
Plastoglobule-Targeting Competence of a Putative Transit Peptide Sequence from Rice Phytoene Synthase 2 in Plastids

Abstract
:
1. Introduction
2. Results
2.1. Bioinformatic Prediction of Putative Plastoglobule-Targeting Sequences from OsPSY1, 2, and 3
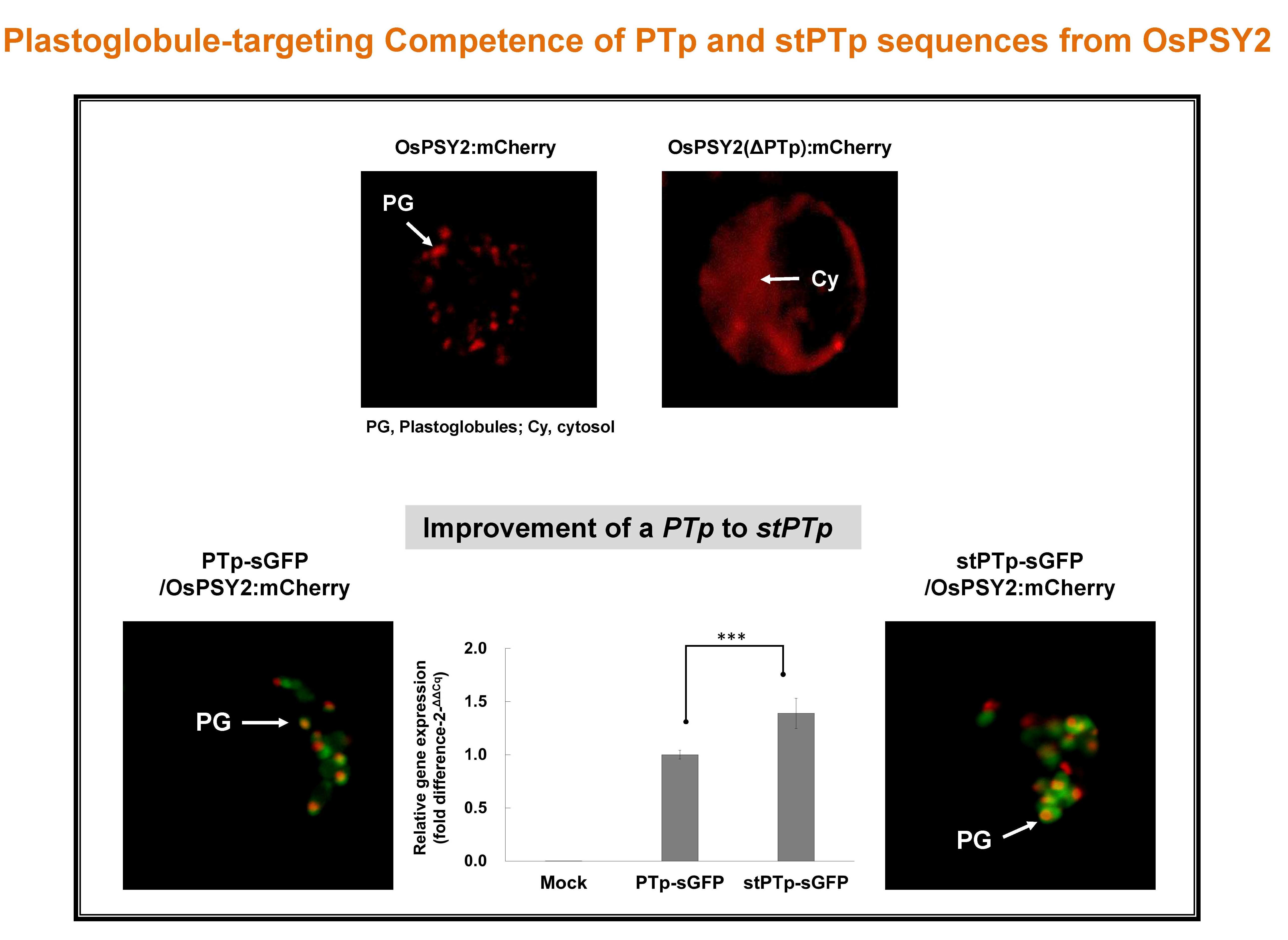
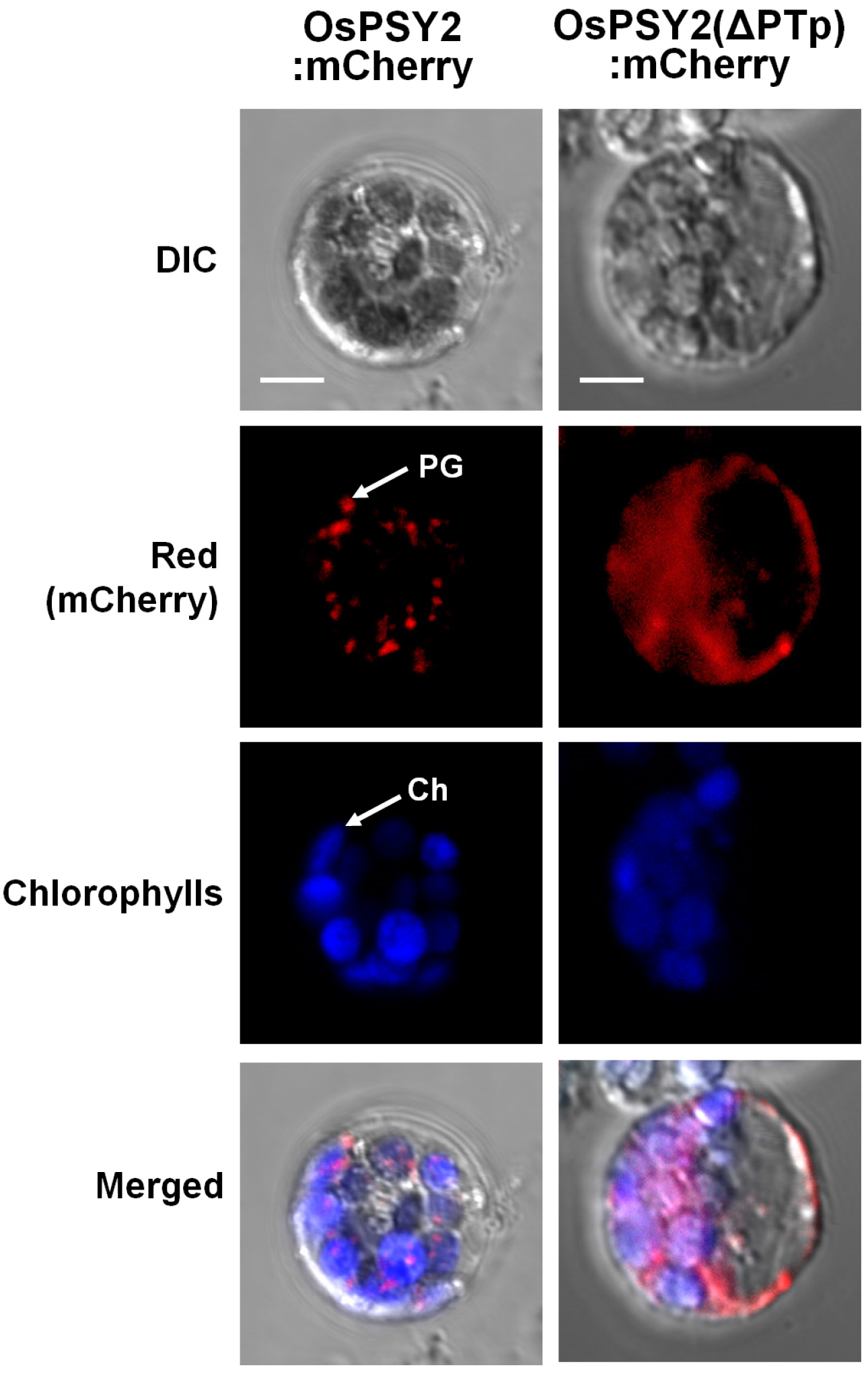
2.2. Subcellular Localization of OsPSY2 in Rice Protoplasts
2.3. Modification of the PTp DNA Structure for Improved Applications in Plant Biotechnology
2.4. Subcellular Localization of sGFP after N-Terminal Fusion of the PTp and stPTp Sequences
3. Discussion
4. Materials and Methods
4.1. Vector Construction
4.2. Protoplast Preparation
4.3. Vector DNA Transfection and Microscopy Analysis
4.4. Molecular Analysis of sGFP Expression
4.5. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Greenwood, A.D.; Leech, R.M.; Williams, J.P. The osmiophilic globules of chloroplasts: I. Osmiophilic globules as a normal component of chloroplasts and their isolation and composition in Vicia faba L. Biochim. Biophys. Acta 1963, 78, 148–162. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K. Die Plastoglobuli. Protoplasma 1969, 68, 315–326. [Google Scholar] [CrossRef]
- Vidi, P.A.; Kanwischer, M.; Baginsky, S.; Austin, J.R.; Csucs, G.; Dormann, P.; Kessler, F.; Brehelin, C. Tocopherol cyclase (VTE1) localization and vitamin E accumulation in chloroplast plastoglobule lipoprotein particles. J. Biol. Chem. 2006, 281, 11225–11234. [Google Scholar] [CrossRef] [PubMed]
- Ytterberg, A.J.; Peltier, J.B.; van Wijk, K.J. Protein profiling of plastoglobules in chloroplasts and chromoplasts. A surprising site for differential accumulation of metabolic enzymes. Plant Physiol. 2006, 140, 984–997. [Google Scholar] [CrossRef] [PubMed]
- Brehelin, C.; Kessler, F.; van Wijk, K.J. Plastoglobules: Versatile lipoprotein particles in plastids. Trends Plant Sci. 2007, 12, 260–266. [Google Scholar] [CrossRef] [PubMed]
- Schweiggert, R.M.; Steingass, C.B.; Heller, A.; Esquivel, P.; Carle, R. Characterization of chromoplasts and carotenoids of red- and yellow-fleshed papaya (Carica papaya L.). Planta 2011, 234, 1031–1044. [Google Scholar] [CrossRef] [PubMed]
