
Mechanism of Action and Applications of Interleukin 24 in Immunotherapy

Abstract
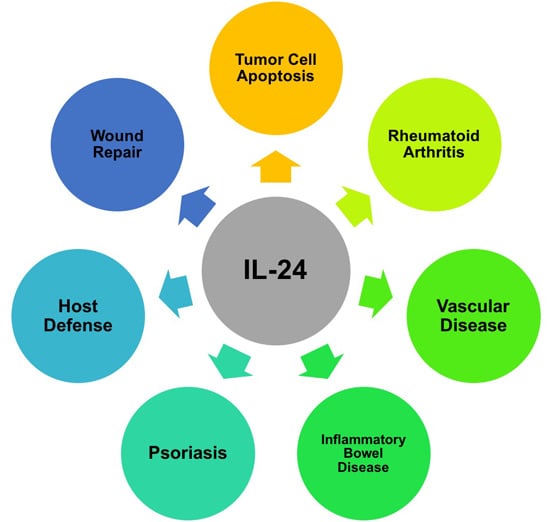
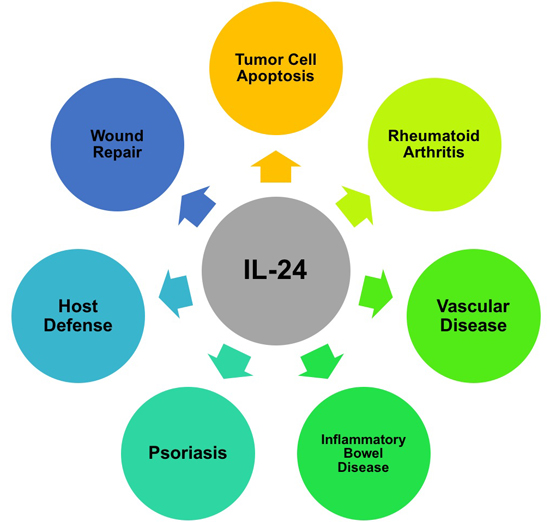
:
1. Introduction
2. The Cellular Sources of IL-24 Expression
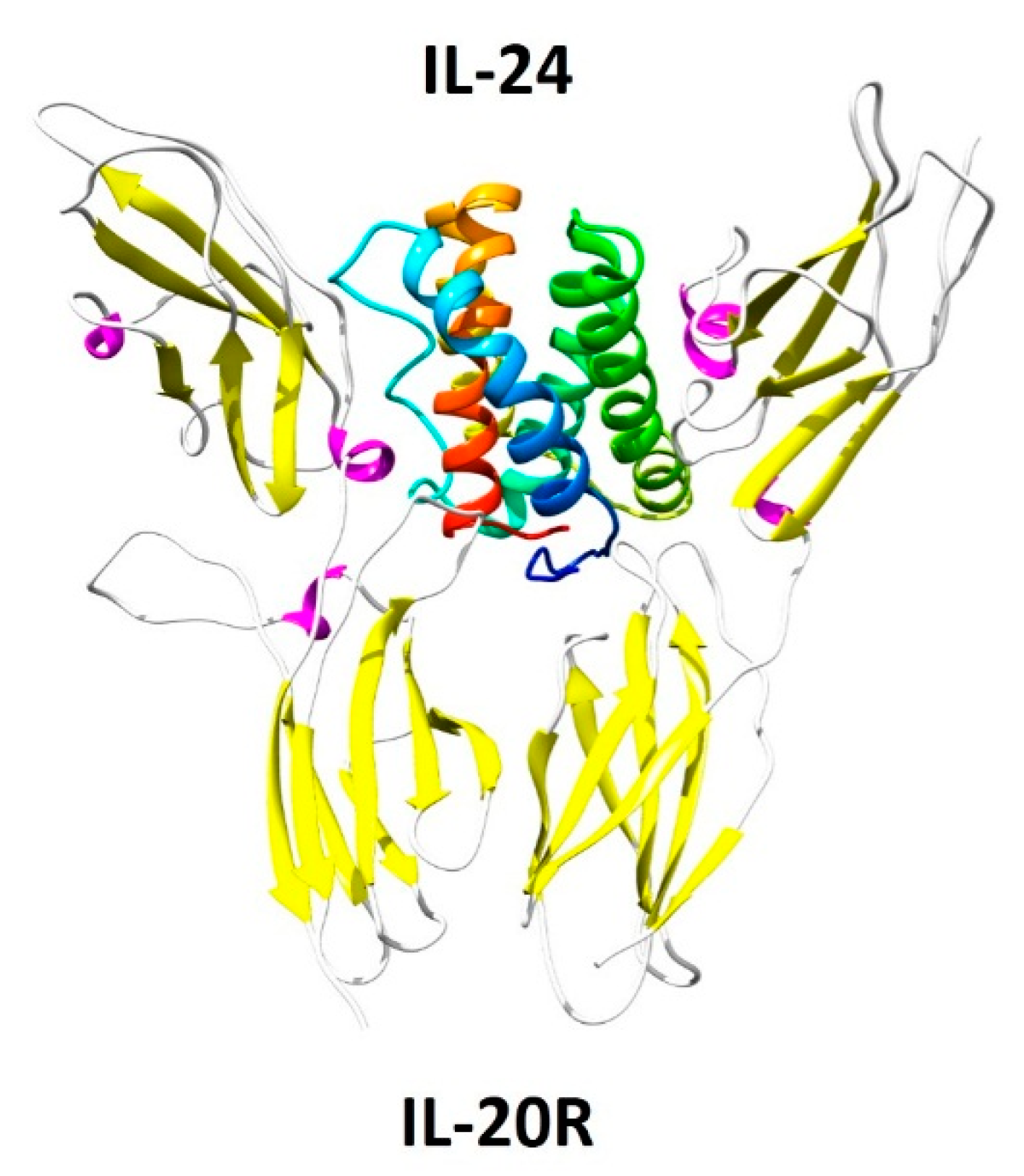
3. IL-24 and Its Receptors
4. Interleukin-24 in Cancer
5. Anti-Angiogenic Properties of IL-24
6. Metastasis
7. IL-24 and Inflammatory Diseases
7.1. Psoriasis
7.2. Rheumatoid Arthritis
7.3. Inflammatory Bowel Disease
8. IL-24 in Host Defense
9. Wound Repair
10. Cardiovascular Disease
11. Discussion
Acknowledgments
Conflicts of Interest
References
- Hofmann, S.R.; Rösen-Wolff, A.; Tsokos, G.C.; Hedrich, C.M. Biological properties and regulation of IL-10 related cytokines and their contribution to autoimmune disease and tissue injury. Clin. Immunol. 2012, 143, 116–127. [Google Scholar] [CrossRef] [PubMed]
- Andoh, A.; Shioya, M.; Nishida, A.; Bamba, S.; Tsujikawa, T.; Kim-Mitsuyama, S.; Fujiyama, Y. Expression of IL-24, an activator of the JAK1/STAT3/SOCS3 cascade, is enhanced in inflammatory bowel disease. J. Immunol. 2009, 183, 687–695. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Tan, Z.; Zhang, R.; Kotenko, S.V.; Liang, P. Interleukin 24 (MDA-7/MOB-5) signals through two heterodimeric receptors, IL-22R1/IL-20R2 and IL-20R1/IL-20R2. J. Biol. Chem. 2002, 277, 7341–7347. [Google Scholar] [CrossRef] [PubMed]
- Buzas, K.; Oppenheim, J.J.; Zack Howard, O.M. Myeloid cells migrate in response to IL-24. Cytokine 2011, 55, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Wahl, C.; Müller, W.; Leithäuser, F.; Adler, G.; Oswald, F.; Reimann, J.; Schirmbeck, R.; Seier, A.; Weiss, J.M.; Prochnow, B.; et al. IL-20 receptor 2 signaling downregulates antigen-specific T cell responses. J. Immunol. 2009, 182, 802–810. [Google Scholar] [CrossRef] [PubMed]
- Maarof, G.; Bouchet-Delbos, L.; Gary-Gouy, H.; Durand-Gasselin, I.; Krzysiek, R.; Dalloul, A. Interleukin-24 inhibits the plasma cell differentiation program in human germinal center B cells. Blood 2010, 115, 1718–1726. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Lin, J.J.; Su, Z.Z.; Goldstein, N.I.; Fisher, P.B. Subtraction hybridization identifies a novel melanoma differentiation associated gene, mda-7, modulated during human melanoma differentiation, growth and progression. Oncogene 1995, 11, 2477–2486. [Google Scholar] [PubMed]
