
Anti-Cancer Effect of Lambertianic Acid by Inhibiting the AR in LNCaP Cells


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
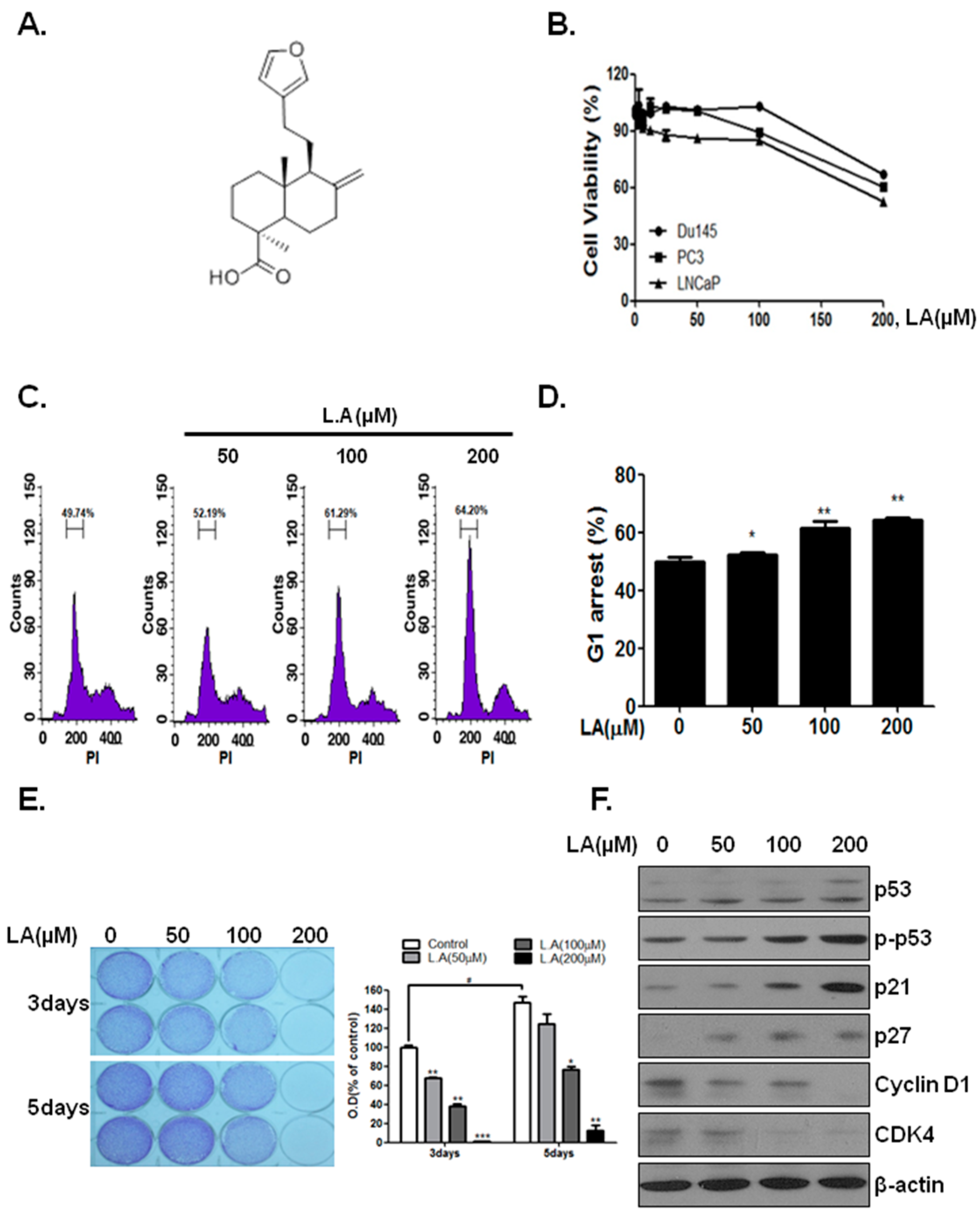
2.1. Lambertianic Acid Inhibits Cell Growt
2.2. Lambertianic Acid Induces the Apoptosis of LNCaP Cells
2.3. Lambertianic Acid Attenuates AR and PSA Expression in LNCaP Cells
2.4. Lambertianic Acid Inhibits Androgen-Stimulated AR Nuclear Translocation
2.5. AR Signaling Mediates Lambertianic Acid-Induced Cell Proliferation Suppression and Apoptosis
3. Discussion
4. Materials and Methods
4.1. Test Compound
4.2. Cell Culture Assay
4.3. Cell Viability Assay
4.4. Western Blot Analysis
4.5. Crystal Violet Staining
4.6. Cell Cycle Analysis
4.7. AR Gene Silencing
4.8. Luciferase Assay
4.9. Caspase-3 Activity
4.10. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| LA | lambertianic acid |
| AR | androgen receptor |
| PBS | phosphate-buffered saline |
| OD | optical density |
References
- Ferlay, J.; Soerjomataram, I.; Dikshit, R.; Eser, S.; Mathers, C.; Rebelo, M.; Parkin, D.M.; Forman, D.; Bray, F. Cancer incidence and mortality worldwide: Sources, methods and major patterns in globocan. Int. J. Cancer 2012, 136, E359–E386. [Google Scholar] [CrossRef] [PubMed]
- Siegel, R.; Ma, J.; Zou, Z.; Jemal, A. Cancer statistics. CA Cancer J. Clin. 2014, 64, 9–29. [Google Scholar] [CrossRef] [PubMed]
- Bennett, N.C.; Gardiner, R.A.; Hooper, J.D.; Johnson, D.W.; Gobe, G.C. Molecular cell biology of androgen receptor signalling. Int. J. Biochem. Cell Biol. 2010, 42, 813–827. [Google Scholar] [CrossRef] [PubMed]
- Andriole, G.; Bruchovsky, N.; Chung, L.W.; Matsumoto, A.M.; Rittmaster, R.; Roehrborn, C.; Russell, D.; Tindall, D. Dihydrotestosterone and the prostate: The scientific rationale for 5alpha-reductase inhibitors in the treatment of benign prostatic hyperplasia. J. Urol. 2004, 172, 1399–1403. [Google Scholar] [CrossRef] [PubMed]
- Feitelson, M.A.; Arzumanyan, A.; Kulathinal, R.J.; Blain, S.W.; Holcombe, R.F.; Mahajna, J.; Marino, M.; Martinez-Chantar, M.L.; Nawroth, R.; Sanchez-Garcia, I.; et al. Sustained proliferation in cancer: Mechanisms and novel therapeutic targets. Semin. Cancer Biol. 2015, 35, S25–S54. [Google Scholar] [CrossRef] [PubMed]
- Heinlein, C.A.; Chang, C. Androgen receptor in prostate cancer. Endocr. Rev. 2004, 25, 276–308. [Google Scholar] [CrossRef] [PubMed]
