.22 | Liver [16] |
| Category | Source | Genes in List/Genes in Annotation | p Value (B&H) |
|---|---|---|---|
| Defense response to virus | Gene Ontology | 21/399 | 5.9 × 10−18 |
| Influenza A | KEGG | 12/100 | 9.8 × 10−6 |
| TNF mediated signaling pathway | Gene Ontology | 9/234 | 3.2 × 10−4 |
| Apoptotic process | Gene Ontology | 15/1314 | 1.5 × 10−3 |
| AIM2 inflammasome complex | Gene Ontology | 3/7 | 2.7 × 10−2 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Plantier, L.; Renaud, H.; Respaud, R.; Marchand-Adam, S.; Crestani, B. Transcriptome of Cultured Lung Fibroblasts in Idiopathic Pulmonary Fibrosis: Meta-Analysis of Publically Available Microarray Datasets Reveals Repression of Inflammation and Immunity Pathways. Int. J. Mol. Sci. 2016, 17, 2091. https://doi.org/10.3390/ijms17122091
Plantier L, Renaud H, Respaud R, Marchand-Adam S, Crestani B. Transcriptome of Cultured Lung Fibroblasts in Idiopathic Pulmonary Fibrosis: Meta-Analysis of Publically Available Microarray Datasets Reveals Repression of Inflammation and Immunity Pathways. International Journal of Molecular Sciences. 2016; 17(12):2091. https://doi.org/10.3390/ijms17122091
Chicago/Turabian StylePlantier, Laurent, Hélène Renaud, Renaud Respaud, Sylvain Marchand-Adam, and Bruno Crestani. 2016. "Transcriptome of Cultured Lung Fibroblasts in Idiopathic Pulmonary Fibrosis: Meta-Analysis of Publically Available Microarray Datasets Reveals Repression of Inflammation and Immunity Pathways" International Journal of Molecular Sciences 17, no. 12: 2091. https://doi.org/10.3390/ijms17122091