- Parrish-Novak, J.; Xu, W.; Brender, T.; Yao, L.; Jones, C.; West, J.; Brandt, C.; Jelinek, L.; Madden, K.; McKernan, P.A.; et al. Interleukins 19, 20, and 24 signal through two distinct receptor complexes. Differences in receptor-ligand interactions mediate unique biological functions. J. Biol. Chem. 2002, 277, 47517–47523. [Google Scholar] [CrossRef] [PubMed]
- Sauane, M.; Gopalkrishnan, R.V.; Lebedeva, I.; Mei, M.X.; Sarkar, D.; Su, Z.Z.; Kang, D.C.; Dent, P.; Pestka, S.; Fisher, P.B. Mda-7/IL-24 induces apoptosis of diverse cancer cell lines through JAK/STAT-independent pathways. J. Cell. Physiol. 2003, 196, 334–345. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Zhang, D.; Gao, Z.; Li, H.; Zhang, B.; Zhang, Q.; Li, L.; Cheng, Q.; Pei, D.; Zheng, J. MDA-7/IL-24 inhibits Nrf2-mediated antioxidant response through activation of p38 pathway and inhibition of ERK pathway involved in cancer cell apoptosis. Cancer Gene Ther. 2014, 21, 416–426. [Google Scholar] [CrossRef] [PubMed]
- Dudakov, J.A.; Hanash, A.M.; van den Brink, M.R. Interleukin-22: Immunobiology and pathology. Annu. Rev. Immunol. 2015, 33, 747–785. [Google Scholar] [CrossRef] [PubMed]
- Sabat, R.; Wallace, E.; Endesfelder, S.; Wolk, K. IL-19 and IL-20: Two novel cytokines with importance in inflammatory diseases. Exp. Opin. Ther. Targets 2007, 11, 601–612. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.; Yi, W.; Chen, Y.; Ma, L.; Wang, J.; Yu, G. Overexpression of Sig1R is closely associated with tumor progression and poor outcome in patients with hilar cholangiocarcinoma. Med. Oncol. 2014, 31. [Google Scholar] [CrossRef] [PubMed]
- Do, W.; Herrera, C.; Mighty, J.; Shumskaya, M.; Redenti, S.M.; Sauane, M. Sigma 1 Receptor plays a prominent role in IL-24-induced cancer-specific apoptosis. Biochem. Biophys. Res. Commun. 2013, 439, 215–220. [Google Scholar] [CrossRef] [PubMed]
- Su, T.P.; Su, T.C.; Nakamura, Y.; Tsai, S.Y. The Sigma-1 receptor as a pluripotent modulator in living systems. Trends Pharmacol. Sci. 2016, 37, 262–278. [Google Scholar] [CrossRef] [PubMed]
