
Characterization of the Microenvironment of Nodular Lymphocyte Predominant Hodgkin Lymphoma



Abstract
:1. Introduction
2. Results
2.1. Comparison of Total Cell Populations between NLPHL and RLN
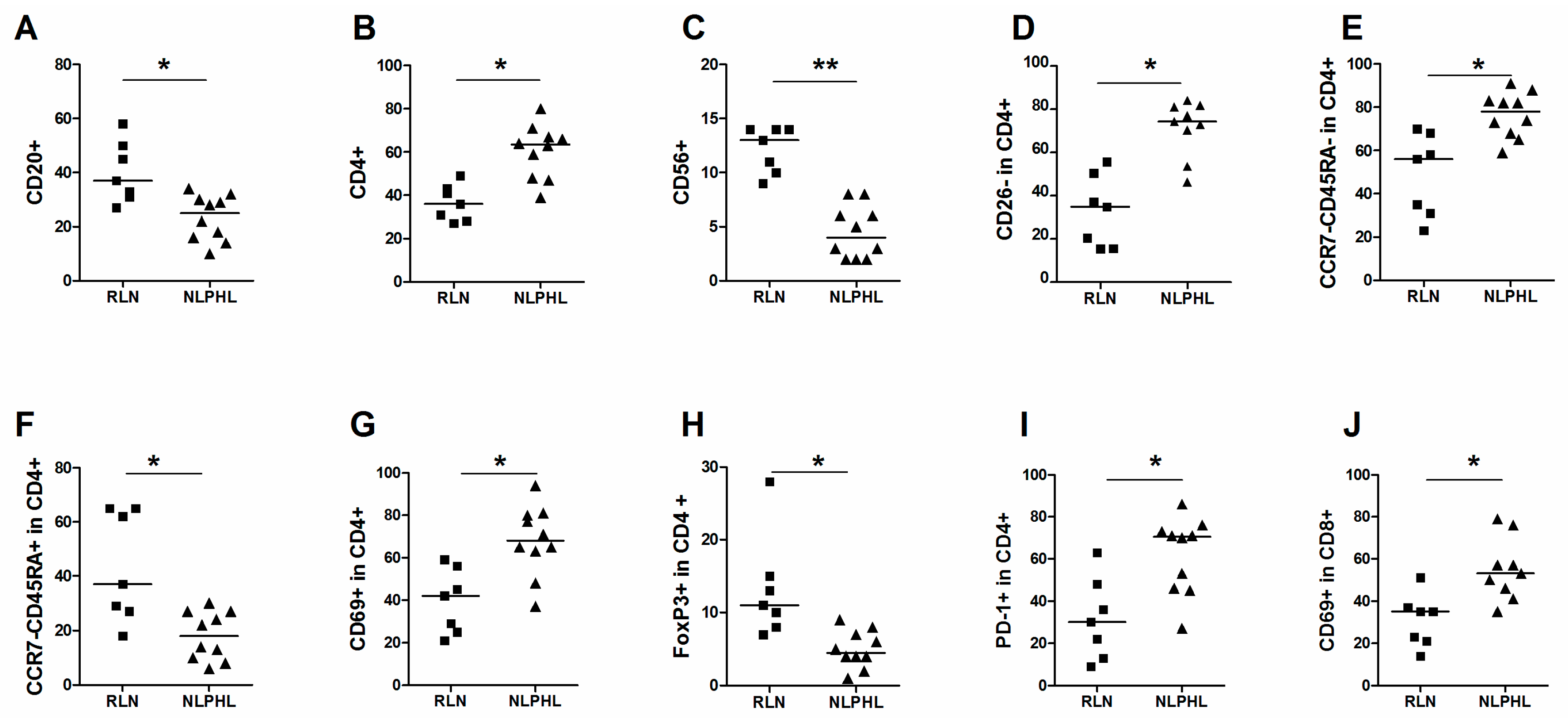
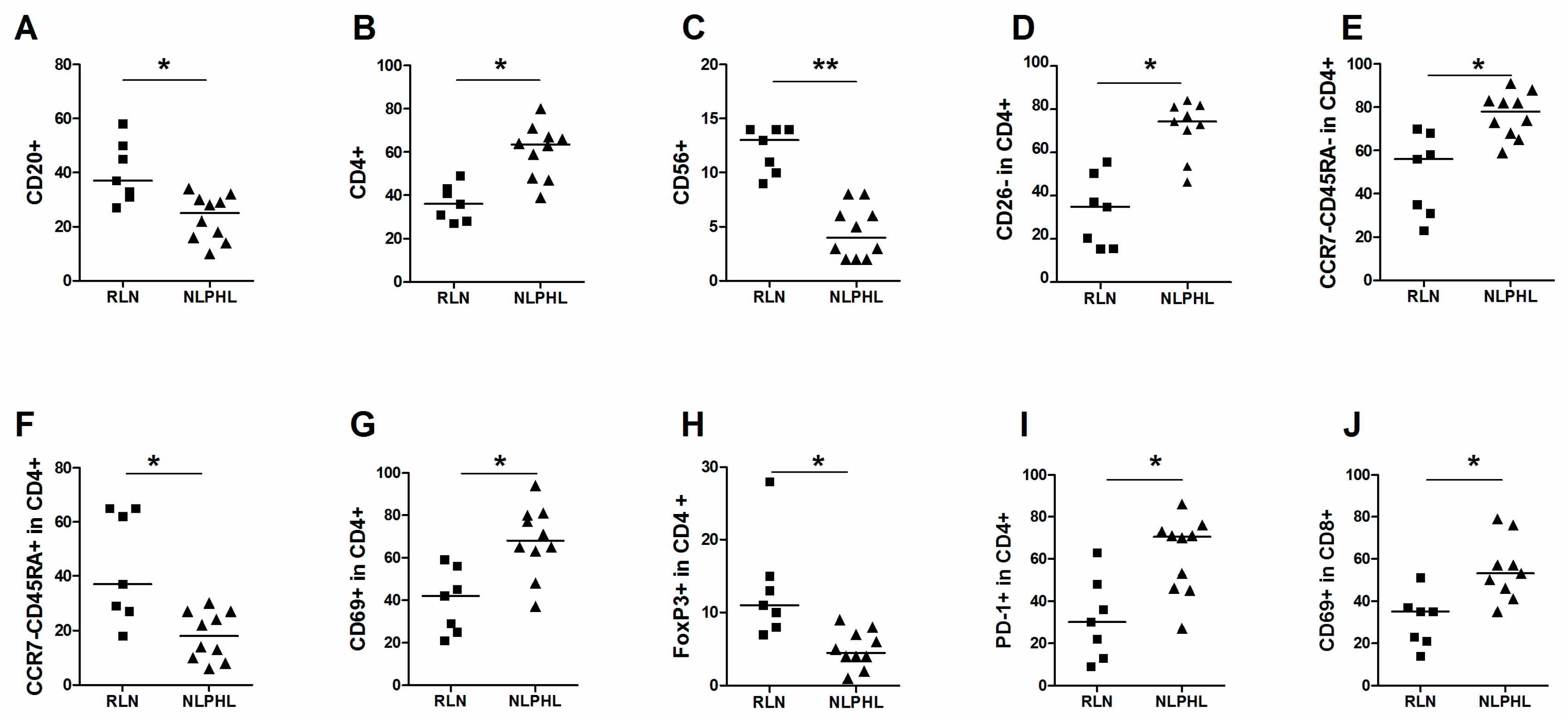
2.2. Comparison of Subpopulations between NLPHL and RLN
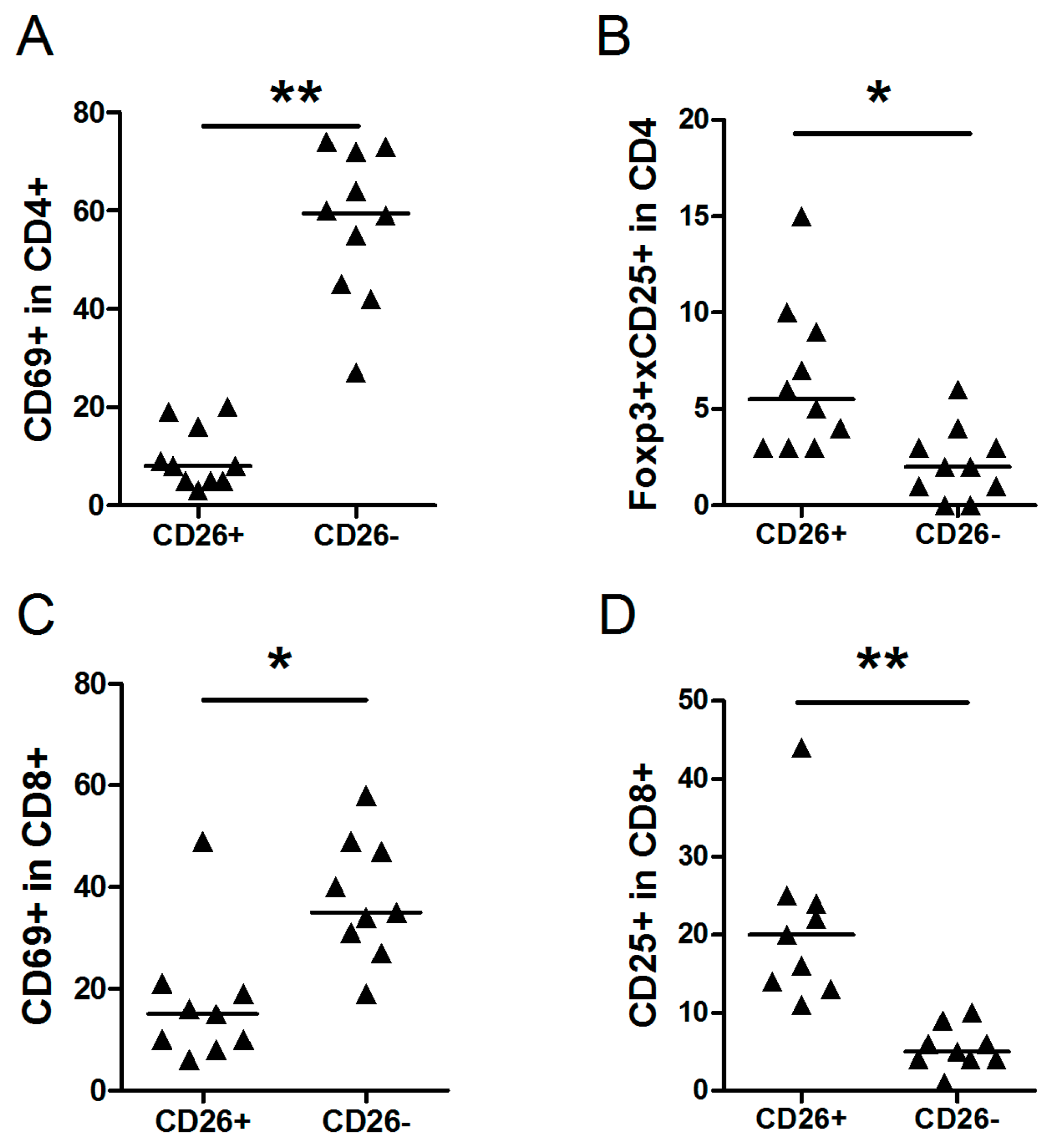
2.3. Comparison of Subpopulations of Cells within CD26- and CD26+ of NLPHL
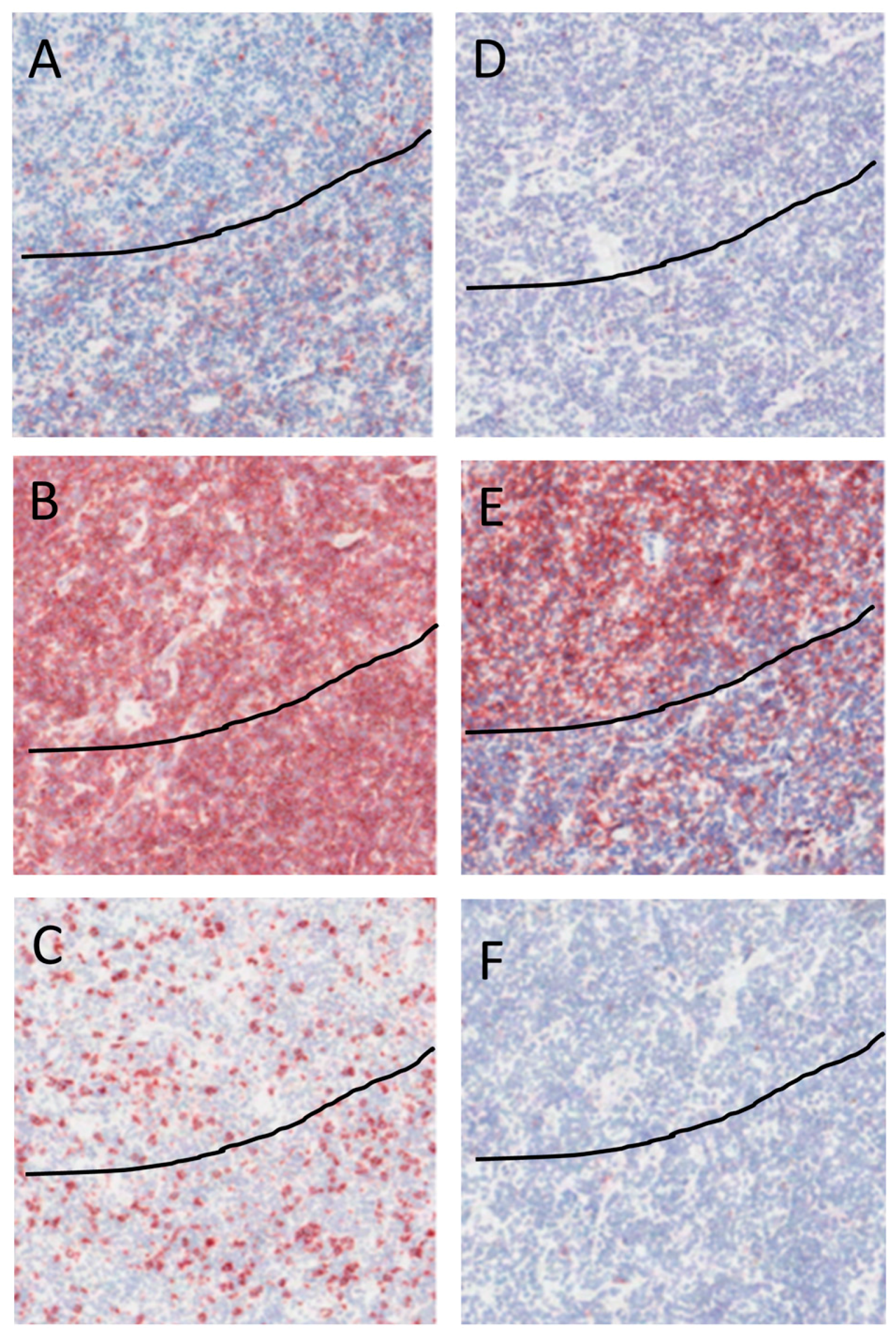
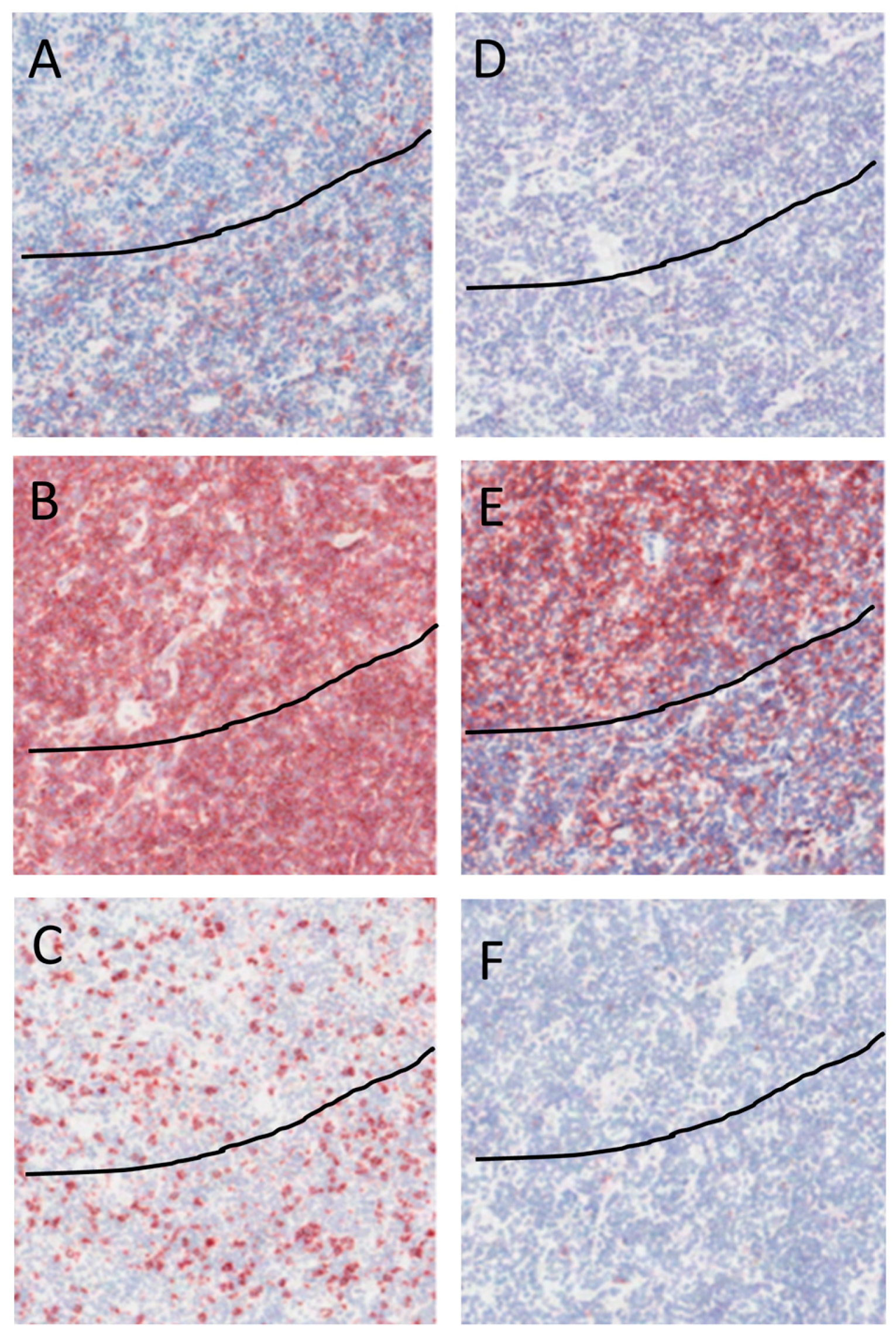
2.4. Immunohistochemical Staining of NLPHL
3. Discussion
4. Methods and Materials
4.1. Patients
4.2. Flow Cytometry
4.3. Immunohistochemistry
4.4. Statistics
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Swerdlow, S.H.; Campo, E.; Harris, N.L.; Jaffe, E.S.; Pileri, S.A.; Stein, H.; Thiele, J.; Vardiman, J.W. WHO Classification of Tumours of Haematopoietic and Lymphoid Tissues, 4th ed.; World Health Organization: Geneva, Switzerland, 2008; pp. 323–325. [Google Scholar]
- Liu, Y.; Sattarzadeh, A.; Diepstra, A.; Visser, L.; van den Berg, A. The microenvironment in classical Hodgkin lymphoma: An actively shaped and essential tumor component. Semin. Cancer Biol. 2014, 24, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Atayar, C.; Poppema, S.; Visser, L.; van den Berg, A. Cytokine gene expression profile distinguishes CD4+/CD57+ T-cells of nodular lymphocyte predominance type of Hodgkin lymphoma from their tonsillar counterparts. J. Pathol. 