
The Protective Effect of Grape-Seed Proanthocyanidin Extract on Oxidative Damage Induced by Zearalenone in Kunming Mice Liver

 ,
,
Abstract
:
1. Introduction
2. Results
2.1. Effects of Aspartate Aminotransferase (AST) and Alanine Aminotransferase (ALT) on the Serum Activities
2.2. The Effect on Antioxidant of Liver
2.3. Effect on Histopathological Variation in Liver
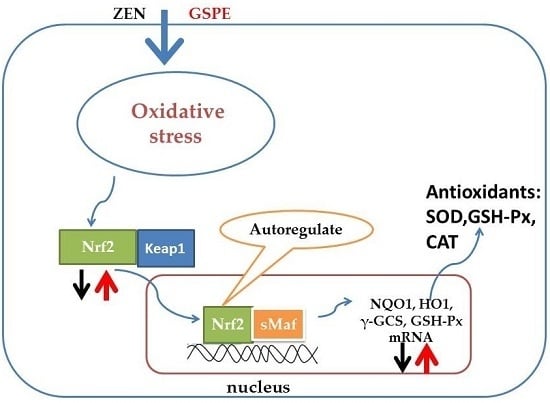
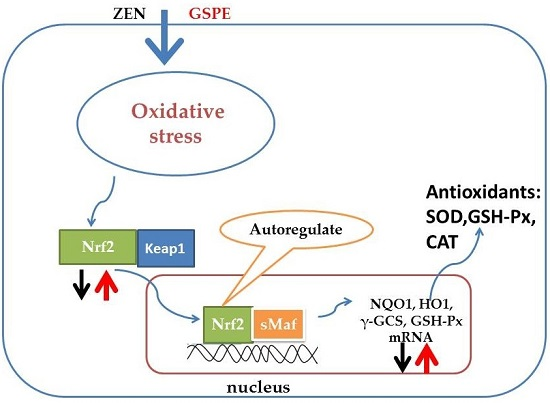
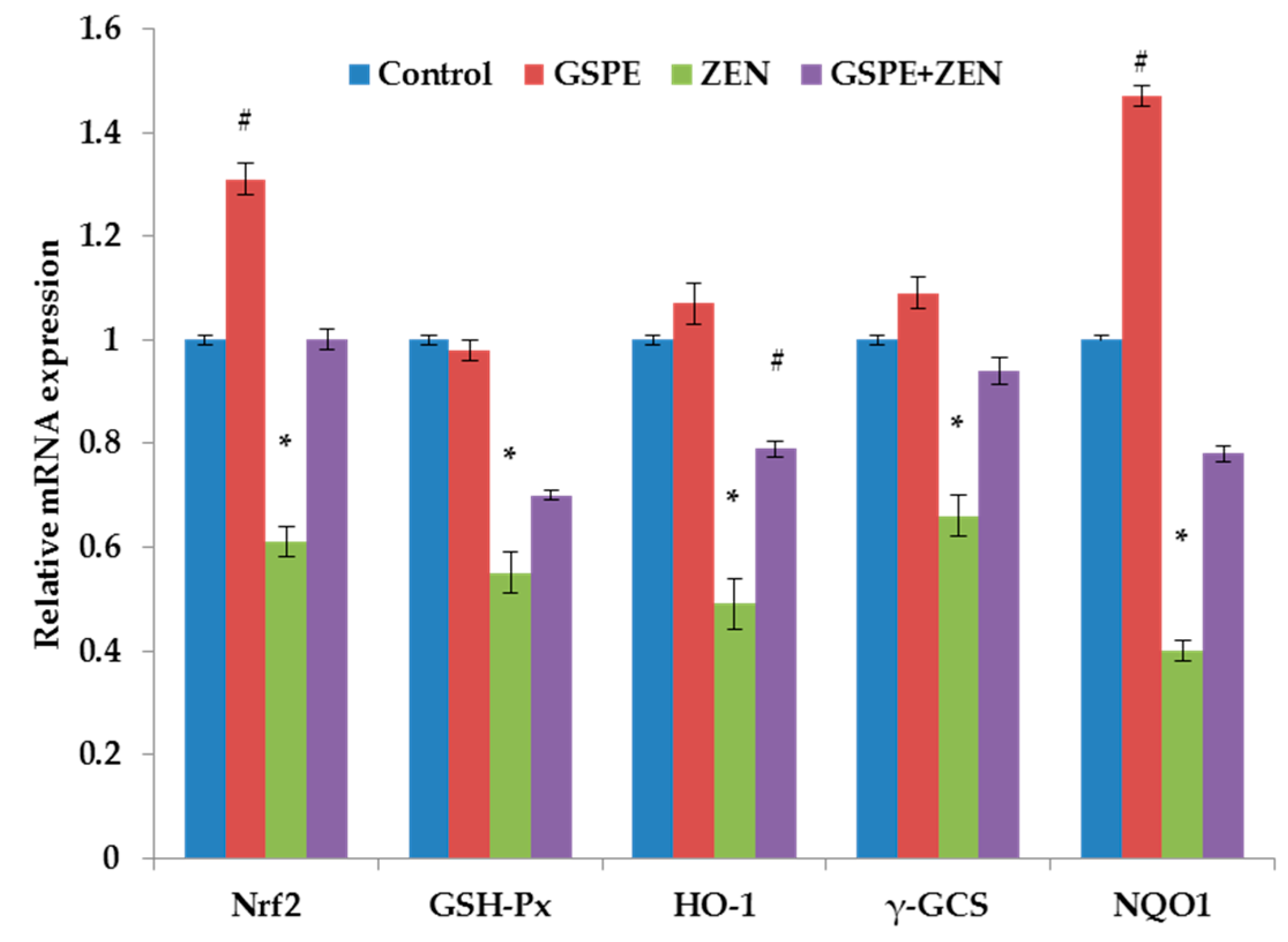
2.4. Effect on the Gene Expression Associated with the Nrf2 Signaling Pathway
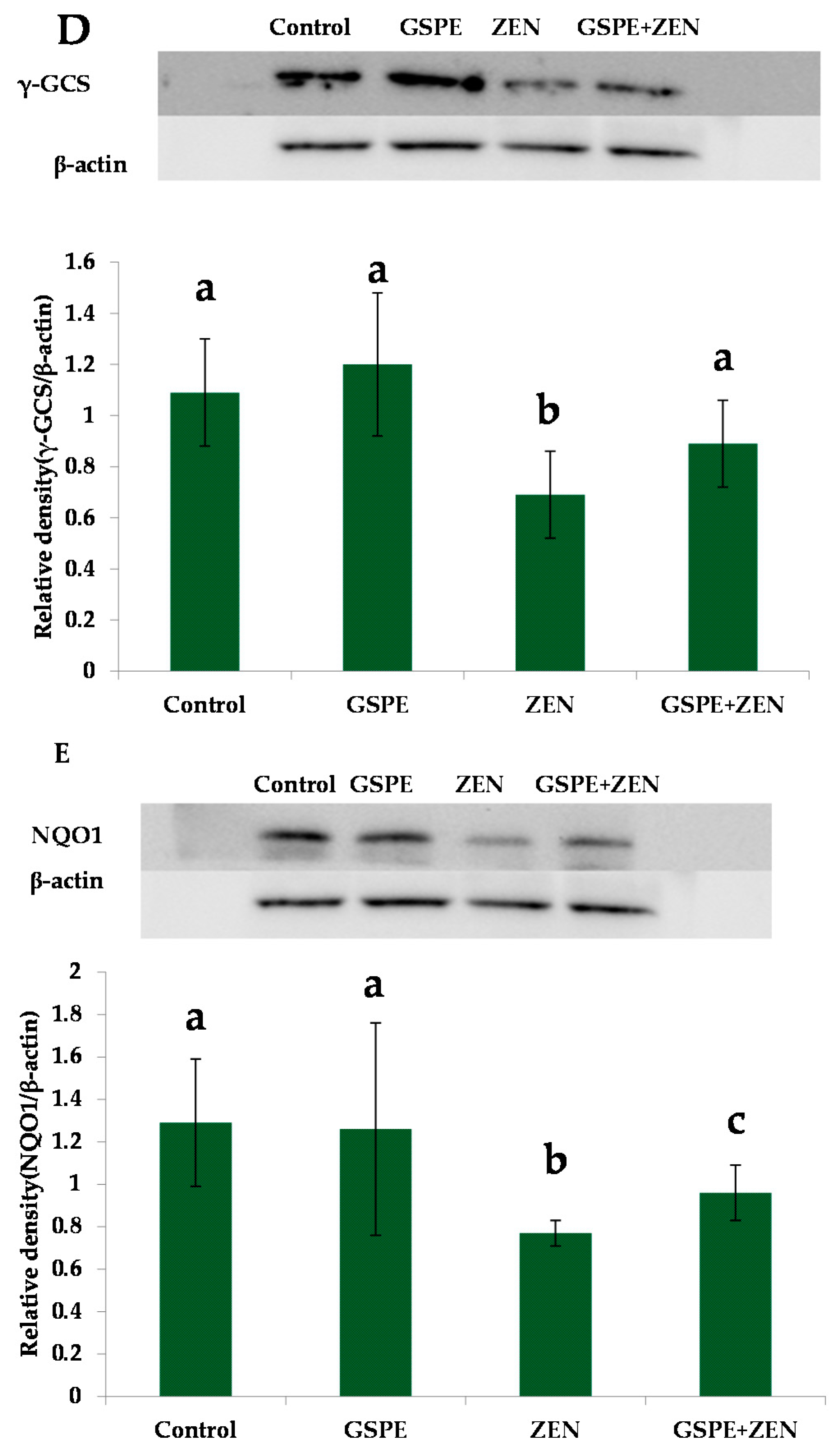
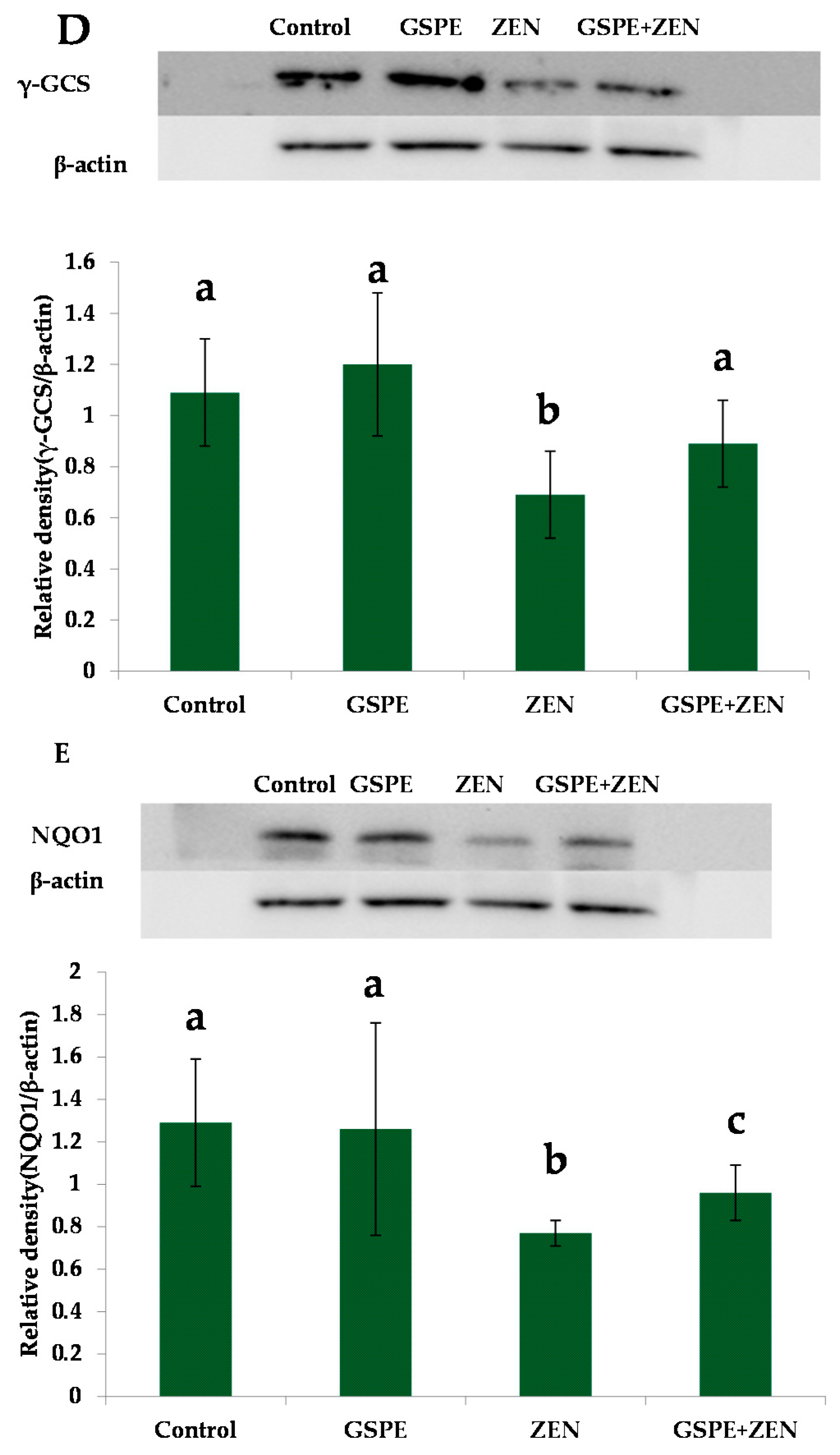
2.5. Effect on Nrf2 Signaling-Related Protein Expressions
3. Discussion
4. Experimental Section
4.1. Animals
4.2. Chemicals
4.3. Experimental Design and Treatment
4.4. Parameters
4.5. Histopathology Examination of Liver Tissue
4.6. Gene Expression
4.7. Western Blot Assay
4.8. Statistical Analysis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Savard, C.; Nogues, P.; Boyer, A.; Chorfi, Y. Prevention of deoxynivalenol- and zearalenone-associated oxidative stress does not restore MA-10 Leydig cell functions. Toxicology 2016, 341–343, 17–27. [Google Scholar] [CrossRef] [PubMed]
- Abid-Essefi, S.; Zaied, C.; Bouaziz, C.; Salem, I.B.; Kaderi, R.; Bacha, H. Protective effect of aqueous extract of Allium sativum against zearalenone toxicity mediated by oxidative stress. Exp. Toxicol. Pathol. 2012, 64, 689–695. [Google Scholar] [CrossRef] [PubMed]
- Koraichi, F.; Videmann, B.; Mazallon, M.; Benahmed, M.; Prouillac, C.; Lecoeur, S. Zearalenone exposure modulates the expression of ABC transporters and nuclear receptors in pregnant rats and fetal liver. Toxicol. Lett. 2012, 211, 246–256. [Google Scholar] [CrossRef] [PubMed]
