
Expression Analysis of PIN Genes in Root Tips and Nodules of Medicago truncatula



Abstract
:1. Introduction
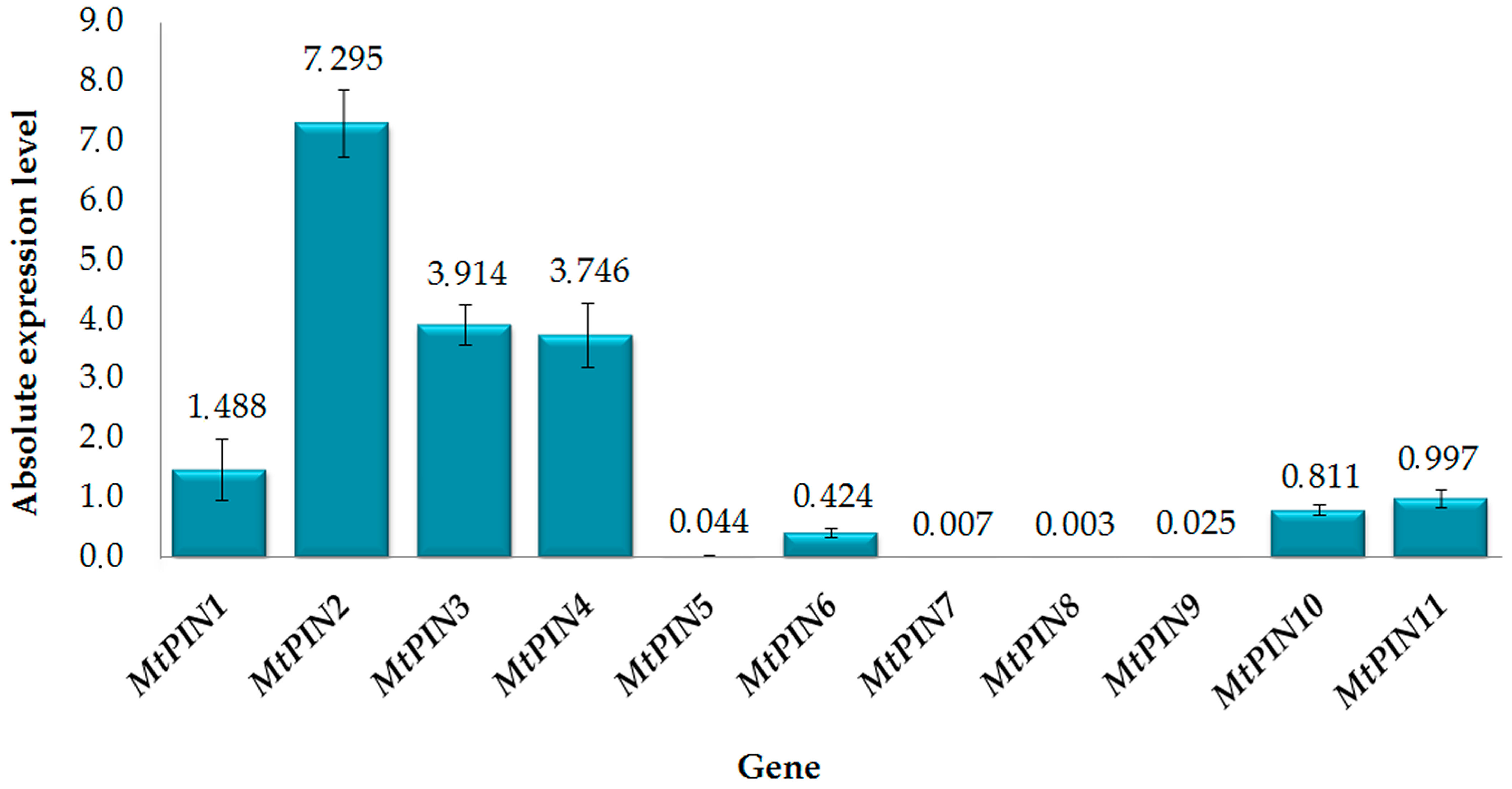
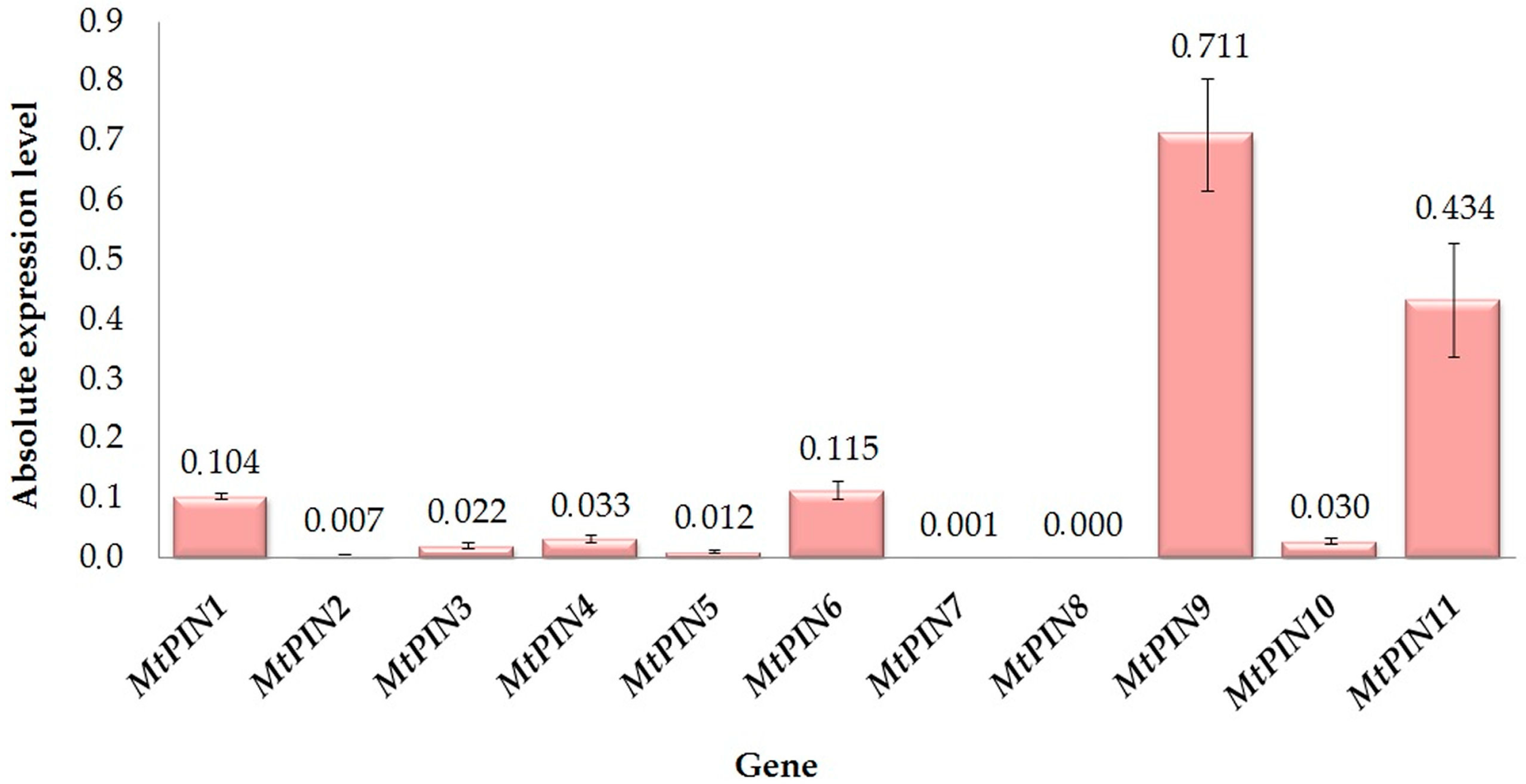
2. Results
3. Discussion
4. Experimental Section
4.1. Plant Material, Inoculation with Rhizobia and Growth Conditions
4.2. RNA Isolation and cDNA Synthesis
4.3. Real-Time Quantitative PCR
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ATP | adenosine triphosphate |
| AUX1 | AUXIN1 |
| BLAST | basic local alignment search tool |
| bp | base pair |
| CDS | coding DNA sequence |
| cv. | cultivar |
| ER | endoplasmic reticulum |
| IAA− | anionic indole-3-acetic acid |
| IAAH | protonated form of IAA |
| LAX | LIKE AUX1 |
| NCBI | national center for biotechnology information |
| PAT | polar auxin transport |
| PCR | polymerase chain reaction |
| PGP | P-glycoprotein |
| qPCR | quantitative polymerase chain reaction |
| RAM | root apical meristem |
| SE | standard error |
References
- Sprent, J.I.; Sprent, P. Nitrogen Fixing Organisms: Pure and Applied Aspects; Chapman and Hall: London, UK, 1990. [Google Scholar]
- Perret, X.; Staehelin, C.; Broughton, W.J. Molecular basis of symbiotic promiscuity. Microbiol. Mol. Biol. Rev. 2000, 64, 180–201. [Google Scholar] [CrossRef] [PubMed]
- Esseling, J.J.; Lhuissier, F.G.P.; Emons, A.M.C. Nod factor-induced root hair curling: Continuous polar growth towards the point of nod factor application. Plant Physiol. 2003, 132, 1982–1988. [Google Scholar] [CrossRef] [PubMed]
- Brewin, N.J. Plant cell wall remodelling in the rhizobium-legume symbiosis. Crit. Rev. Plant Sci. 2004, 23, 293–316. [Google Scholar] [CrossRef]
- Barnett, M.J.; Fisher, R.F. Global gene expression in the rhizobial-legume symbiosis. Symbiosis 2006, 42, 1–24. [Google Scholar]
- Poole, P.; Allaway, D. Carbon and nitrogen metabolism in Rhizobium. Adv. Microb. Physiol. 2000, 43, 117–163. [Google Scholar] [PubMed]