2006, 208, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Nam-Cha, S.H.; Roncador, G.; Sanchez-Verde, L.; Montes-Moreno, S.; Acevedo, A.; Dominguez-Franjo, P.; Piris, M.A. PD-1, a follicular T-cell marker useful for recognizing nodular lymphocyte-predominant Hodgkin lymphoma. Am. J. Surg. Pathol. 2008, 32, 1252–1257. [Google Scholar] [CrossRef] [PubMed]
- Sattarzadeh, A.; Diepstra, A.; Rutgers, B.; van den Berg, A.; Visser, L. CD57+ T-cells are a subpopulation of T-follicular helper cells in nodular lymphocyte predominant Hodgkin lymphoma. Exp. Hematol. Oncol. 2015, 4, 27. [Google Scholar] [CrossRef] [PubMed]
- Rahemtullah, A.; Harris, N.L.; Dorn, M.E.; Preffer, F.I.; Hasserjian, R.P. Beyond the lymphocyte predominant cell: CD4+CD8+ T-cells in nodular lymphocyte predominant Hodgkin lymphoma. Leuk Lymphoma 2008, 49, 1870–1878. [Google Scholar] [CrossRef] [PubMed]
- Bosler, D.S.; Douglas-Nikitin, V.K.; Harris, V.N.; Smith, M.D. Detection of T-regulatory cells has a potential role in the diagnosis of classical Hodgkin lymphoma. Cytometry B Clin. Cytom. 2008, 74, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Quesada, A.E.; Assylbekova, B.; Jabcuga, C.E.; Zhang, R.; Covinsky, M.; Rios, A.; Nguyen, N.D.; Brown, R.E. Expression of Sirt1 and FoxP3 in classical Hodgkin lymphoma and tumor infiltrating lymphocytes: Implications for immune dysregulation, prognosis and potential therapeutic targeting. Int. J. Clin. Exp. Pathol. 2015, 8, 13241–13248. [Google Scholar] [PubMed]