- Jiang, S.Z.; Yang, Z.B.; Yang, W.R.; Gao, J.; Liu, F.X.; Broomhead, J.; Chi, F. Effects of purified zearalenone on growth performance, organ size, serum metabolites, and oxidative stress in postweaning gilts. J. Anim. Sci. 2011, 89, 3008–3015. [Google Scholar] [CrossRef] [PubMed]
- Maaroufi, K.; Chekir, L.; Creppy, E.E.; Ellouz, F.; Bacha, H. Zearalenone induces modifications of haematological and biochemical parameters in rats. Toxicon 1996, 34, 535–540. [Google Scholar] [CrossRef]
- Sun, L.H.; Lei, M.Y.; Zhang, N.Y.; Gao, X.; Li, C.; Krumm, C.S.; Qi, D.S. Individual and combined cytotoxic effects of aflatoxin B1, zearalenone, deoxynivalenol and fumonisin B1 on BRL 3A rat liver cells. Toxicon 2015, 95, 6–12. [Google Scholar] [CrossRef] [PubMed]
- Zhou, C.; Zhang, Y.; Yin, S.; Jia, Z.; Shan, A. Biochemical changes and oxidative stress induced by zearalenone in the liver of pregnant rats. Hum. Exp. Toxicol. 2015, 34, 65–73. [Google Scholar] [CrossRef] [PubMed]
- Zourgui, L.; El Golli, E.; Bouaziz, C.; Bacha, H.; Hassen, W. Cactus (Opuntia ficus-indica) cladodes prevent oxidative damage induced by the mycotoxin zearalenone in Balb/C mice. Food Chem. Toxicol. 2008, 46, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Abid-Essefi, S.; Ouanes, Z.; Hassen, W.; Baudrimont, I.; Creppy, E.; Bacha, H. Cytotoxicity, inhibition of DNA and protein syntheses and oxidative damage in cultured cells exposed to zearalenone. Toxicol. In Vitro 2004, 18, 467–474. [Google Scholar] [CrossRef] [PubMed]