- Taplin, M.E.; Balk, S.P. Androgen receptor: A key molecule in the progression of prostate cancer to hormone independence. J. Cell. Biochem. 2004, 91, 483–490. [Google Scholar] [CrossRef] [PubMed]
- Cleutjens, K.B.; van der Korput, H.A.; van Eekelen, C.C.; van Rooij, H.C.; Faber, P.W.; Trapman, J. An androgen response element in a far upstream enhancer region is essential for high, androgen-regulated activity of the prostate-specific antigen promoter. Mol. Endocrinol. 1997, 11, 148–161. [Google Scholar] [CrossRef] [PubMed]
- Cleutjens, K.B.; van Eekelen, C.C.; van der Korput, H.A.; Brinkmann, A.O.; Trapman, J. Two androgen response regions cooperate in steroid hormone regulated activity of the prostate-specific antigen promoter. J. Biol. Chem. 1996, 271, 6379–6388. [Google Scholar] [PubMed]
- Riegman, P.H.; Vlietstra, R.J.; van der Korput, J.A.; Romijn, J.C.; Trapman, J. Characterization of the prostate-specific antigen gene: A novel human kallikrein-like gene. Biochem. Biophys. Res. Commun. 1989, 159, 95–102. [Google Scholar] [CrossRef]
- Yu, H.M.; Wu, Y.; Ju, P.; Wang, B.H.; Yang, X.D.; Wang, H.M.; Xu, L.C. Enos-jnk1-ar signaling pathway mediates deltamethrin-induced germ cells apoptosis in testes of adult rats. Environ. Toxicol. Pharmacol. 2014, 38, 733–741. [Google Scholar] [CrossRef] [PubMed]
- Tong, K.L.; Chan, K.L.; AbuBakar, S.; Low, B.S.; Ma, H.Q.; Wong, P.F. The in vitro and in vivo anti-cancer activities of a standardized quassinoids composition from eurycoma longifolia on lncap human prostate cancer cells. PLoS ONE 2015, 10, e0121752. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Liu, G.; Tong, D.; Parmar, H.; Hasenmayer, D.; Yuan, W.; Zhang, D.; Jiang, J. Metformin represses androgen-dependent and androgen-independent prostate cancers by targeting androgen receptor. Prostate 2015, 75, 1187–1196. [Google Scholar] [CrossRef] [PubMed]
- Shenk, J.L.; Fisher, C.J.; Chen, S.Y.; Zhou, X.F.; Tillman, K.; Shemshedini, L. P53 represses androgen-induced transactivation of prostate-specific antigen by disrupting har amino- to carboxyl-terminal interaction. J. Biol. Chem. 2001, 276, 38472–38479. [Google Scholar] [CrossRef] [PubMed]
- Won, S.H.; Lee, H.J.; Jeong, S.J.; Lu, J.; Kim, S.H. Activation of p53 signaling and inhibition of androgen receptor mediate tanshinone iia induced g1 arrest in lncap prostate cancer cells. Phytother. Res. 2012, 26, 669–674. [Google Scholar] [CrossRef] [PubMed]