- Davidi, L.; Levin, Y.; Ben-Dor, S.; Pick, U. Proteome analysis of cytoplasmatic and plastidic β-carotene lipid droplets in Dunaliella bardawil. Plant Physiol. 2015, 167, 60–79. [Google Scholar] [CrossRef] [PubMed]
- Rottet, S.; Besagni, C.; Kessler, F. The role of plastoglobules in thylakoid lipid remodeling during plant development. Biochim. Biophys. Acta 2015, 1847, 889–899. [Google Scholar] [CrossRef] [PubMed]
- Bai, C.; Rivera, S.M.; Medina, V.; Alves, R.; Vilaprinyo, E.; Sorribas, A.; Canela, R.; Capell, T.; Sandmann, G.; Christou, P.; et al. An in vitro system for the rapid functional characterization of genes involved in carotenoid biosynthesis and accumulation. Plant J. 2014, 77, 464–475. [Google Scholar] [CrossRef] [PubMed]
- Zeng, Y.; Du, J.; Wang, L.; Pan, Z.; Xu, Q.; Xiao, S.; Deng, X. A comprehensive analysis of chromoplast differentiation reveals complex protein changes associated with plastoglobule biogenesis and remodeling of protein systems in sweet orange flesh. Plant Physiol. 2015, 168, 1648–1665. [Google Scholar] [CrossRef] [PubMed]
- Vidi, P.A.; Kessler, F.; Brehelin, C. Plastoglobules: A new address for targeting recombinant proteins in the chloroplast. BMC Biotechnol. 2007, 7, 4. [Google Scholar] [CrossRef] [PubMed]
- Giuliano, G. Plant carotenoids: Genomics meets multi-gene engineering. Curr. Opin. Plant Biol. 2014, 19, 111–117. [Google Scholar] [CrossRef] [PubMed]
- Shumskaya, M.; Wurtzel, E.T. The carotenoid biosynthetic pathway: Thinking in all dimensions. Plant Sci. 2013, 208, 58–63. [Google Scholar] [CrossRef] [PubMed]
- Ye, X.; Al-Babili, S.; Kloti, A.; Zhang, J.; Lucca, P.; Beyer, P.; Potrykus, I. Engineering the provitamin A (β-carotene) biosynthetic pathway into (carotenoid-free) rice endosperm. Science 2000, 287, 303–305. [Google Scholar] [CrossRef] [PubMed]
- Ha, S.H.; Liang, Y.S.; Jung, H.; Ahn, M.J.; Suh, S.C.; Kweon, S.J.; Kim, D.H.; Kim, Y.M.; Kim, J.K. Application of two bicistronic systems involving 2A and IRES sequences to the biosynthesis of carotenoids in rice endosperm. Plant Biotechnol. J. 2010, 8, 928–938. [Google Scholar] [CrossRef] [PubMed]