- Palmer, C.P.; Mahen, R.; Schnell, E.; Djamgoz, M.B.; Aydar, E. Sigma-1 receptors bind cholesterol and remodel lipid rafts in breast cancer cell lines. Cancer Res. 2007, 67, 11166–11175. [Google Scholar] [CrossRef] [PubMed]
- Ortega-Roldan, J.L.; Ossa, F.; Schnell, J.R. Characterization of the human sigma-1 receptor chaperone domain structure and binding immunoglobulin protein (BiP) interactions. J. Biol. Chem. 2013, 288, 21448–21457. [Google Scholar] [CrossRef] [PubMed]
- Cunningham, C.C.; Chada, S.; Merritt, J.A.; Tong, A.; Senzer, N.; Zhang, Y.; Mhashilkar, A.; Parker, K.; Vukelja, S.; Richards, D.; et al. Clinical and local biological effects of an intratumoral injection of mda-7 (IL24; INGN 241) in patients with advanced carcinoma: A phase I study. Mol. Ther. 2005, 11, 149–159. [Google Scholar] [CrossRef] [PubMed]
- Tong, A.W.; Nemunaitis, J.; Su, D.; Zhang, Y.; Cunningham, C.; Senzer, N.; Netto, G.; Rich, D.; Mhashilkar, A.; Parker, K.; et al. Intratumoral injection of INGN 241, a nonreplicating adenovector expressing the melanoma-differentiation associated gene-7 (mda-7/IL24): Biologic outcome in advanced cancer patients. Mol. Ther. 2005, 11, 160–172. [Google Scholar] [CrossRef] [PubMed]
- Sauane, M.; Lebedeva, I.; Su, Z.; Choo, H.; Randolph, A.; Valerie, K.; Dent, P.; Gopalkrishnan, R.V.; Fisher, P.B. Melanoma differentiation associated gene-7/interleukin-24 promotes tumor cell-specific apoptosis through both secretory and nonsecretory pathways. Cancer Res. 2004, 64. [Google Scholar] [CrossRef]
- Sauane, M.; Su, Z.; Dash, R.; Liu, X.; Norris, J.S.; Sarkar, D.; Lee, S.; Allegood, J.C.; Dent, P.; Spiegel, S.; et al. Ceramide plays a prominent role in MDA-7/IL-24-induced cancer-specific apoptosis. J. Cell. Physiol. 2009, 222, 546–555. [Google Scholar] [CrossRef] [PubMed]
- Sauane, M.; Su, Z.; Gupta, P.; Lebedeva, I.; Dent, P.; Sarkar, D.; Fisher, P.B. Autocrine regulation of mda-7/IL-24 mediates cancer-specific apoptosis. Proc. Natl. Acad. Sci. USA 2008, 105, 9763–9768. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Su, Z.; Lebedeva, I.; Sauane, M.; Gopalkrishnan, R.V.; Valerie, K.; Dent, P.; Fisher, P.B. mda-7 (IL-24) mediates selective apoptosis in human melanoma cells by inducing the coordinated overexpression of the GADD family of genes by means of p38 MAPK. Proc. Natl. Acad. Sci. USA 2002, 99, 10054–10059. [Google Scholar] [CrossRef] [PubMed]