- Ott, T.; van Dongen, J.T.; Guenther, C.; Krusell, L.; Desbrosses, G.; Vigeolas, H.; Bock, V.; Czechowski, T.; Geigenberger, P.; Udvardi, M.K. Symbiotic leghemoglobins are crucial for nitrogen fixation in legume root nodules but not for general plant growth and development. Curr. Biol. 2005, 15, 531–535. [Google Scholar] [CrossRef] [PubMed]
- Barsch, A.; Carvalho, H.G.; Cullimore, J.V.; Niehaus, K. GC-MS based metabolite profiling implies three interdependent ways of ammonium assimilation in Medicago truncatula root nodules. J. Biotechnol. 2006, 127, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Vasse, J.; de Billy, F.; Camut, S.; Truchet, G. Correlation between ultrastructural differentiation of bacteroids and nitrogen fixation in alfalfa nodules. J. Bacteriol. 1990, 172, 4295–4306. [Google Scholar] [PubMed]
- Timmers, A.C.J.; Soupéne, E.; Auriac, M.C.; de Billy, F.; Vasse, J.; Boistard, P.; Truchet, G. Saprophytic intracellular rhizobia in alfalfa nodules. Mol. Plant Microbe Interact. 2000, 13, 1204–1213. [Google Scholar] [CrossRef] [PubMed]
- Vieten, A.; Sauer, M.; Brewer, P.B.; Friml, J. Molecular and cellular aspects of auxin-transport-mediated development. Trends Plant Sci. 2007, 12, 160–168. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Yang, H.; Peer, W.A.; Richter, G.; Blakeslee, J.J.; Bhandyopadhyay, A.; Titapiwatanakun, B.; Undurraga, S.; Khodakovskaya, M.; Richards, E.L.; et al. Arabidopsis H+-PPase AVP1 regulates auxin-mediated organ development. Science 2005, 310, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Michniewicz, M.; Brewer, P.B.; Friml, J. Polar auxin transport and asymmetric auxin distribution. In The Arabidopsis Book; Last, R., Ed.; American Society of Plant Biologists: Rockville, MD, USA, 2007; pp. 1–28. [Google Scholar]
- Mravec, J.; Skůpa, P.; Bailly, A.; Hoyerová, K.; Křeček, P.; Bielach, A.; Petrasek, J.; Zhang, J.; Gaykova, V.; Stierhof, Y.-D.; et al. Subcellular homeostasis of phytohormone auxin is mediated by the ER-localized PIN5 transporter. Nature 2009, 459, 1136–1140. [Google Scholar] [CrossRef] [PubMed]
- Ding, Z.; Wang, B.; Moreno, I.; Dupláková, N.; Simon, S.; Carraro, N.; Reemmer, J.; Pěnčik, A.; Chen, X.; Tejos, R.; et al. ER-localized auxin transporter PIN8 regulates auxin homeostasis and male gametophyte development in Arabidopsis. Nat. Commun. 2012, 3, 941. [Google Scholar] [CrossRef] [PubMed]
