
Derivate Isocorydine (d-ICD) Suppresses Migration and Invasion of Hepatocellular Carcinoma Cell by Downregulating ITGA1 Expression


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
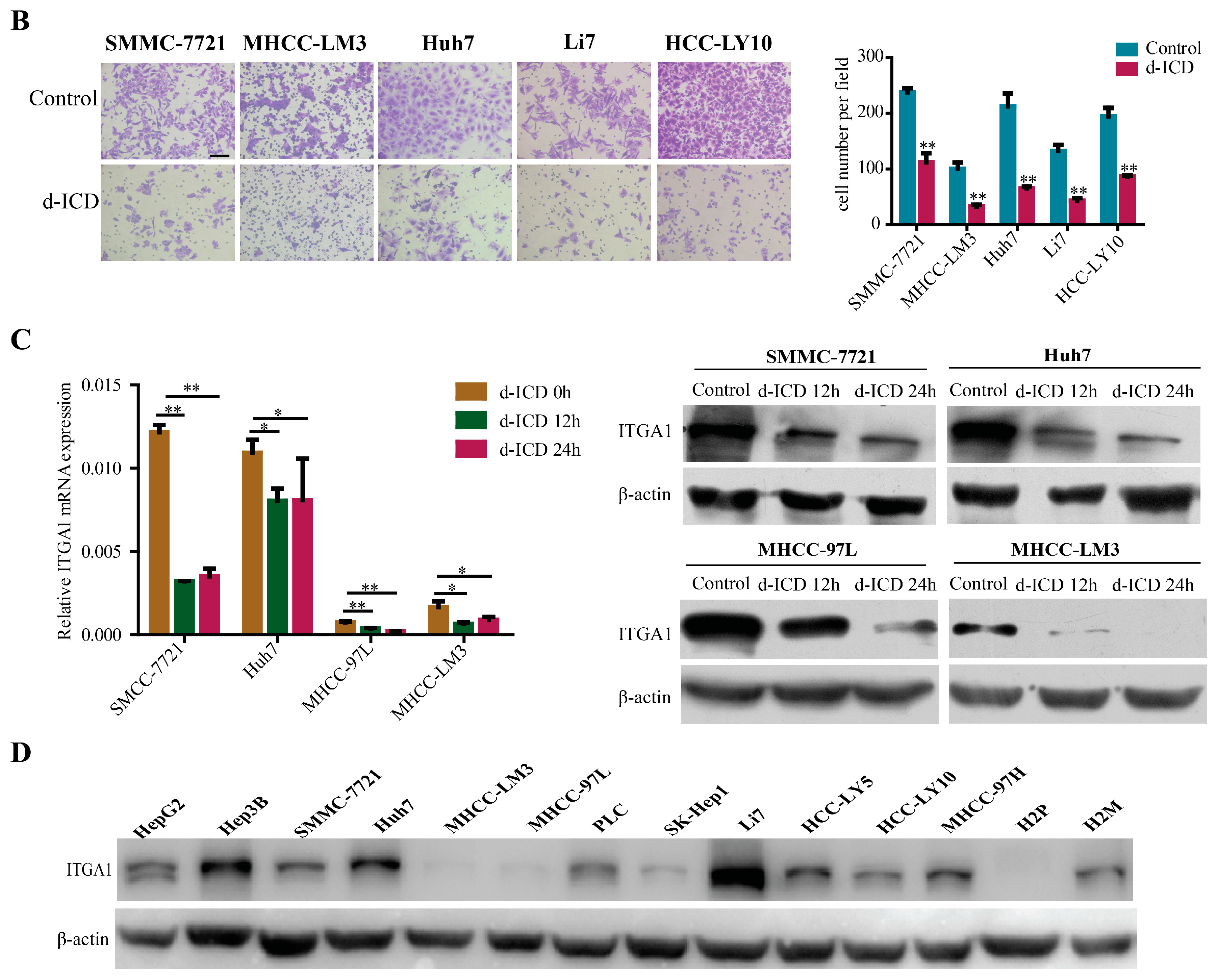
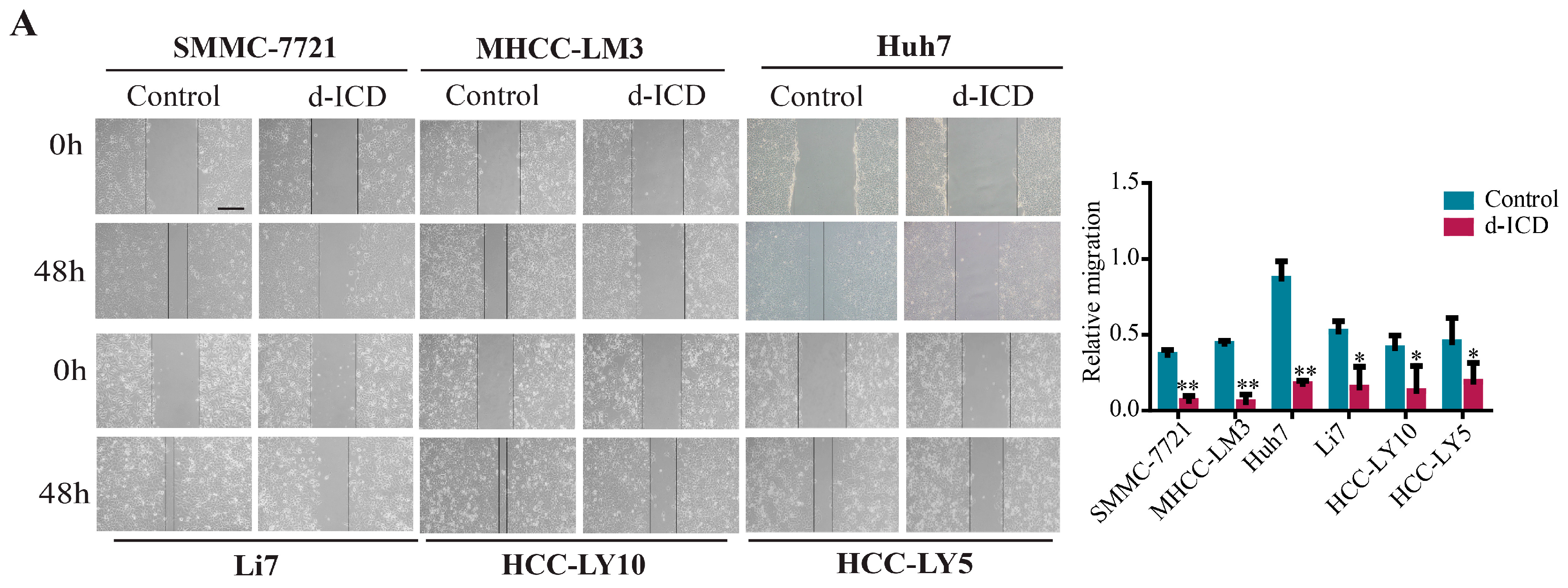
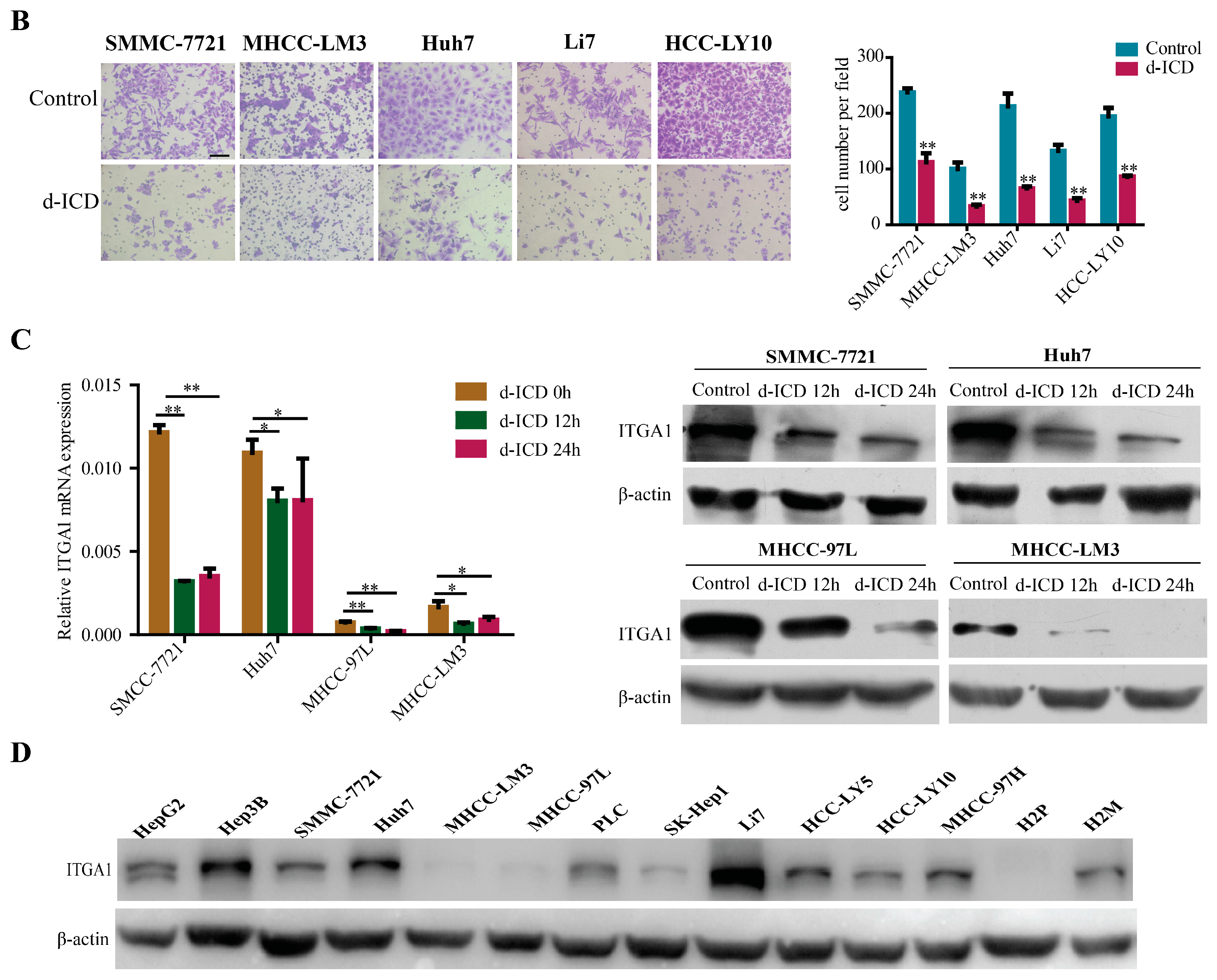
2.1. d-ICD Inhibits HCC Cell Migration and Invasion In Vitro and Downregulates ITGA1 Expression
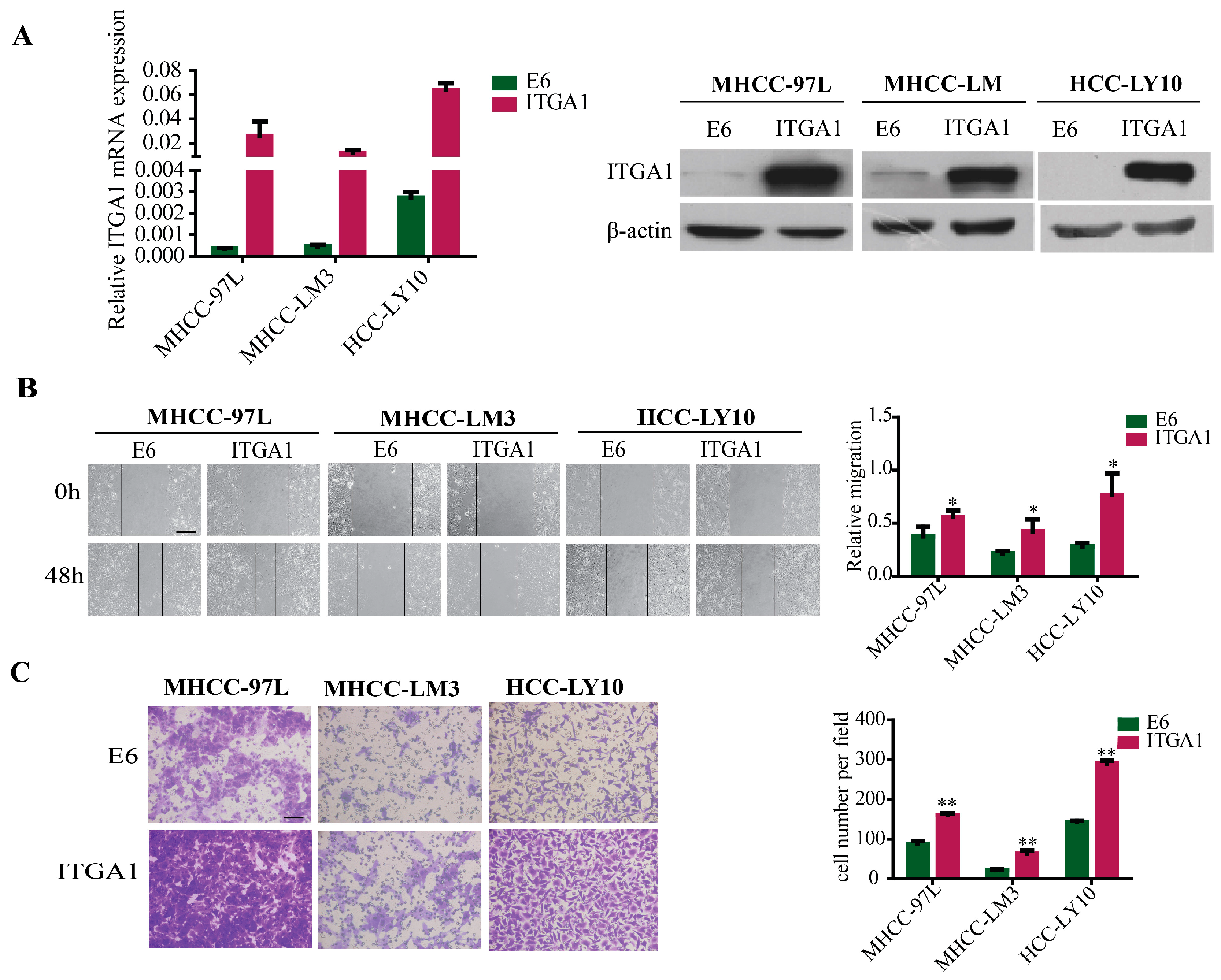
2.2. Overexpression of ITGA1 Promotes HCC Cell Migration and Invasion In Vitro
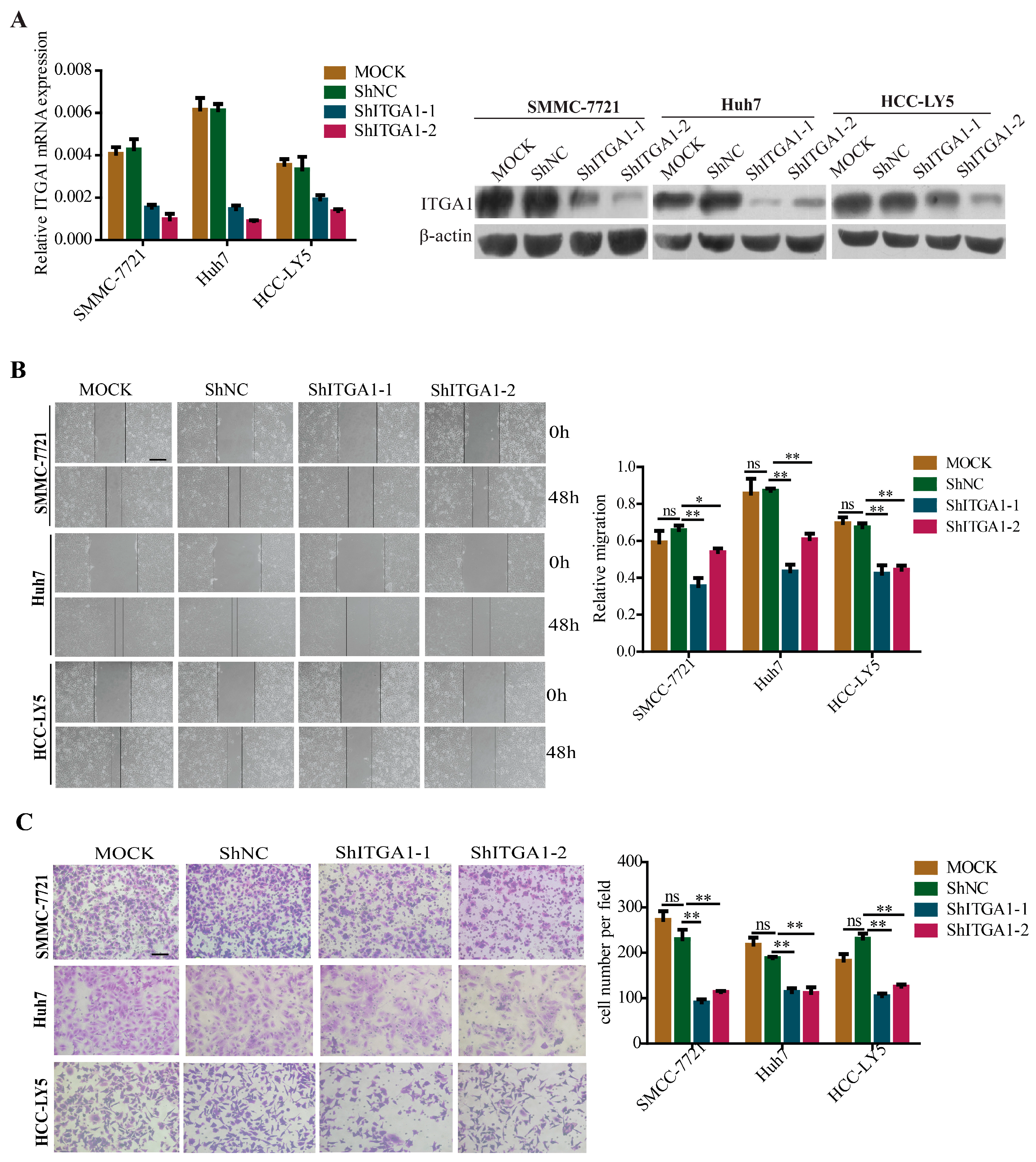
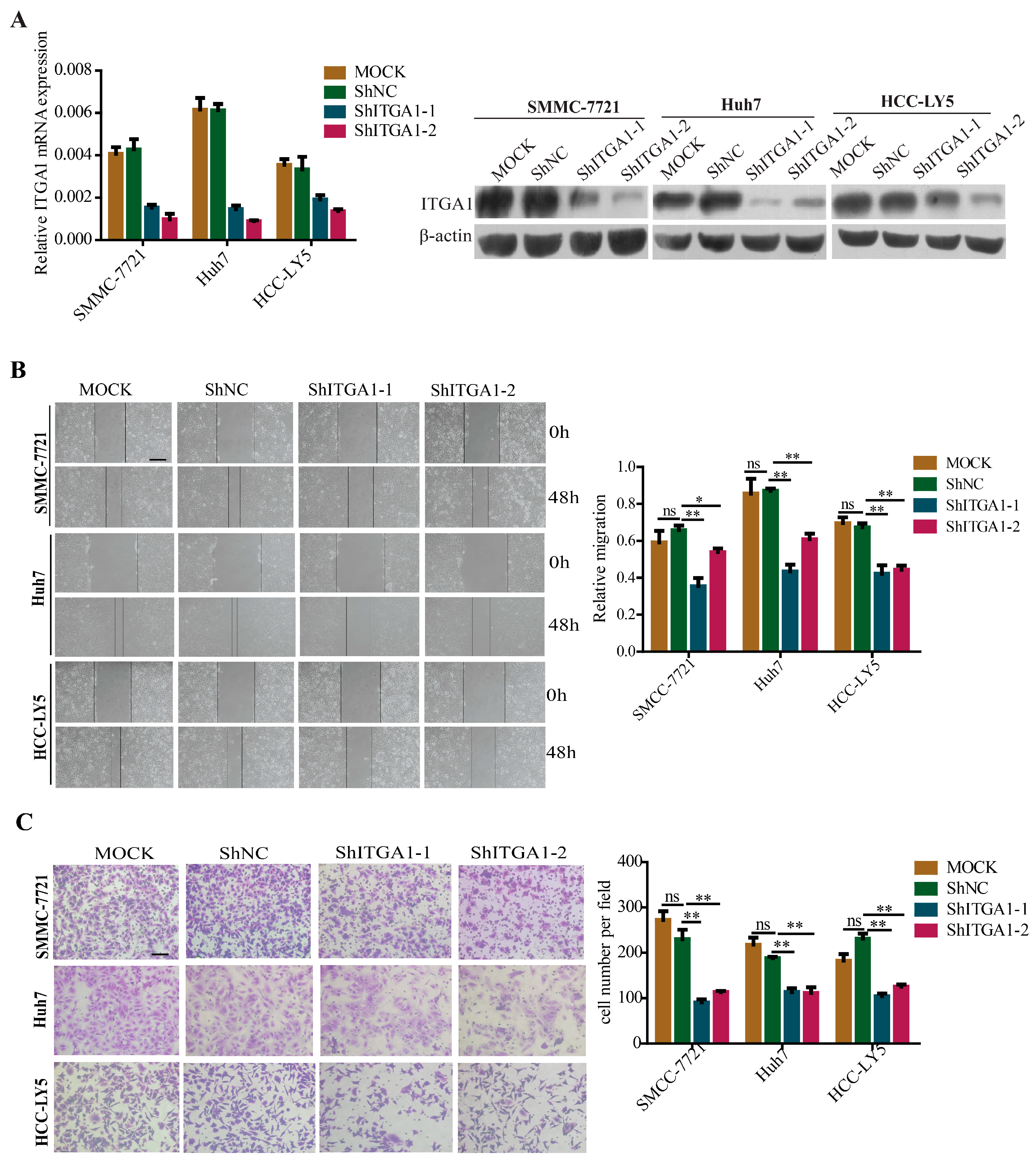
2.3. Silencing ITGA1 Expression Inhibits HCC Cell Migration and Invasion In Vitro
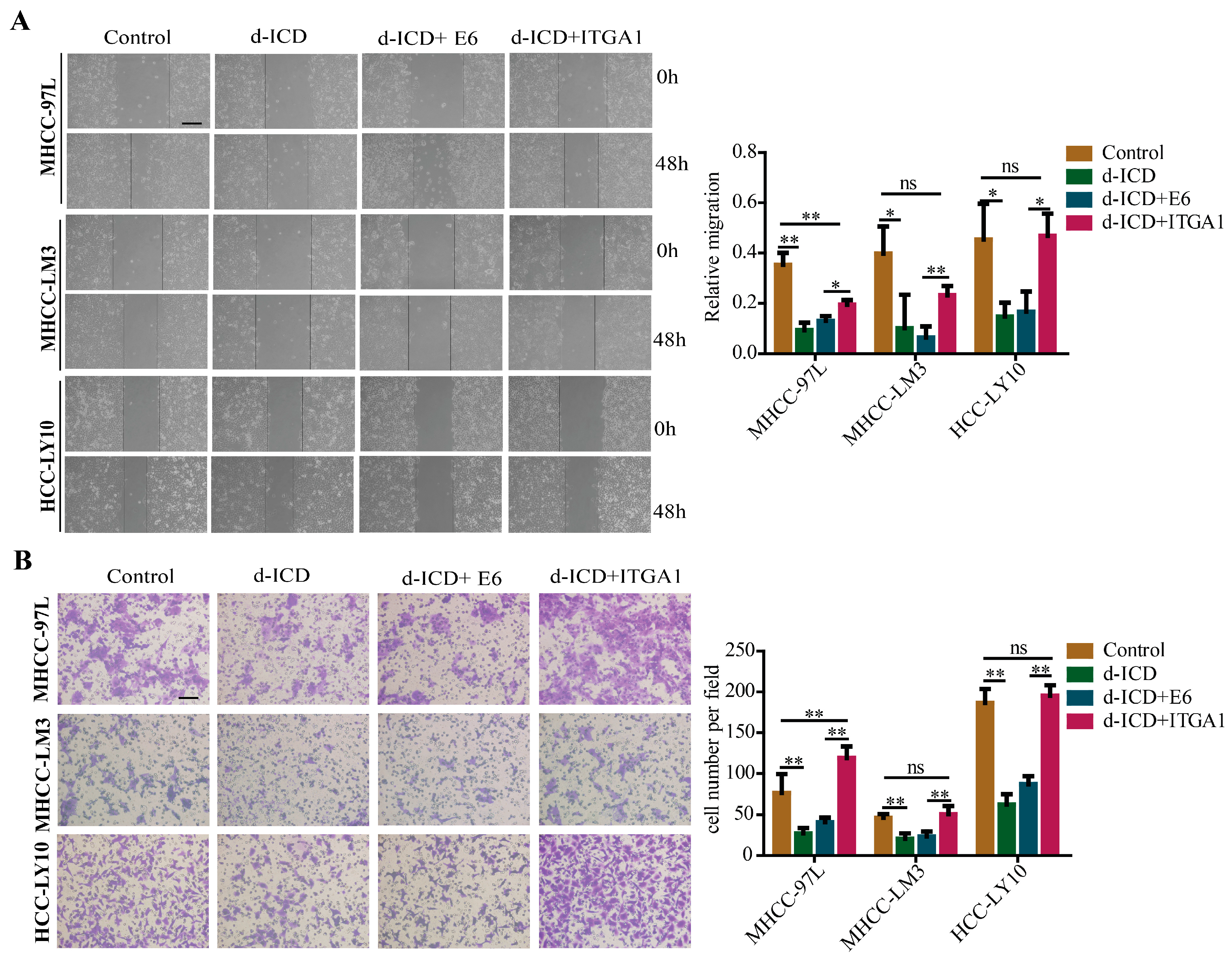
2.4. Overexpression of ITGA1 Partly Rescues d-ICD-Induced Migration and Invasion Inhibition in HCC Cells
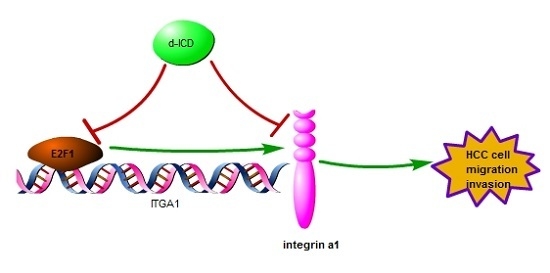