- Otkjaer, K.; Holtmann, H.; Kragstrup, T.W.; Paludan, S.R.; Johansen, C.; Gaestel, M.; Kragballe, K.; Iversen, L. The p38 MAPK Regulates IL-24 Expression by Stabilization of the 3′ UTR of IL-24 mRNA. PLoS ONE 2010, 5, e8671. [Google Scholar] [CrossRef] [PubMed]
- Sahoo, A.; Yun, M.J.; Kwon, H.K.; Yi, H.J.; Lee, S.; Chang, S.; Park, Z.Y.; Hwang, K.C.; Im, S.H. A novel splicing variant of mouse interleukin (IL)-24 antagonizes IL-24-induced apoptosis. J. Biol. Chem. 2008, 283, 28860–28872. [Google Scholar] [CrossRef] [PubMed]
- Whitaker, E.L.; Filippov, V.; Filippova, M.; Guerrero-Juarez, C.F.; Duerksen-Hughes, P.J. Splice variants of mda-7/IL-24 differentially affect survival and induce apoptosis in U2OS cells. Cytokine 2011, 56, 272–281. [Google Scholar] [CrossRef] [PubMed]
- Filippov, V.; Schmidt, E.L.; Filippova, M.; Duerksen-Hughes, P.J. Splicing and splice factor SRp55 participate in the response to DNA damage by changing isoform ratios of target genes. Gene 2008, 420, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Caudell, E.G.; Mumm, J.B.; Poindexter, N.; Ekmekcioglu, S.; Mhashilkar, A.M.; Yang, X.H.; Retter, M.W.; Hill, P.; Chada, S.; Grimm, E.A. The protein product of the tumor suppressor gene, melanoma differentiation-associated gene 7, exhibits immunostimulatory activity and is designated IL-24. J. Immunol. 2002, 168, 6041–6046. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, R.; Mhashilkar, A.M.; Tanaka, F.; Saito, Y.; Branch, C.D.; Sieger, K.; Mumm, J.B.; Stewart, A.L.; Boquio, A.; Dumoutier, L.; et al. Melanoma differentiation-associated gene 7/interleukin (IL)-24 is a novel ligand that regulates angiogenesis via the IL-22 receptor. Cancer Res. 2003, 63, 5105–5113. [Google Scholar] [PubMed]
- Oncology Drugs in the Pipeline. HemOnc Today, 2015. Available online: http://m1.wyanokecdn.com/154294cceb75a0f83401b139bc08edf7.pdf (accessed on 8 May 2016).