- Marin, D.E.; Pistol, G.C.; Neagoe, I.V.; Calin, L.; Taranu, I. Effects of zearalenone on oxidative stress and inflammation in weanling piglets. Food Chem. Toxicol. 2013, 58, 408–415. [Google Scholar] [CrossRef] [PubMed]
- Hassen, W.; Ayed-Boussema, I.; Oscoz, A.A.; Lopez Ade, C.; Bacha, H. The role of oxidative stress in zearalenone-mediated toxicity in Hep G2 cells: Oxidative DNA damage, gluthatione depletion and stress proteins induction. Toxicology 2007, 232, 294–302. [Google Scholar] [CrossRef] [PubMed]
- Hou, Y.J.; Zhao, Y.Y.; Xiong, B.; Cui, X.S.; Kim, N.H.; Xu, Y.X.; Sun, S.C. Mycotoxin-containing diet causes oxidative stress in the mouse. PLoS ONE 2013, 8, e60374. [Google Scholar]
- Boeira, S.P.; Funck, V.R.; Borges Filho, C.; Del’Fabbro, L.; de Gomes, M.G.; Donato, F.; Royes, L.F.; Oliveira, M.S.; Jesse, C.R.; Furian, A.F. Lycopene protects against acute zearalenone-induced oxidative, endocrine, inflammatory and reproductive damages in male mice. Chem. Biol. Interact. 2015, 230, 50–57. [Google Scholar] [CrossRef] [PubMed]
- El Golli-Bennour, E.; Bacha, H. Hsp70 expression as biomarkers of oxidative stress: Mycotoxins’ exploration. Toxicology 2011, 287, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Ben Salem, I.; Boussabbeh, M.; Helali, S.; Abid-Essefi, S.; Bacha, H. Protective effect of Crocin against zearalenone-induced oxidative stress in liver and kidney of Balb/c mice. Environ. Sci. Pollut. Res. 2015, 22, 19069–19076. [Google Scholar] [CrossRef] [PubMed]
- Bagchi, D.; Sen, C.K.; Ray, S.D.; Das, D.K.; Bagchi, M.; Preuss, H.G.; Vinson, J.A. Molecular mechanisms of cardioprotection by a novel grape seed proanthocyanidin extract. Mutat. Res. 2003, 21, 523–524. [Google Scholar] [CrossRef]