- Greaves, P.; Clear, A.; Owen, A.; Iqbal, S.; Lee, A.; Matthews, J.; Wilson, A.; Calaminici, M.; Gribben, J.G. Defining characteristics of classical Hodgkin lymphoma microenvironment T-helper cells. Blood 2013, 122, 2856–2863. [Google Scholar] [CrossRef] [PubMed]
- Poppema, S.; van den Berg, A. Interaction between host T cells and Reed-Sternberg cells in Hodgkin lymphomas. Seminars Cancer Biol. 2000, 10, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Wu, D.; Thomas, A.; Fromm, J.R. Reactive T cells by flow cytometry distinguish Hodgkin lymphomas from T cell/histiocyte-rich large B cell lymphoma. Cytometry B Clin. Cytom. 2016, 90, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Brune, V.; Tiacci, E.; Pfeil, I.; Doring, C.; Eckerle, S.; van Noesel, C.J.; Klapper, W.; Falini, B.; von Heydebreck, A.; Metzler, D.; et al. Origin and pathogenesis of nodular lymphocyte-predominant Hodgkin lymphoma as revealed by global gene expression analysis. J. Exp. Med. 2008, 205, 2251–2268. [Google Scholar] [CrossRef] [PubMed]
- Leonardo, S.M.; De Santis, J.L.; Gehrand, A.; Malherbe, L.P.; Gauld, S.B. Expansion of follicular helper T cells in the absence of Treg cells: implications for loss of B-cell anergy. Eur. J. Immunol. 2012, 42, 2597–2607. [Google Scholar] [CrossRef] [PubMed]
- Hartmann, S.; Doring, C.; Jakobus, C.; Rengstl, B.; Newrzela, S.; Tousseyn, T.; Sagaert, X.; Ponzoni, M.; Facchetti, F.; de Wolf-Peeters, C.; et al. Nodular lymphocyte predominant Hodgkin lymphoma and T cell/histiocyte rich large B cell lymphoma–endpoints of a spectrum of one disease? PLoS ONE 2013, 8, e78812. [Google Scholar] [CrossRef] [PubMed]