- Chiocca, E.A.; Rabkin, S.D. Oncolytic viruses and their application to cancer immunotherapy oncolytic viruses. Cancer Immunol. Res. 2012, 2, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Gu, J.; Dong, A.; Zhang, Y.; Zhong, L.; He, L.; Wang, Y.; Zhang, J.; Zhang, Z.; Huiwang, J.; et al. Potent antitumor activity of oncolytic adenovirus expressing mda-7/IL-24 for colorectal cancer. Hum. Gene Ther. 2005, 16, 845–858. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Dong, A.; Gu, J.; Liu, Z.; Zhang, Y.; Zhang, W.; Wang, Y.; He, L.; Qian, C.; Qian, Q.; et al. The antitumor activity of TRAIL and IL-24 with replicating oncolytic adenovirus in colorectal cancer. Cancer Gene Ther. 2006, 13, 1011–1022. [Google Scholar] [CrossRef] [PubMed]
- Luo, J.; Xia, Q.; Zhang, R.; Lv, C.; Zhang, W.; Wang, Y.; Cui, Q.; Liu, L.; Cai, R.; Qian, C. Treatment of cancer with a novel dual-targeted conditionally replicative adenovirus armed with mda-7/IL-24 gene. Clin. Cancer Res. 2008, 14, 2450–2457. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.J.; Zhang, J.; Wu, Y.M.; Qiang, J.; Liu, X.J.; Yan, L.C.; Zhou, X.M.; Xiao, R.J.; Wang, Y.G.; Cao, X.; et al. Complete eradication of hepatomas using an oncolytic adenovirus containing AFP promoter controlling E1A and an E1B deletion to drive IL-24 expression. Cancer Gene Ther. 2012, 19, 619–629. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.; Jiang, A.J.; Cheng, Q.; Tian, H.; Li, L.T.; Zheng, J.N. A dual-regulated oncolytic adenovirus expressing interleukin-24 sensitizes melanoma cells to temozolomide via the induction of apoptosis. Tumor Biol. 2013, 34, 1263–1271. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.J.; Wang, Y.G.; Cao, X.; Zhong, S.Y.; Wei, R.C.; Wu, Y.M.; Yue, X.T.; Li, G.C.; Liu, X.Y. Potent antitumor effect of interleukin-24 gene in the survivin promoter and retinoblastoma double-regulated oncolytic adenovirus. Hum. Gene Ther. 2009, 20, 818–830. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Liang, P. IL-24 transgenic mice: In vivo evidence of overlapping functions for IL-20, IL-22, and IL-24 in the epidermis. J. Immunol. 2010, 184, 1793–1798. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Huang, C.; Kato-Maeda, M.; Hopewell, P.C.; Daley, C.L.; Krensky, A.M.; Clayberger, C. Messenger RNA expression of IL-8, FOXP3, and IL-12 differentiates latent tuberculosis infection from disease. J. Immunol. 2007, 178, 3688–3694. [Google Scholar] [CrossRef] [PubMed]
- Xiao, L.; Li, X.; Niu, N.; Qian, J.; Xie, G.; Wang, Y. Dichloroacetate (DCA) enhances tumor cell death in combination with oncolytic adenovirus armed with MDA-7/IL-24. Mol. Cell. Biochem. 2010, 340, 31–40. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.; Xu, L.; Yuan, H.; Zhang, Y.; Zhang, X.; Zhao, D. Oncolytic adenovirus-mediated mda-7/IL-24 expression suppresses osteosarcoma growth and enhances sensitivity to doxorubicin. Mol. Med. Rep. 2015, 12, 6358–6364. [Google Scholar] [CrossRef] [PubMed]
- Lou, W.; Chen, Q.; Ma, L.; Liu, J.; Yang, Z.; Shen, J.; Cui, Y.; Bian, X.W.; Qian, C. Oncolytic adenovirus co-expressing miRNA-34a and IL-24 induces superior antitumor activity in experimental tumor model. J. Mol. Med. 2013, 91, 715–725. [Google Scholar] [CrossRef] [PubMed]
- Chada, S.; Sutton, R.B.; Ekmekcioglu, S.; Ellerhorst, J.; Mumm, J.B.; Leitner, W.W.; Yang, H.Y.; Sahin, A.A.; Hunt, K.K.; Fuson, K.L.; et al. MDA-7/IL-24 is a unique cytokine-tumor suppressor in the IL-10 Family. Int. Immunopharmacol. 2004, 4, 649–667. [Google Scholar] [CrossRef] [PubMed]