- Sawchuk, M.G.; Edgar, A.; Scarpella, E. Patterning of leaf vein networks by convergent auxin transport pathways. PLoS Genet. 2013, 9, e1003294. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Jones, A.R. Endoplasmic reticulum: The rising compartment in auxin biology. Plant Physiol. 2010, 154, 458–462. [Google Scholar] [CrossRef] [PubMed]
- Schnabel, E.L.; Frugoli, J. The PIN and LAX families of auxin transport genes in Medicago truncatula. Mol. Genet. Genom. 2004, 272, 420–432. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Ge, L.; Wang, Y.; Chen, R. Signaling and transport of auxin and plant development. In Polar Auxin Transport; Chen, R., Baluška, F., Eds.; Springer: Berlin & Heidelberg, Germany, 2013; pp. 239–258. [Google Scholar]
- Hirsch, A.M.; Bhuvaneswari, T.V.; Torrey, J.G.; Bisseling, T. Early nodulin genes are induced in alfalfa root outgrowths elicited by auxin transport inhibitors. Proc. Natl. Acad. Sci. USA 1989, 86, 1244–1248. [Google Scholar] [CrossRef] [PubMed]
- Huo, X.; Schnabel, E.; Hughes, K.; Frugoli, J. RNAi phenotypes and the localization of a protein: GUS fusion imply a role for Medicago truncatula PIN genes in nodulation. J. Plant Growth Regul. 2006, 25, 156–165. [Google Scholar] [CrossRef] [PubMed]
- Wasson, A.P.; Pellerone, F.I.; Mathesius, U. Silencing the flavonoid pathway in Medicago truncatula inhibits root nodule formation and prevents auxin transport regulation by rhizobia. Plant Cell 2006, 18, 1617–1629. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Yue, R.; Bai, Y.; Feng, R.; Sun, T.; Wang, X.; Yang, Y.; Tie, S.; Wang, H. Identification and analysis of Medicago truncatula auxin transporter gene families uncover their roles in responses to Sinorhizobium meliloti infection. Plant Cell Physiol. 2015, 56, 1930–1943. [Google Scholar] [CrossRef] [PubMed]