- Weber, J.P.; Fuhrmann, F.; Feist, R.K.; Lahmann, A.; Al Baz, M.S.; Gentz, L.J.; Vu Van, D.; Mages, H.W.; Haftmann, C.; Riedel, R.; et al. ICOS maintains the T follicular helper cell phenotype by down-regulating Kruppel-like factor 2. J. Exp. Med. 2015, 212, 217–233. [Google Scholar] [CrossRef] [PubMed]
- Leavenworth, J.W.; Verbinnen, B.; Yin, J.; Huang, H.; Cantor, H. A p85α-osteopontin axis couples the receptor ICOS to sustained BCL-6 expression by follicular helper and regulatory T cells. Nat. Immunol. 2015, 16, 96–106. [Google Scholar] [CrossRef] [PubMed]
- Ansell, S.M.; Lesokhin, A.M.; Borrello, I.; Halwani, A.; Scott, E.C.; Gutierrez, M.; Schuster, S.J.; Millenson, M.M.; Cattry, D.; Freeman, G.J.; et al. PD-1 blockade with nivolumab in relapsed or refractory Hodgkin’s lymphoma. N. Engl. J. Med. 2015, 372, 311–319. [Google Scholar] [CrossRef] [PubMed]
- Chen, B.J.; Chapuy, B.; Ouyang, Y.; Sun, H.H.; Roemer, M.G.M.; Xu, M.L.; Yu, H.; Fletcher, C.D.M.; Freeman, G.J.; Shipp, M.A.; et al. PD-L1 expression is characteristic of a subset of aggressive B-cell lymphomas and virus-associated malignancies. Clin. Cancer Res. 2013, 19, 3462–3473. [Google Scholar] [CrossRef] [PubMed]
- Yang, Z.-Z.; Grote, D.M.; Ziesmer, S.C.; Xiu, B.; Novak, A.J.; Ansell, S.M. PD-1 expression defines two distinct T-cell sub-populations in follicular lymphoma that differentially impact patient survival. Blood Cancer J. 2015, 5, e281. [Google Scholar] [CrossRef] [PubMed]