- Gopalan, B.; Litvak, A.; Sharma, S.; Mhashilkar, A.M.; Chada, S. Activation of the Fas-FasL signaling pathway by MDA-7/IL-24 kills human ovarian cancer cells. Cancer Res. 2005, 65, 3017–3024. [Google Scholar] [PubMed]
- Das, D.; Persaud, L.; Dejoie, J.; Happy, M.; Brannigan, O.; de Jesus, D.; Sauane, M. Tumor necrosis factor-related apoptosis-inducing ligand (TRAIL) activates caspases in human prostate cancer cells through sigma 1 receptor. Biochem. Biophys. Res. Commun. 2016, 470, 1–5. [Google Scholar] [CrossRef] [PubMed]
- Ramesh, R.; Ito, I.; Saito, Y.; Mhashilkar, A.; Branch, C.D.; Chada, S. Overexpression of the melanoma differentiation associated-7 (mda-7)/interleukin-24 (IL-24) gene impairs lung cancer cell migration by modulating matrix metalloproteinase-2 (MMP-2) and E-cadherin expression. Mol. Ther. 2004, 9, 510–518. [Google Scholar] [CrossRef] [PubMed]
- Chada, S.; Bocangel, D.; Ramesh, R.; Grimm, E.A.; Mumm, J.B.; Mhashilkar, A.M.; Zheng, M. mda-7/IL24 kills pancreatic cancer cells by inhibition of the Wnt/PI3K signaling pathways: Identification of IL-20 receptor-mediated bystander activity against pancreatic cancer. Mol. Ther. 2005, 11, 724–733. [Google Scholar] [CrossRef] [PubMed]
- Akashi, T.; Koizumi, K.; Tsuneyama, K.; Saiki, I.; Takano, Y.; Fuse, H. Chemokine receptor CXCR4 expression and prognosis in patients with metastatic prostate cancer. Cancer Sci. 2008, 99, 539–542. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, J.; Munshi, A.; Ramesh, R. Molecular targets and signaling pathways regulated by interleukin (IL)-24 in mediating its antitumor activities. J. Mol. Signal. 2013, 8, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Panneerselvam, J.; Jin, J.; Shanker, M.; Lauderdale, J.; Bates, J.; Wang, Q.; Zhao, Y.D.; Archibald, S.J.; Hubin, T.J.; Ramesh, R. IL-24 Inhibits lung cancer cell migration and invasion by disrupting the SDF-1/CXCR4 signaling axis. PLoS ONE 2015, 10, e0122439. [Google Scholar] [CrossRef] [PubMed]
- Bech, R.; Otkjaer, K.; Birkelund, S.; Vorup-Jensen, T.; Agger, R.; Johansen, C.; Iversen, L.; Kragballe, K.; Rømer, J. Interleukin 20 protein locates to distinct mononuclear cells in psoriatic skin. Exp. Dermatol. 2014, 23, 349–352. [Google Scholar] [CrossRef] [PubMed]
- Kumari, S.; Bonnet, M.C.; Ulvmar, M.H.; Wolk, K.; Karagianni, N.; Witte, E.; Uthoff-Hachenberg, C.; Renauld, J.C.; Kollias, G.; Toftgard, R.; et al. Tumor necrosis factor receptor signaling in keratinocytes triggers interleukin-24-dependent psoriasis-like skin inflammation in mice. Immunity 2013, 39, 899–911. [Google Scholar] [CrossRef] [PubMed]
- Sa, S.M.; Valdez, P.A.; Wu, J.; Jung, K.; Zhong, F.; Hall, L.; Kasman, I.; Winer, J.; Modrusan, Z.; Danilenko, D.M.; et al. The effects of IL-20 subfamily cytokines on reconstituted human epidermis suggest potential roles in cutaneous innate defense and pathogenic adaptive immunity in psoriasis. J. Immunol. 2007, 178, 2229–2240. [Google Scholar] [CrossRef] [PubMed]
- Kragstrup, T.W.; Otkjaer, K.; Holm, C.; Jørgensen, A.; Hokland, M.; Iversen, L.; Deleuran, B. The expression of IL-20 and IL-24 and their shared receptors are increased in rheumatoid arthritis and spondyloarthropathy. Cytokine 2008, 41, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Fonseca-Camarillo, G.; Furuzawa-Carballeda, J.; Granados, J.; Yamamoto-Furusho, J.K. Expression of interleukin (IL)-19 and IL-24 in inflammatory bowel disease patients: A cross-sectional study. Clin. Exp. Immunol. 2014, 177, 64–75. [Google Scholar] [CrossRef] [PubMed]