- Bustos-Sanmamed, P.; Mao, G.; Deng, Y.; Elouet, M.; Khan, G.A.; Bazin, J.; Turner, M.; Subramanian, S.; Yu, O.; Crespi, M.; et al. Overexpression of miR160 affects root growth and nitrogen-fixing nodule number in Medicago truncatula. Funct. Plant Biol. 2013, 40, 1208–1220. [Google Scholar] [CrossRef]
- Breakspear, A.; Liu, C.; Roy, S.; Stacey, N.; Rogers, C.; Trick, M.; Morieri, G.; Mysore, K.S.; Wen, J.; Oldroyd, G.E.D.; et al. The root hair “infectome” of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for Auxin signaling in rhizobial infection. Plant Cell 2014, 26, 4680–4701. [Google Scholar] [CrossRef] [PubMed]
- Ramakers, C.; Ruijter, J.M.; Deprez, R.H.; Moorman, A.F. Assumption-free analysis of quantitative real-time polymerase chain reaction (PCR) data. Neurosci. Lett. 2003, 339, 62–66. [Google Scholar] [CrossRef]
- Pfaffl, M.W.; Horgan, G.W.; Dempfle, L. Relative expression software tool (REST) for group-wise comparison and statistical analysis of relative expression results in real-time PCR. Nucleic Acids Res. 2002, 30, e36. [Google Scholar] [CrossRef] [PubMed]
- Blilou, I.; Xu, J.; Wildwater, M.; Willemsen, V.; Paponov, I.; Friml, J.; Heidstra, R.; Aida, M.; Palme, K.; Scheres, B. The PIN auxin efflux facilitator network controls growth and patterning in Arabidopsis roots. Nature 2005, 433, 39–44. [Google Scholar] [CrossRef] [PubMed]
- Holmes, P.; Goffard, N.; Weiller, G.F.; Rolfe, B.G.; Imin, N. Transcriptional profiling of Medicago truncatula meristematic root cells. BMC Plant Biol. 2008, 8, 1. [Google Scholar] [CrossRef] [PubMed]
- Müller, A.; Guan, C.; Gälweiler, L.; Tänzler, P.; Huijser, P.; Marchant, A.; Parry, G.; Bennett, M.; Wisman, E.; Palme, K. AtPIN2 defines a locus of Arabidopsis for root gravitropism control. EMBO J. 1998, 17, 6903–6911. [Google Scholar] [CrossRef] [PubMed]
- Rahman, A.; Takahashi, M.; Shibasaki, K.; Wu, S.; Inaba, T.; Tsurumi, S.; Baskin, T.I. Gravitropism of Arabidopsis thaliana roots requires the polarization of PIN2 toward the root tip in meristematic cortical cells. Plant Cell 2010, 22, 1762–1776. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Wiśniewska, J.; Benková, E.; Mendgen, K.; Palme, K. Lateral relocation of auxin efflux regulator PIN3 mediates tropism in Arabidopsis. Nature 2002, 415, 806–809. [Google Scholar] [CrossRef] [PubMed]