- Gerlach, U.A.; Vogt, K.; Schlickeiser, S.; Meisel, C.; Streitz, M.; Kunkel, D.; Appelt, C.; Ahrlich, S.; Lachmann, N.; Neuhaus, P.; et al. Elevation of CD4+ differentiated memory T cells is associated with acute cellular and antibody-mediated rejection after liver transplantation. Transplantation 2013, 95, 1512–1520. [Google Scholar] [CrossRef] [PubMed]
- Sallusto, F.; Lenig, D.; Forster, R.; Lipp, M.; Lanzavecchia, A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature 1999, 401, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Schenkel, J.M.; Masopust, D. Tissue-resident memory T cells. Immunity 2014, 41, 886–897. [Google Scholar] [CrossRef] [PubMed]
- Shiow, L.R.; Rosen, D.B.; Brdickova, N.; Xu, Y.; An, J.; Lanier, L.L.; Cyster, J.G.; Matloubian, M. CD69 acts downstream of interferon-α/β to inhibit S1P1 and lymphocyte egress from lymphoid organs. Nature 2006, 440, 540–544. [Google Scholar] [CrossRef] [PubMed]
- Sancho, D.; Gomez, M.; Viedma, F.; Esplugues, E.; Gordon-Alonso, M.; Garcia-Lopez, M.A.; de la Fuente, H.; Martinez-A, C.; Lauzurica, P.; Sanchez-Madrid, F. CD69 downregulates autoimmune reactivity through active transforming growth factor-β production in collagen-induced arthritis. J. Clin. Investig. 2003, 112, 872–882. [Google Scholar] [CrossRef] [PubMed]
- Esplugues, E.; Sancho, D.; Vega-Ramos, J.; Martinez, C.; Syrbe, U.; Hamann, A.; Engel, P.; Sanchez-Madrid, F.; Lauzurica, P. Enhanced antitumor immunity in mice deficient in CD69. J. Exp. Med. 2003, 197, 1093–1106. [Google Scholar] [CrossRef] [PubMed]