- Zhen, J.; Qu, Z.; Fang, H.; Fu, L.; Wu, Y.; Wang, H.; Zang, H.; Wang, W. Effects of grape seed proanthocyanidin extract on pentylenetetrazole-induced kindling and associated cognitive impairment in rats. Int. J. Mol. Med. 2014, 34, 391–398. [Google Scholar] [CrossRef] [PubMed]
- El-Ashmawy, I.M.; Saleh, A.; Salama, O.M. Effects of marjoram volatile oil and grape seed extract on ethanol toxicity in male rats. Basic. Clin. Pharmacol. Toxicol. 2007, 101, 320–327. [Google Scholar] [CrossRef] [PubMed]
- Feng, Y.Z.; Liu, Y.M.; Fratkins, J.D.; Leblan, M.H. Grape seed extract suppresses lipid peroxidation and reduces hypoxic ischemic brain injury in neonatal rats. Brain Res. Bull. 2005, 66, 120–127. [Google Scholar] [CrossRef] [PubMed]
- Petersen, K.S.; Smith, C. Ageing-associated oxidative stress and inflammation are alleviated by products from grapes. Oxid. Med. Cell. Longev. 2016, 2016, 6236309. [Google Scholar] [CrossRef] [PubMed]
- Sano, T.; Oda, E.; Yamashita, T.; Naemura, A.; Ijiri, Y.; Yamakoshi, J.; Yamamoto, J. Anti-thrombotic effectof proanthocyanidin, a purified ingredient of grape seed. Thromb. Res. 2005, 115, 115–121. [Google Scholar] [CrossRef] [PubMed]
- Engelbrecht, A.M.; Mattheyse, M.; Ellis, B.; Loos, B.; Thomas, M.; Smith, R.; Peters, S.; Smith, C.; Myburgh, K. Proanthocyanidin from grape seeds inactivates the PI3- kinase/PKB pathway and induces apoptosis in a colon cancer cell line. Cancer Lett. 2007, 258, 144–153. [Google Scholar] [CrossRef] [PubMed]
- Mansouri, E.; Khorsandi, L.; Zare Moaiedi, M. Grape seed proanthocyanidin extract improved some of biochemical parameters and antioxidant disturbances of red blood cells in diabetic rats. Iran. J. Pharm. Res. 2015, 14, 329–334. [Google Scholar] [PubMed]