- Friml, J.; Benková, E.; Blilou, I.; Wisniewska, J.; Hamann, T.; Ljung, K.; Woody, S.; Sandberg, G.; Scheres, B.; Jürgens, G.; et al. AtPIN4 mediates sink-driven auxin gradients and root patterning in Arabidopsis. Cell 2002, 108, 661–673. [Google Scholar] [CrossRef]
- Gälweiler, L.; Guan, C.; Müller, A.; Wisman, E.; Mendgen, K.; Yephremov, A.; Palme, K. Regulation of polar auxin transport by AtPIN1 in Arabidopsis vascular tissue. Science 1998, 282, 2226–2230. [Google Scholar] [PubMed]
- Woodward, A.W.; Bartel, B. Auxin: Regulation, action, and interaction. Ann. Bot. 2005, 95, 707–735. [Google Scholar] [CrossRef] [PubMed]
- Fedorova, E.E.; Redondo, F.J.; Koshiba, T.; Pueyo, J.J.; de Felipe, M.R.; Lucas, M.M. Aldehyde oxidase (AO) in the root nodules of Lupinus albus and Medicago truncatula: Identification of AO in meristematic and infection zones. Mol. Plant Microbe Interact. 2005, 18, 405–413. [Google Scholar] [CrossRef] [PubMed]
- Rightmyer, A.P.; Long, S.R. Pseudonodule formation by wild-type and asymbiotic mutant Medicago truncatula in response to auxin transport inhibitors. Mol. Plant Microbe Interact. 2011, 24, 1372–1384. [Google Scholar] [CrossRef] [PubMed]
- Mathesius, U.; Bayliss, C.; Weinman, J.; Schlaman, H.R.M.; Spaink, H.P.; Rolfe, B.G.; McCully, M.E.; Djordjevic, M.A. Flavonoids synthesized in cortical cells during nodule initiation are early developmental markers in white clover. Mol. Plant Microbe Interact. 1998, 11, 1223–1232. [Google Scholar] [CrossRef]
- Prayitno, J.; Rolfe, B.G.; Mathesius, U. The ethylene-insensitive sickle mutant of Medicago truncatula shows altered auxin transport regulation during nodulation. Plant Physiol. 2006, 142, 168–180. [Google Scholar] [CrossRef] [PubMed]
- De Billy, F.; Grosjean, C.; May, S.; Bennett, M.; Cullimore, J.V. Expression studies on AUX1-like genes in Medicago truncatula suggest that auxin is required at two steps in early nodule development. Mol. Plant Microbe Interact. 2001, 14, 267–277. [Google Scholar] [CrossRef] [PubMed]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef] [PubMed]