- Kim, M.J.; Kim, J.K.; Kim, H.J.; Pak, J.H.; Lee, J.H.; Kim, D.H.; Choi, H.K.; Jung, H.W.; Lee, J.D.; Chung, Y.S.; et al. Genetic modification of the soybean to enhance the β-carotene content through seed-specific expression. PLoS ONE 2012, 7, e48287. [Google Scholar] [CrossRef] [PubMed]
- Shumskaya, M.; Bradbury, L.M.; Monaco, R.R.; Wurtzel, E.T. Plastid localization of the key carotenoid enzyme phytoene synthase is altered by isozyme, allelic variation, and activity. Plant Cell 2012, 24, 3725–3741. [Google Scholar] [CrossRef] [PubMed]
- Singh, V.K.; Govindarajan, R.; Naik, S.; Kumar, A. The Effect of hairpin structure on PCR amplification efficiency. Mol. Biol. Today 2000, 1, 67–69. [Google Scholar]
- You, M.K.; Lim, S.H.; Kim, M.J.; Jeong, Y.S.; Lee, M.G.; Ha, S.H. Improvement of the fluorescence intensity during a flow cytometric analysis for rice protoplasts by localization of a green fluorescent protein into chloroplasts. Int. J. Mol. Sci. 2015, 16, 788–804. [Google Scholar] [CrossRef] [PubMed]
- Welsch, R.; Wust, F.; Bar, C.; Al-Babili, S.; Beyer, P. A third phytoene synthase is devoted to abiotic stress-induced abscisic acid formation in rice and defines functional diversification of phytoene synthase genes. Plant Physiol. 2008, 147, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Jang, I.-C.; Nahm, B.H.; Kim, J.-K. Subcellular targeting of green fluorescent protein to protein to plastids in transgenic rice plants provides a high-level expression system. Mol. Breed. 1999, 5, 453–461. [Google Scholar] [CrossRef]
- Lutcke, H.A.; Chow, K.C.; Mickel, F.S.; Moss, K.A.; Kern, H.F.; Scheele, G.A. Selection of AUG initiation codons differs in plants and animals. EMBO J. 1987, 6, 43–48. [Google Scholar] [PubMed]
- Hartley, J.L.; Temple, G.F.; Brasch, M.A. DNA cloning using in vitro site-specific recombination. Genome Res. 2000, 10, 1788–1795. [Google Scholar] [CrossRef] [PubMed]
- Higuchi, R.; Krummel, B.; Saiki, R.K. A general method of in vitro preparation and specific mutagenesis of DNA fragments: Study of protein and DNA interactions. Nucleic Acids Res. 1988, 16, 7351–7367. [Google Scholar] [CrossRef] [PubMed]