- Yıldırım, Ş.; Topaloğlu, N.; Tekin, M.; Küçük, A.; Erdem, H.; Erbaş, M.; Yıldırım, A. Protective role of Proanthocyanidin in experimental ovarian torsion. Med. J. Islam. Repub. Iran. 2015, 29, 185. [Google Scholar] [PubMed]
- Akaberi, M.; Hosseinzadeh, H. Grapes (Vitis vinifera) as a potential candidate for the therapy of the metabolic syndrome. Phytother. Res. 2016, 30, 540–556. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.; Li, Y.; Ding, Y.; Dai, X.; Ma, X.; Bao, L.; Zhang, Z.; Li, Y. Grape seed proanthocyanidin extracts prevent high glucose-induced endothelia dysfunction via PKC and NF-κB inhibition. Biosci. Biotechnol. Biochem. 2015, 79, 493–503. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.; Zhu, Y.; Liu, Z.; Gao, Z.; Li, B.; Zhang, D.; Zhang, Z.; Jiang, X.; Liu, Z.; Meng, L.; et al. Grape seed proanthocyanidin extract ameliorates diabetic bladder dysfunction via the activation of the Nrf2 pathway. PLoS ONE 2015, 10, e0126457. [Google Scholar] [CrossRef] [PubMed]
- Long, M.; Zhang, Y.; Li, P.; Yang, S.H.; Zhang, W.K.; Han, J.X.; Wang, Y.; He, J.B. Intervention of grape seed proanthocyanidin extract on the subchronic immune injury in mice induced by aflatoxin B1. Int. J. Mol. Sci. 2016, 17, 516. [Google Scholar] [CrossRef] [PubMed]
- Ishii, T.; Itoh, K.; Yamamoto, M. Roles of Nrf2 in activation of antioxidant enzyme genes via antioxidant responsive elements. Methods Enzymol. 2002, 348, 182–190. [Google Scholar] [PubMed]
- Kobayashi, M.; Yamamoto, M. Molecular mechanisms activating the Nrf2-Keap1 pathway of antioxidant gene regulation. Antioxid. Redox Signal. 2005, 7, 385–394. [Google Scholar] [CrossRef] [PubMed]