- Bianco, C.; Senatore, B.; Arbucci, S.; Pieraccini, G.; Defez, R. Modulation of endogenous indole-3-acetic acid biosynthesis in bacteroids within Medicago sativa nodules. Appl. Environ. Microbiol. 2014, 80, 4286–4293. [Google Scholar] [CrossRef] [PubMed]
- Vincent, J.M. A Manual for the Practical Study of Root Nodule Bacteria; Blackwell Scientific Publications Ltd.: Oxford, UK, 1970. [Google Scholar]
- Untergasser, A.; Nijveen, H.; Rao, X.; Bisseling, T.; Geurts, R.; Leunissen, J.A.M. Primer3Plus, an enhanced web interface to Primer3. Nucleic Acids Res. 2007, 35, W71–W74. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| M. truncatula Gene/Protein | A. thaliana Orthologous Gene according to Schnabel and Frugoli and Peng et al. | A. thaliana Orthologous Coding DNA Sequences (CDS) Identified Using BLAST Search | A. thaliana Orthologous Protein Sequences Identified Using BLAST Search |
|---|---|---|---|
| MtPIN1 | AtPIN3 | AtPIN4 | AtPIN4 |
| AtPIN4 | |||
| AtPIN7 | |||
| MtPIN2 | AtPIN2 | AtPIN2 | AtPIN2 |
| MtPIN3 | AtPIN3 | AtPIN3 | AtPIN3 |
| AtPIN4 | |||
| AtPIN7 | |||
| MtPIN4 | AtPIN1 | AtPIN1 | AtPIN1 |
| MtPIN5 | AtPIN1 | AtPIN1 | AtPIN1 |
| MtPIN6 | AtPIN6 | AtPIN6 | AtPIN6 |
| MtPIN7 | AtPIN2 | AtPIN2 | AtPIN7 |
| AtPIN7 | |||
| MtPIN8 | AtPIN8 | AtPIN8 | AtPIN8 |
| MtPIN9 | AtPIN5 | AtPIN5 | AtPIN5 |
| MtPIN10 | AtPIN1 | AtPIN1 | AtPIN1 |
| MtPIN11 | AtPIN8 | AtPIN8 | AtPIN8 |
| Gene | Type | Reaction Efficiency | Expression | Standard Error | 95% Confidence Interval | p-Value | Result |
|---|---|---|---|---|---|---|---|
| MtRPS7b | Reference gene | 0.8951 | 1.000 | ||||
| MtPIN1 | Target gene | 0.7752 | 0.107 | 0.090–0.123 | 0.085–0.137 | 0.001 | DOWN |
| MtPIN2 | Target gene | 0.9265 | 0.001 | 0.001–0.002 | 0.001–0.002 | 0.000 | DOWN |
| MtPIN3 | Target gene | 0.9236 | 0.007 | 0.006–0.011 | 0.004–0.012 | 0.000 | DOWN |
| MtPIN4 | Target gene | 0.9509 | 0.010 | 0.007–0.017 | 0.004–0.023 | 0.000 | DOWN |
| MtPIN5 | Target gene | 0.9748 | 0.288 | 0.194–0.445 | 0.117–0.576 | 0.000 | DOWN |