- Bruckheimer, E.M.; Spurgers, K.; Weigel, N.L.; Logothetis, C.; McDonnell, T.J. Regulation of bcl-2 expression by dihydrotestosterone in hormone sensitive lncap-fgc prostate cancer cells. J. Urol. 2003, 169, 1553–1557. [Google Scholar] [CrossRef] [PubMed]
- Yang, X.; Zhang, Y.C.; Zhang, H.; Wang, J. Isolation and identification of diterpenoids from Pinus koraiensis. J. Chin. Med. Mater. 2008, 31, 53–55. [Google Scholar]
- Chae, H.S.; Khiev, P.; Lee, H.K.; Oh, S.R.; Chin, Y.W. Anti-allergic effect of a chloroform-soluble extract of cinnamomum cambodianum in bone marrow-derived mast cells. Immunopharmacol. Immunotoxicol. 2012, 34, 639–644. [Google Scholar] [CrossRef] [PubMed]
- Tolstikova, T.G.; Voevoda, T.V.; Dolgikh, M.P.; Sorokina, I.V. Neurotropic activity of lambertian acid and its amino derivatives. Eksp. Klin. Farmakol. 2002, 65, 9–11. [Google Scholar] [PubMed]
- Yang, X.; Ding, Y.; Sun, Z.H.; Zhang, D.M. Studies on chemical constituents of pinus armandii. Acta Pharm. Sin. 2005, 40, 435–437. [Google Scholar]
- Chae, H.S.; Chin, Y.W. Anti-allergic effect of Lambertianic acid from thuja orientalis in mouse bone marrow-derived mast cells. Immunopharmacol. Immunotoxicol. 2012, 34, 250–255. [Google Scholar] [CrossRef] [PubMed]
- Lee, M.S.; Cho, S.M.; Lee, M.H.; Lee, E.O.; Kim, S.H.; Lee, H.J. Ethanol extract of Pinus koraiensis leaves containing Lambertianic acid exerts anti-obesity and hypolipidemic effects by activating adenosine monophosphate-activated protein kinase (ampk). BMC Complement. Altern. Med. 2016, 16, 51. [Google Scholar] [CrossRef] [PubMed]
- Balk, S.P.; Ko, Y.J.; Bubley, G.J. Biology of prostate-specific antigen. J. Clin. Oncol. 2003, 21, 383–391. [Google Scholar] [CrossRef] [PubMed]
- Pezaro, C.; Woo, H.H.; Davis, I.D. Prostate cancer: Measuring psa. Intern. Med. J. 2014, 44, 433–440. [Google Scholar] [CrossRef] [PubMed]
- Knudsen, K.E.; Arden, K.C.; Cavenee, W.K. Multiple g1 regulatory elements control the androgen-dependent proliferation of prostatic carcinoma cells. J. Biol. Chem. 1998, 273, 20213–20222. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Lee, H.J.; Wang, L.; Jiang, C.; Baek, N.I.; Kim, S.H.; Lu, J. Anti-androgen receptor signaling and prostate cancer inhibitory effects of sucrose- and benzophenone-compounds. Pharm. Res. 2009, 26, 1140–1148. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Jiang, C.; Wang, Z.; Lee, H.J.; Hu, H.; Malewicz, B.; Lee, H.J.; Lee, J.H.; Baek, N.I.; Jeong, J.H.; et al. A novel class of pyranocoumarin anti-androgen receptor signaling compounds. Mol. Cancer Ther. 2007, 6, 907–917. [Google Scholar] [CrossRef] [PubMed]