- Wu, R.; Sattarzadeh, A.; Rutgers, B.; Diepstra, A.; van den Berg, A.; Visser, L. The microenvironment of classical Hodgkin lymphoma: heterogeneity by Epstein-Barr virus presence and location within the tumor. Blood Cancer J. 2016, 6, e417. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
{kind=link}
| Cell Type | Population | Median % (Range) | p-Value | |||
|---|---|---|---|---|---|---|
| RLN | NLPHL | |||||
| Main cell populations | CD20+ in live | 37 | 27–58 | 25 | 10–34 | 0.007 |
| CD3+ in live | 57 | 39–65 | 76 | 46–89 | 0.017 | |
| CD4+ in live | 36 | 29–51 | 62 | 41–71 | 0.002 | |
| CD8+ in live | 22 | 15–40 | 20 | 13–27 | 0.1 | |
| CD56+ in live | 13 | 9–14 | 4 | 2–8 | 0.00005 | |
| CD68+ in live | 2 | 1–2 | 1.5 | 1–3 | 1 | |
| CD4+CD8+ in live | 3 | 2–9 | 8 | 3–17 | 0.016 | |
| CD26- | CD26- in CD4+ | 35 | 15–56 | 74 | 46–84 | 0.0012 |
| CD26- in CD8+ | 34 | 26–61 | 34 | 26–72 | 0.2439 | |
| Maturation of CD4+ | Naive (CCR7+CD45RA+ in CD4+) | 6 | 3–11 | 2.5 | 1–7 | 0.016 |
| TCM (CCR7+CD45RA- in CD4+) | 1 | 0–2 | 2 | 1–8 | 0.08 | |
| TEM (CCR7-CD45RA- in CD4+) | 56 | 23–70 | 78 | 59–91 | 0.002 | |
| TEMRA (CCR7-CD45RA+ in CD4+) | 37 | 18–65 | 18 | 6–30 | 0.007 | |
| Activation of CD4+ | CD69+ in CD4+ | 42 | 21–59 | 68 | 37–94 | 0.003 |
| CD25+ in CD4+ | 10 | 5–21 | 11 | 4–18 | 0.5 | |
| Th1 | CXCR3+ in CD4+ | 18 | 11–56 | 15 | 8–33 | 0.3 |
| Th2 | ST2L+ in CD4+ | 14 | 4–23 | 9 | 4–18 | 0.3 |
| CXCR4+ in CD4+ | 5 | 3–14 | 6 | 3–9 | 1 | |
| Treg | GITR+ in CD4+ | 17 | 10–66 | 33 | 15–43 | 0.3 |
| GITR+CD25+ in CD4+ | 6 | 2–11 | 8.5 | 5–17 | 0.2 | |