| MtPIN6 | Target gene | 0.9849 | 0.235 | 0.151–0.351 | 0.105–0.440 | 0.000 | DOWN |
| MtPIN7 | Target gene | 0.8672 | 0.146 | 0.098–0.215 | 0.079–0.300 | 0.001 | DOWN |
| MtPIN8 | Target gene | 0.7412 | 0.012 | 0.009–0.016 | 0.007–0.019 | 0.002 | DOWN |
| MtPIN9 | Target gene | 1.0000 | 29.32 | 19.258–40.578 | 15.769–55.179 | 0.000 | UP |
| MtPIN10 | Target gene | 0.9001 | 0.044 | 0.033–0.055 | 0.023–0.090 | 0.002 | DOWN |
| MtPIN11 | Target gene | 0.9686 | 0.445 | 0.263–0.658 | 0.179–1.515 | 0.007 | DOWN |
| Temperature | Time |
|---|---|
| PCR | |
| 50 °C | 20 s |
| 95 °C | 10 min |
| 40 Cycles: | |
| 95 °C | 15 s |
| 60 °C | 1 min |
| Melting curve | |
| 95 °C | 15 s |
| 60 °C | 1 min |
| 95 °C | 30 s |
| 60 °C | 15 s |
| Gene/ID | Forward (F) and Reverse (R) Primer Sequence | Sequence of Each MtPIN First Exon Fragment | Product Length |
|---|---|---|---|
| 5′–3′ | (bp) | ||
| MtPIN1 (MTR_4g084870) | F: TCCACTTTACGTAGCCATGATCT | ATGATAACCTGGCACGATCTATACACAGTTTTAACCGCAGTAGTTAGCCTATGGCTCCGTACGGTGGTGGAAAATCCGGCATAAACCGTTTCGTCGC | 74 |
| R: AACATTGGTCCGGTGAGAAT | |||
| MtPIN2 (MTR_4g127100) | F: CGAAGATGAGACATTGAGGATG | ATGATTACCGGTAAGGATATATACGATGTTTTCGCA...CATAAGAAAAGGGGAGGGAGGAGTATGAGGTTCTTACCCTCCTCCAAATCCTATGC | 74 |
| R: CACCATTATTGAACAACTCACCA | |||
| MtPIN3 (MTR_1g030890) | F: CTGGCCTCAACGTGTTCC | ATGATAACACTAAAAGATCTTTACACTGTCTTAACAGCAGTGGTTCCA...GAAATTCGGAACAATCGGAAGAGGGTGCTAGCTGATGAACATAATCAAAA | 68 |
| R: CACCACCATCCTGATCTCCT | |||
| MtPIN4 (MTR_6g069510) | F: TGGTGCCACTTTATGTAGCTATG | ATGATCACTTTAACAGATTTCTACCATGTCATGACAGCAAATCTTAGCTTATGGATCAGTAAAATGGTGGAAAATATTTTCACCTGATTTTGTTGCA | 92 |
| R: ACGGTTGATTCCTGAACATTG | |||
| MtPIN5 (MTR_8g107360) | F: CGTGGCTATGATATTAGCTTATGG | ATGATAACGTTAACAGATTTCTACCATGTGATGACATCAATGGTGCCACTTTATTCAGTGAAATGGTGGAAGATACGGCATCAATCGCTTCGTTGCT | 66 |
| R: GAGCATTGATCGGGAGAGAA | |||
| MtPIN6 (MTR_1g029190) | F: TAAACCGATTCGTCGCAGTT | ATGGTGACAAGAGAAGATTTATACAACGTGATGTGTGCAATGGTACCTC...TTTGCTGTTCCAGTTCTATCTTTTTATCAAATGGACACAAAATTTAT | 67 |
| R: GGATTGTTGAGAGAAATGAAGTGA | |||