2.5. E2F1 Upregulates ITGA1 Expression and d-ICD Inhibits E2F1 Expression in HCC Cells
3. Discussion
4. Materials and Methods
4.1. Cell Lines and Cell Culture
4.2. Western Blotting Analyses
4.3. Quantitative Real-Time Polymerase Chain Reaction (qRT-PCR)
4.4. Plasmid Construction, Lentivirus Production, and Cell Transfection
4.5. Wound Healing Assay
4.6. Trans-Well Invasion Assay
4.7. Dual Luciferase Reporter Assay
4.8. Chromatin Immunoprecipitation Assay (ChIP)
4.9. Cell Apoptosis Analysis
4.10. Cell Proliferation Assays
4.11. Statistical Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Torre, L.A.; Bray, F.; Siegel, R.L.; Ferlay, J.; Lortet-Tieulent, J.; Jemal, A. Global cancer statistics, 2012. Cancer J. Clin. 2015, 65, 87–108. [Google Scholar] [CrossRef] [PubMed]
- Marquardt, J.U.; Thorgeirsson, S.S. SnapShot: Hepatocellular carcinoma. Cancer Cell 2014, 25, 550. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Hou, H.; Lu, P.; Zhang, L.; Zhao, F.; Ge, C.; Wang, T.; Yao, M.; Li, J. Isocorydine inhibits cell proliferation in hepatocellular carcinoma cell lines by inducing G2/M cell cycle arrest and apoptosis. PLoS ONE 2012, 7, e36808. [Google Scholar] [CrossRef] [PubMed]
- Lu, P.; Sun, H.; Zhang, L.; Hou, H.; Zhang, L.; Zhao, F.; Ge, C.; Yao, M.; Wang, T.; Li, J. Isocorydine targets the drug-resistant cellular side population through PDCD4-related apoptosis in hepatocellular carcinoma. Mol. Med. 2012, 18, 1136–1146. [Google Scholar] [CrossRef] [PubMed]
- Zhong, M.; Liu, Y.; Liu, J.; Di, D.; Xu, M.; Yang, Y.; Li, W.; Chen, Y.; Liu, J. Isocorydine derivatives and their anticancer activities. Molecules 2014, 19, 12099–12115. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.; Tian, H.; Li, M.; Ge, C.; Zhao, F.; Zhang, L.; Li, H.; Liu, J.; Wang, T.; Yao, M.; et al. Derivate isocorydine inhibits cell proliferation in hepatocellular carcinoma cell lines by inducing G2/M cell cycle arrest and apoptosis. Tumour Biol. 2016, 37, 5951–5961. [Google Scholar] [CrossRef] [PubMed]