- Tamai, H.; Miyake, K.; Yamaguchi, H.; Takatori, M.; Dan, K.; Inokuchi, K.; Shimada, T. AAV8 vector expressing IL24 efficiently suppresses tumor growth mediated by specific mechanisms in MLL/AF4-positive ALL model mice. Blood 2012, 119, 64–71. [Google Scholar] [CrossRef] [PubMed]
- Jin, S.H.; Choi, D.; Chun, Y.J.; Noh, M. Keratinocyte-derived IL-24 plays a role in the positive feedback regulation of epidermal inflammation in response to environmental and endogenous toxic stressors. Toxicol. Appl. Pharmacol. 2014, 280, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Ma, Y.; Chen, H.; Wang, Q.; Luo, F.; Yan, J.; Zhang, X.L. IL-24 protects against Salmonella typhimurium infection by stimulating early neutrophil Th1 cytokine production, which in turn activates CD8+ T cells. Eur. J. Immunol. 2009, 39, 3357–3368. [Google Scholar] [CrossRef] [PubMed]
- Bastonero, S.; Le Priol, Y.; Armand, M.; Bernard, C.S.; Reynaud-Gaubert, M.; Olive, D.; Parzy, D.; de Bentzmann, S.; Capo, C.; Mege, J.-L. New microbicidal functions of tracheal glands: Defective anti-infectious response to Pseudomonas aeruginosa in cystic fibrosis. PLoS ONE 2009, 4, e5357. [Google Scholar] [CrossRef] [PubMed]
- Wu, B.; Huang, C.; Kato-Maeda, M.; Hopewell, P.C.; Daley, C.L.; Krensky, A.M.; Clayberger, C. IL-24 modulates IFN-γ expression in patients with tuberculosis. Immunol. Lett. 2008, 117, 57–62. [Google Scholar] [CrossRef] [PubMed]
- Buzas, K.; Megyeri, K. Staphylococci induce the production of melanoma differentiation-associated protein-7/IL-24. Acta Microbiol. Immunol. Hung. 2006, 53, 431–440. [Google Scholar] [CrossRef] [PubMed]
- Miller, L.S.; Cho, J.S. Immunity against Staphylococcus aureus cutaneous infections. Nat. Rev. Immunol. 2011, 11, 505–518. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.S.; Pietras, E.M.; Garcia, N.C.; Ramos, R.I.; Farzam, D.M.; Monroe, H.R.; Magorien, J.E.; Blauvelt, A.; Kolls, J.K.; Cheung, A.L.; et al. IL-17 is essential for host defense against cutaneous Staphylococcus aureus infection in mice. J. Clin. Investig. 2010, 120, 1762–1773. [Google Scholar] [CrossRef] [PubMed]
- Myles, I.A.; Fontecilla, N.M.; Valdez, P.A.; Vithayathil, P.J.; Naik, S.; Belkaid, Y.; Ouyang, W.; Datta, S.K. Signaling via the IL-20 receptor inhibits cutaneous production of IL-1β and IL-17A to promote infection with methicillin-resistant Staphylococcus aureus. Nat. Immunol. 2013, 14, 804–811. [Google Scholar] [CrossRef] [PubMed]
- Bosanquet, D.C.; Harding, K.G.; Ruge, F.; Sanders, A.J.; Jiang, W.G. Expression of IL-24 and IL-24 receptors in human wound tissues and the biological implications of IL-24 on keratinocytes. Wound Repair Regen. 2012, 20, 896–903. [Google Scholar] [CrossRef] [PubMed]
- Poindexter, N.J.; Williams, R.R.; Powis, G.; Jen, E.; Caudle, A.S.; Chada, S.; Grimm, E.A. IL-24 is expressed during wound repair and inhibits TGFα-induced migration and proliferation of keratinocytes. Exp. Dermatol. 2010, 19, 714–722. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.; Sanders, A.J.; Zhang, Y.; Martin, T.A.; Owen, S.; Ruge, F.; Jiang, W.G. Interleukin-24 (IL-24) expression and biological impact on HECV endothelial cells. Cancer Genom. Proteom. 2015, 12, 243–250. [Google Scholar]