- Ku, K.J.; Liu, X.; Fang, M.; Wu, Y.N.; Gong, Z.Y. Zearalenone induces oxidative damage involving Keap1/Nrf2/HO-1 pathway in hepatic L02 cells. Mol. Cell. Toxicol. 2014, 10, 451–457. [Google Scholar]
- Liu, M.; Gao, R.; Meng, Q.; Zhang, Y.; Bi, C.; Shan, A. Toxic effects of maternal zearalenone exposure on intestinal oxidative stress, barrier function, immunological and morphological changes in rats. PLoS ONE 2014, 9, e106412. [Google Scholar] [CrossRef] [PubMed]
- Boeira, S.P.; Filho, C.B.; Del’Fabbro, L.; Roman, S.S.; Royes, L.F.; Fighera, M.R. Lycopene treatment prevents hematological, reproductive and histopathological damage induced by acute zearalenone administration in male Swiss mice. Exp. Toxicol. Pathol. 2014, 66, 179–185. [Google Scholar] [CrossRef] [PubMed]
- Abbès, S.; Ouanes, Z.; ben Salah-Abbès, J.; Houas, Z.; Oueslati, R.; Bacha, H.; Othman, O. The protective effect of hydrated sodium calcium aluminosilicate against haematological, biochemical and pathological changes induced by Zearalenone in mice. Toxicon 2006, 47, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Vaca, C.E.; Wilhelm, J.; Hartwig, A.; Harms-Ringgdahl, M. Interaction of lipid peroxidation products with DNA. A review. Mutat. Res. 1988, 195, 137–149. [Google Scholar] [CrossRef]
- Jiang, Y.M.; Wang, Y.; Tan, H.S.; Yu, T.; Fan, X.M.; Chen, P.; Zeng, H.; Huang, M.; Bi, H.C. Schisandrol B protects against acetaminophen-induced acute hepatotoxicity in mice via activation of the Nrf2/ARE signaling pathway. Acta Pharmacol. Sin. 2016, 37, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Gum, S.I.; Cho, M.K. Recent updates on acetaminophen hepatotoxicity: The role of Nrf2 in hepatoprotection. Toxicol. Res. 2013, 29, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Otterbein, L.E.; Choi, A.M.K. Heme oxygenase: Colors of defense against cellular stress. Am. J. Physiol. Lung Cell Mol. Physiol. 2000, 279, 1029–1037. [Google Scholar]