- Li, M.; Zhang, L.; Ge, C.; Chen, L.; Fang, T.; Li, H.; Tian, H.; Liu, J.; Chen, T.; Jiang, G.; et al. An isocorydine derivative (d-ICD) inhibits drug resistance by downregulating IGF2BP3 expression in hepatocellular carcinoma. Oncotarget 2015, 6, 25149–25160. [Google Scholar] [CrossRef] [PubMed]
- Barczyk, M.; Carracedo, S.; Gullberg, D. Integrins. Cell Tissue Res. 2010, 339, 269–280. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, I.; Korhonen, M.; Kariniemi, A.L.; Gould, V.E.; Laitinen, L.; Ylanne, J. Integrins in human cells and tumors. Cell Differ. Dev. 1990, 32, 215–227. [Google Scholar] [CrossRef]
- Schadendorf, D.; Gawlik, C.; Haney, U.; Ostmeier, H.; Suter, L.; Czarnetzki, B.M. Tumour progression and metastatic behaviour in vivo correlates with integrin expression on melanocytic tumours. J. Pathol. 1993, 170, 429–434. [Google Scholar] [CrossRef] [PubMed]
- Sanders, R.J.; Mainiero, F.; Giancotti, F.G. The role of integrins in tumorigenesis and metastasis. Cancer Investig. 1998, 16, 329–344. [Google Scholar] [CrossRef] [PubMed]
- Zallocchi, M.; Johnson, B.M.; Meehan, D.T.; Delimont, D.; Cosgrove, D. α1β1 integrin/Rac1-dependent mesangial invasion of glomerular capillaries in Alport syndrome. Am. J. Pathol. 2013, 183, 1269–1280. [Google Scholar] [CrossRef] [PubMed]
- Senger, D.R.; Perruzzi, C.A.; Streit, M.; Koteliansky, V.E.; de Fougerolles, A.R.; Detmar, M. The α1β1 and α2β1 integrins provide critical support for vascular endothelial growth factor signaling, endothelial cell migration, and tumor angiogenesis. Am. J. Pathol. 2002, 160, 195–204. [Google Scholar] [CrossRef]