- Lee, K.M.; Kang, H.A.; Park, M.; Lee, H.Y.; Choi, H.R.; Yun, C.H.; Oh, J.W.; Kang, H.S. Interleukin-24 attenuates β-glycerophosphate-induced calcification of vascular smooth muscle cells by inhibiting apoptosis, the expression of calcification and osteoblastic markers, and the Wnt/β-catenin pathway. Biochem. Biophys. Res. Commun. 2012, 428, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Chada, S.; Mhashilkar, A.; Miano, J.M. Tumor suppressor MDA-7/IL-24 selectively inhibits vascular smooth muscle cell growth and migration. Mol. Ther. 2003, 8, 220–229. [Google Scholar] [CrossRef]
- Lee, K.M.; Kang, H.A.; Ko, C.B.; Oh, E.H.; Park, M.; Lee, H.Y.; Choi, H.R.; Yun, C.H.; Jung, W.W.; Oh, J.W.; et al. Differential gene expression profiles in spontaneously hypertensive rats induced by administration of enalapril and nifedipine. Int. J. Mol. Med. 2013, 31, 179–187. [Google Scholar] [PubMed]
- Vargas-Alarcón, G.; Posadas-Romero, C.; Villarreal-Molina, T.; Alvarez-León, E.; Angeles-Martinez, J.; Posadas-Sanchez, R.; Monroy-Muñoz, I.; Luna-Fuentes, S.; González-Salazar, C.; Ramirez-Bello, J.; et al. IL-24 gene polymorphisms are associated with cardiometabolic parameters and cardiovascular risk factors but not with premature coronary artery disease: The genetics of atherosclerotic disease Mexican study. J. Interferon Cytokine Res. 2014, 34, 659–666. [Google Scholar] [CrossRef] [PubMed]
- Gupta, P.; Walter, M.R.; Su, Z.Z.; Lebedeva, I.V.; Emdad, L.; Randolph, A.; Valerie, K.; Sarkar, D.; Fisher, P.B. BiP/GRP78 is an intracellular target for MDA-7/IL-24 induction of cancer-specific apoptosis. Cancer Res. 2006, 66, 8182–8191. [Google Scholar] [CrossRef] [PubMed]
- Pataer, A.; Vorburger, S.A.; Chada, S.; Balachandran, S.; Barber, G.N.; Roth, J.A.; Hunt, K.K.; Swisher, S.G. Melanoma differentiation-associated gene-7 protein physically associates with the double-stranded RNA-activated protein kinase PKR. Mol. Ther. 2005, 11, 717–723. [Google Scholar] [CrossRef] [PubMed]
- Cruz, A.; Nguyen, B.; Sauane, M.; Lopez, G.E. Structural and functional characterization of interleukin-24 based on atomistic molecular modeling. Chem. Lett. 2016, 45, 327–329. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
| Inducers of Expression | Cellular Sources | Transcription Factors | Receptor |
|---|---|---|---|
| IL-1β | Monocytes | Jak1 | IL-20R1/IL-20R2 |
| IL-17 | Melanocytes | Tyrosine kinase 2 | IL-22R1/IL-20R2 |
| IL-22 | TH2 lymphocytes | Stat 1 | - |
| LPS | Fibroblasts Keratinocytes | Stat 3 | - |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Persaud, L.; De Jesus, D.; Brannigan, O.; Richiez-Paredes, M.; Huaman, J.; Alvarado, G.; Riker, L.; Mendez, G.; Dejoie, J.; Sauane, M. Mechanism of Action and Applications of Interleukin 24 in Immunotherapy. Int. J. Mol. Sci. 2016, 17, 869. https://doi.org/10.3390/ijms17060869
Persaud L, De Jesus D, Brannigan O, Richiez-Paredes M, Huaman J, Alvarado G, Riker L, Mendez G, Dejoie J, Sauane M. Mechanism of Action and Applications of Interleukin 24 in Immunotherapy. International Journal of Molecular Sciences. 2016; 17(6):869. https://doi.org/10.3390/ijms17060869
Chicago/Turabian StylePersaud, Leah, Dayenny De Jesus, Oliver Brannigan, Maria Richiez-Paredes, Jeannette Huaman, Giselle Alvarado, Linda Riker, Gissete Mendez, Jordan Dejoie, and Moira Sauane. 2016. "Mechanism of Action and Applications of Interleukin 24 in Immunotherapy" International Journal of Molecular Sciences 17, no. 6: 869. https://doi.org/10.3390/ijms17060869