- Moffit, J.S.; Aleksunes, L.M.; Kardas, M.J.; Slitt, A.L.; Klaassen, C.D.; Manautou, J.E. Role of NAD(P)H: Quinone oxidoreductase 1 in clofibrate-mediated hepatoprotection from acetaminophen. Toxicology 2007, 230, 197–206. [Google Scholar] [CrossRef] [PubMed]
- Kwak, M.K.; Itoh, K.; Yamamoto, M.; Kensler, T.W. Enhanced expression of the transcription factor Nrf2 by cancer chemopreventive agents: Role of antioxidant response element-like sequences in the Nrf2 promoter. Mol. Cell. Biol. 2002, 22, 2883–2892. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | ALT | AST |
|---|---|---|
| Control | 210.5 ± 12.28 a | 89.27 ± 7.20 a |
| GSPE (100 mg/kg) | 204.71 ± 10.12 a | 95.38 ± 16.68 a |
| ZEN (40 mg/kg) | 288.65 ± 34.57 b | 114.19 ± 15.64 b |
| GSPE (100 mg/kg) + ZEN (40 mg/kg) | 235.60 ± 21.25 c | 97.56 ± 12.13 a |
| Group | MDA (nmol/mgprot) | SOD (U/mgprot) | GSH-Px (U/mgprot) |
|---|---|---|---|
| Control | 7.03 ± 1.29 a | 36.32 ± 3.32 a | 213.65 ± 10.12 a |
| GSPE (100 mg/kg) | 6.21 ± 0.57 a | 40.26 ± 2.21 b | 236.15 ± 22.22 b |
| ZEN (40 mg/kg) | 9.81 ± 1.55 b | 32.58 ± 3.11 c | 193.64 ± 8.63 c |
| GSPE (100 mg/kg) + ZEN (40 mg/kg) | 8.07 ± 0.89 a | 33.99 ± 0.99 a | 207.95 ± 8.59 a |
| Gene | Accession No. | Primer Sequences (5′-3′) | Product Size/bp |
|---|---|---|---|
| Nrf2 | NM_010902.3 | F: TCCTATGCGTGAATCCCAAT | 103 bp |
| R: GCGGCTTGAATGTTTGTCTT | |||
| GSH-Px | X03920.1 | F: GAAGTGCGAAGTGAATGG | 224 bp |
| R: TGTCGATGGTACGAAAGC | |||
| HO-1 | NM_010442.2 | F: GGGCTGTGAACTCTGTCCAAT | 162 bp |