| CD127- in CD4+ | 15 | 5–31 | 24 | 19–56 | 0.02 | |
| CD127-CD25+ in CD4+ | 2 | 1–3 | 1 | 1–4 | 0.8 | |
| CD152+ in CD4+ | 8 | 5–17 | 6.5 | 3–9 | 0.3 | |
| CD152+CD25+ in CD4+ | 7 | 3–13 | 4.5 | 2–6 | 0.4 | |
| FoxP3+ in CD4+ | 11 | 7–28 | 4.5 | 1–9 | 0.001 | |
| CD25+FoxP3+ in CD4+ | 4 | 2–11 | 2.5 | 1–5 | 0.1 | |
| CD25+CD45RA- inCD4+ | 4 | 0–6 | 4 | 0–11 | 0.1 | |
| TFH | CD57+ in CD4+ | 8 | 3–22 | 21 | 3–39 | 0.1 |
| PD-1+ in CD4+ | 30 | 9–63 | 71 | 27–86 | 0.009 | |
| PD-1+CD57+ in CD4+ | 7 | 2–21 | 20 | 2–38 | 0.2 | |
| CXCR5+ICOS+ in CD4+ | 8 | 5–27 | 5 | 1–15 | 0.017 | |
| CXCR5+ X ICOS+ in CD25+/CD4+ | 31 | 18–50 | 26 | 11–36 | 0.4 | |
| Bcl6+ in CD4+ | 15 | 3–35 | 25 | 4–40 | 0.2 | |
| CXCR5+BCL6+ in CD4+ | 12 | 2–30 | 13 | 1–30 | 0.7 | |
| Bcl6+CD57+ in CD4+ | 7 | 3–15 | 11 | 3–15 | 0.1 | |
| Cytotoxic CD4+ | TIA-1+ in CD4+ | 7 | 1–13 | 15 | 3–32 | 0.012 |
| Granzyme-B+ in CD4+ | 2 | 1–5 | 3 | 1–4 | 0.8 | |
| Activation of CD8+ | CD25+ in CD8+ | 30 | 12–32 | 27 | 14–51 | 0.6 |
| CD69+ in CD8+ | 35 | 14–51 | 53 | 35–79 | 0.005 | |
| CD8+ | CXCR4+ in CD8+ | 30 | 21–44 | 33.5 | 10–78 | 0.8 |
| CXCR3+ in CD8+ | 41 | 20–63 | 33 | 21–44 | 0.2 | |
| TIA-1+ in CD8+ | 25 | 19–49 | 40.5 | 28–72 | 0.03 | |
| Granzyme-B+ in CD8+ | 6 | 4–27 | 11 | 4–21 | 0.04 | |
| NK/NKT | CD56+ in CD3- | 17 | 15–26 | 4 | 1–22 | 0.8 |
| CD16+ in CD3- | 13 | 6–24 | 8.5 | 3–21 | 0.2 | |
| CD57+ in CD3- | 4 | 1–8 | 5 | 3–16 | 0.1 | |
| CD56+CD16+ in CD3- | 4 | 1–11 | 1.5 | 1–6 | 0.1 | |