- Kokontis, J.M.; Lin, H.P.; Jiang, S.S.; Lin, C.Y.; Fukuchi, J.; Hiipakka, R.A.; Chung, C.J.; Chan, T.M.; Liao, S.; Chang, C.H.; et al. Androgen suppresses the proliferation of androgen receptor-positive castration-resistant prostate cancer cells via inhibition of cdk2, cyclina, and skp2. PLoS ONE 2014, 9, e109170. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.M.; Sicinski, P. Targeting cyclins and cyclin-dependent kinases in cancer: Lessons from mice, hopes for therapeutic applications in human. Cell Cycle 2006, 5, 2110–2114. [Google Scholar] [CrossRef] [PubMed]
- Liao, X.; Tang, S.; Thrasher, J.B.; Griebling, T.L.; Li, B. Small-interfering rna-induced androgen receptor silencing leads to apoptotic cell death in prostate cancer. Mol. Cancer Ther. 2005, 4, 505–515. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Altuwaijri, S.; Deng, F.; Chen, L.; Lal, P.; Bhanot, U.K.; Korets, R.; Wenske, S.; Lilja, H.G.; Chang, C.; et al. Nf-kappab regulates androgen receptor expression and prostate cancer growth. Am. J. Pathol. 2009, 175, 489–499. [Google Scholar] [CrossRef] [PubMed]
- Pelekanou, V.; Notas, G.; Stathopoulos, E.N.; Castanas, E.; Kampa, M. Androgen receptors in early and castration resistant prostate cancer: Friend or foe? Hormones 2013, 12, 224–235. [Google Scholar] [CrossRef] [PubMed]





© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lee, M.-S.; Lee, S.-O.; Kim, S.-H.; Lee, E.-O.; Lee, H.-J. Anti-Cancer Effect of Lambertianic Acid by Inhibiting the AR in LNCaP Cells. Int. J. Mol. Sci. 2016, 17, 1066. https://doi.org/10.3390/ijms17071066
Lee M-S, Lee S-O, Kim S-H, Lee E-O, Lee H-J. Anti-Cancer Effect of Lambertianic Acid by Inhibiting the AR in LNCaP Cells. International Journal of Molecular Sciences. 2016; 17(7):1066. https://doi.org/10.3390/ijms17071066
Chicago/Turabian StyleLee, Myoung-Sun, Seon-Ok Lee, Sung-Hoon Kim, Eun-Ok Lee, and Hyo-Jeong Lee. 2016. "Anti-Cancer Effect of Lambertianic Acid by Inhibiting the AR in LNCaP Cells" International Journal of Molecular Sciences 17, no. 7: 1066. https://doi.org/10.3390/ijms17071066