| MtPIN7 (MTR_4g127090) | F: TTGTGCCACTATATGTCGCTATG | ATGATTACCGGCAAGGACATATACAATGTTTTAGCGGCGAATATTAGCATATGGTTCGGTCCGATGGTGGAAAATCTTCACACCAGATTGTCTC | 94 |
| R: AAACGGTTTATTCCAGAACATTG | |||
| MtPIN8 (MTR_7g009370) | F: TTTCCTTAGCCAATGTTTATCATGT | ATGAAATAACAACAACTGTCCCATTATATGTAACAATGATACTAGCCTTCACACAAGAACAATGTTCAGGAATAAACAAATTTGTTGC | 95 |
| R: GATCTTAAACCATTTCACTGAGACATA | |||
| MtPIN9 (MTR_7g079720) | F: AGCAGTGGTGCCACTCTATTTT | ATGATTGGGTGGGAAGACGTGTACAAAGTTATTGTGCACTAATATTAGGCTATGGTTCTGTAAGGTGGTGGAAAATTTTCACAAACTAGTTT | 96 |
| R: TTGTTTATTGCATCACATTGTTCTC | |||
| MtPIN10 (MTR_7g089360) | F: TGGTGTTGCTAAAGCTAATGGA | ATGATAAGTGCTTTAGACTTATACCATGTCCTCACAGCAGTAGTACC...AATGGTGGAAATGGCTACCCATTTTTTCACCTGTGGCTAATAAGAAAAA | 61 |
| R: CCCTGCACTATGAGGAGCA | |||
| MtPIN11 (MTR_6g011400) | F: ACAGCCACTGTCCCATTATATGT | ATGATTTCCTTAATTGATGTCTATCATGTAGTAAACTATGTTACTAGCATACATTTGTGTTAAATGGTGGAAACTTTGGCATAAACAAATTTGTAGC | 87 |
| R: TGCACATTGATCTGGTGTGA | |||
| MtRPS7b (MTR_8g087480) | F: GAAACAACACTGCAATTTACAGGA | ATGACCTTACCATACCCATACCATCACCATTGTTGT...AACTATCAGGCAAAGATGTTGTCTTTAGTTCTGTTTCTCAATTTTGATTTTGTTTCATG | 74 |
| R: CCTAAGCCTCAGTAACGGGATA |
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sańko-Sawczenko, I.; Łotocka, B.; Czarnocka, W. Expression Analysis of PIN Genes in Root Tips and Nodules of Medicago truncatula. Int. J. Mol. Sci. 2016, 17, 1197. https://doi.org/10.3390/ijms17081197
Sańko-Sawczenko I, Łotocka B, Czarnocka W. Expression Analysis of PIN Genes in Root Tips and Nodules of Medicago truncatula. International Journal of Molecular Sciences. 2016; 17(8):1197. https://doi.org/10.3390/ijms17081197
Chicago/Turabian StyleSańko-Sawczenko, Izabela, Barbara Łotocka, and Weronika Czarnocka. 2016. "Expression Analysis of PIN Genes in Root Tips and Nodules of Medicago truncatula" International Journal of Molecular Sciences 17, no. 8: 1197. https://doi.org/10.3390/ijms17081197
APA StyleSańko-Sawczenko, I., Łotocka, B., & Czarnocka, W. (2016). Expression Analysis of PIN Genes in Root Tips and Nodules of Medicago truncatula. International Journal of Molecular Sciences, 17(8), 1197. https://doi.org/10.3390/ijms17081197