| CD56+CD107a+ in CD3- | 1 | 1–1 | 1 | 0–2 | 0.2 | |
| CD56+CD16+ in CD3+ | 4 | 2–10 | 2 | 1–5 | 0.04 | |
| Macrophage | CD163+ in CD68+ | 17 | 7–32 | 19 | 12–36 | 0.1 |
| Cell Type | Median % [Range] | p-Value | |||
|---|---|---|---|---|---|
| CD26+ | CD26- | ||||
| CD69+ in CD4+ | 8 | 3–20 | 60 | 27–74 | 0.00001 |
| CD25+ in CD4+ | 5 | 2–11 | 3 | 1–7 | 0.2 |
| FoxP3+ in CD4+ | 2 | 1–3 | 4 | 0–7 | 0.1 |
| Foxp3+CD25+ in CD4+ | 5 | 3–15 | 2 | 0–6 | 0.003 |
| CXCR3+ in CD4+ | 4 | 2–12 | 7 | 3–21 | 0.017 |
| ST2L+ in CD4+ | 3.5 | 2–6 | 4.4 | 2–12 | 0.3 |
| CXCR3+ in CD8+ | 17 | 10–29 | 19 | 10–29 | 0.5 |
| CD69+ in CD8+ | 15 | 6–49] | 34 | 19–58 | 0.004 |
| CD25+ in CD8+ | 20 | 11–44 | 5 | 1–10 | 0.00004 |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sattarzadeh, A.; Visser, L.; Rutgers, B.; Diepstra, A.; Berg, A.V.d. Characterization of the Microenvironment of Nodular Lymphocyte Predominant Hodgkin Lymphoma. Int. J. Mol. Sci. 2016, 17, 2127. https://doi.org/10.3390/ijms17122127
Sattarzadeh A, Visser L, Rutgers B, Diepstra A, Berg AVd. Characterization of the Microenvironment of Nodular Lymphocyte Predominant Hodgkin Lymphoma. International Journal of Molecular Sciences. 2016; 17(12):2127. https://doi.org/10.3390/ijms17122127
Chicago/Turabian StyleSattarzadeh, Ahmad, Lydia Visser, Bea Rutgers, Arjan Diepstra, and Anke Van den Berg. 2016. "Characterization of the Microenvironment of Nodular Lymphocyte Predominant Hodgkin Lymphoma" International Journal of Molecular Sciences 17, no. 12: 2127. https://doi.org/10.3390/ijms17122127