| R: GGTGAGGGAACTGTGTCAGG | |||
| γ-GCS | U85414.1 | F: TGGATGATGCCAACGAGTC | 185 bp |
| R: CCTAGTGAGCAGTACCACGAATA | |||
| NQO1 | NM_008706.5 | F: TTCTGTGGCTTCCAGGTCTT | 104 bp |
| R: TCCAGACGTTTCTTCCATCC | |||
| β-actin | BC138614.1 | F: CTGTCCCTGTATGCCTCTG | 221 bp |
| R: TTGATGTCACGCACGATT |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Long, M.; Yang, S.-H.; Han, J.-X.; Li, P.; Zhang, Y.; Dong, S.; Chen, X.; Guo, J.; Wang, J.; He, J.-B. The Protective Effect of Grape-Seed Proanthocyanidin Extract on Oxidative Damage Induced by Zearalenone in Kunming Mice Liver. Int. J. Mol. Sci. 2016, 17, 808. https://doi.org/10.3390/ijms17060808
Long M, Yang S-H, Han J-X, Li P, Zhang Y, Dong S, Chen X, Guo J, Wang J, He J-B. The Protective Effect of Grape-Seed Proanthocyanidin Extract on Oxidative Damage Induced by Zearalenone in Kunming Mice Liver. International Journal of Molecular Sciences. 2016; 17(6):808. https://doi.org/10.3390/ijms17060808
Chicago/Turabian StyleLong, Miao, Shu-Hua Yang, Jian-Xin Han, Peng Li, Yi Zhang, Shuang Dong, Xinliang Chen, Jiayi Guo, Jun Wang, and Jian-Bin He. 2016. "The Protective Effect of Grape-Seed Proanthocyanidin Extract on Oxidative Damage Induced by Zearalenone in Kunming Mice Liver" International Journal of Molecular Sciences 17, no. 6: 808. https://doi.org/10.3390/ijms17060808
APA StyleLong, M., Yang, S.-H., Han, J.-X., Li, P., Zhang, Y., Dong, S., Chen, X., Guo, J., Wang, J., & He, J.-B. (2016). The Protective Effect of Grape-Seed Proanthocyanidin Extract on Oxidative Damage Induced by Zearalenone in Kunming Mice Liver. International Journal of Molecular Sciences, 17(6), 808. https://doi.org/10.3390/ijms17060808