- Zhang, R.; Wang, N.; Zhang, M.; Zhang, L.N.; Guo, Z.X.; Luo, X.G.; Zhou, H.; He, H.P.; Zhang, T.C. Rho/MRTF-A-induced integrin expression regulates angiogenesis in differentiated multipotent mesenchymal stem cells. Stem Cells Int. 2015, 2015, 534758. [Google Scholar] [CrossRef] [PubMed]
- Schadendorf, D.; Fichtner, I.; Makki, A.; Alijagic, S.; Kupper, M.; Mrowietz, U.; Henz, B.M. Metastatic potential of human melanoma cells in nude mice—Characterisation of phenotype, cytokine secretion and tumour-associated antigens. Br. J. Cancer 1996, 74, 194–199. [Google Scholar] [CrossRef] [PubMed]
- Gulubova, M.V. Collagen type IV, laminin, α-smooth muscle actin (αSMA), α1 and α6 integrins expression in the liver with metastases from malignant gastrointestinal tumours. Clin. Exp. Metastasis 2004, 21, 485–494. [Google Scholar] [CrossRef] [PubMed]
- Boudjadi, S.; Carrier, J.C.; Beaulieu, J.F. Integrin α1 subunit is up-regulated in colorectal cancer. Biomark. Res. 2013, 1, 16. [Google Scholar] [CrossRef] [PubMed]