- Dubin, M.J.; Bowler, C.; Benvenuto, G. A modified Gateway cloning strategy for overexpressing tagged proteins in plants. Plant Methods 2008, 4, 3. [Google Scholar] [CrossRef] [PubMed]
- Bart, R.; Chern, M.; Park, C.J.; Bartley, L.; Ronald, P.C. A novel system for gene silencing using siRNAs in rice leaf and stem-derived protoplasts. Plant Methods 2006, 2, 13. [Google Scholar] [CrossRef] [PubMed]
- Song, M.H.; Lim, S.H.; Kim, J.K.; Jung, E.S.; Maria John, K.M.; You, M.K.; Ahn, S.N.; Lee, C.H.; Ha, S.H. In planta cleavage of carotenoids by Arabidopsis carotenoid cleavage dioxygenase 4 in transgenic rice plants. Plant Biotechnol. Rep. 2016, 10, 291–300. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein (Size, aa) | ChloroP a | TMpred b | TopPred c | HMMTOP d | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Tp * | TM † No. | TM 1 | TM 2 | TM 3 | TM No. | TM 1 | TM 2 | TM 3 | TM No. | TM 1 | TM 2 | |
| OsPSY1 (420) | 21 | 2 | 247–265 | 271–293 | 2 | 66–86 | 247–267 | 2 | 249–266 | 275–292 | ||
| OsPSY2 (398) | 80 | 3 | 1–22 | 48–66 | 233–252 | 3 | 1–21 | 46–66 | 233–253 | 2 | 230–252 | 261–278 |
| OsPSY3 (444) | 54 | 1 | 264–285 | 1 | 267–287 | 1 | 264–285 | |||||
© 2016 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC-BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
You, M.K.; Kim, J.H.; Lee, Y.J.; Jeong, Y.S.; Ha, S.-H. Plastoglobule-Targeting Competence of a Putative Transit Peptide Sequence from Rice Phytoene Synthase 2 in Plastids. Int. J. Mol. Sci. 2017, 18, 18. https://doi.org/10.3390/ijms18010018
You MK, Kim JH, Lee YJ, Jeong YS, Ha S-H. Plastoglobule-Targeting Competence of a Putative Transit Peptide Sequence from Rice Phytoene Synthase 2 in Plastids. International Journal of Molecular Sciences. 2017; 18(1):18. https://doi.org/10.3390/ijms18010018
Chicago/Turabian StyleYou, Min Kyoung, Jin Hwa Kim, Yeo Jin Lee, Ye Sol Jeong, and Sun-Hwa Ha. 2017. "Plastoglobule-Targeting Competence of a Putative Transit Peptide Sequence from Rice Phytoene Synthase 2 in Plastids" International Journal of Molecular Sciences 18, no. 1: 18. https://doi.org/10.3390/ijms18010018