- Fukuda, K.; Saikawa, Y.; Yagi, H.; Wada, N.; Takahashi, T.; Kitagawa, Y. Role of integrin α1 subunits in gastric cancer patients with peritoneal dissemination. Mol. Med. Rep. 2012, 5, 336–340. [Google Scholar] [PubMed]
- Chen, X.; Su, Y.; Fingleton, B.; Acuff, H.; Matrisian, L.M.; Zent, R.; Pozzi, A. An orthotopic model of lung cancer to analyze primary and metastatic NSCLC growth in integrin α1-null mice. Clin. Exp. Metastasis 2005, 22, 185–193. [Google Scholar] [CrossRef] [PubMed]
- Macias-Perez, I.; Borza, C.; Chen, X.; Yan, X.; Ibanez, R.; Mernaugh, G.; Matrisian, L.M.; Zent, R.; Pozzi, A. Loss of integrin α1β1 ameliorates Kras-induced lung cancer. Cancer Res. 2008, 68, 6127–6135. [Google Scholar] [CrossRef] [PubMed]
- Cam, H.; Dynlacht, B.D. Emerging roles for E2F: Beyond the G1/S transition and DNA replication. Cancer Cell 2003, 3, 311–316. [Google Scholar] [CrossRef]
- La Thangue, N.B. The yin and yang of E2F-1: Balancing life and death. Nat. Cell Biol. 2003, 5, 587–589. [Google Scholar] [CrossRef] [PubMed]
- Jiang, H.; Martin, V.; Gomez-Manzano, C.; Johnson, D.G.; Alonso, M.; White, E.; Xu, J.; McDonnell, T.J.; Shinojima, N.; Fueyo, J. The RB-E2F1 pathway regulates autophagy. Cancer Res. 2010, 70, 7882–7893. [Google Scholar] [CrossRef] [PubMed]
- Palaiologou, M.; Koskinas, J.; Karanikolas, M.; Fatourou, E.; Tiniakos, D.G. E2F-1 is overexpressed and pro-apoptotic in human hepatocellular carcinoma. Int. J. Pathol. 2012, 460, 439–446. [Google Scholar] [CrossRef] [PubMed]
- Pozzi, A.; Moberg, P.E.; Miles, L.A.; Wagner, S.; Soloway, P.; Gardner, H.A. Elevated matrix metalloprotease and angiostatin levels in integrin α1 knockout mice cause reduced tumor vascularization. Proc. Natl. Acad. Sci. USA 2000, 97, 2202–2207. [Google Scholar] [CrossRef] [PubMed]
- Gardner, H. Integrin α1β1. Adv. Exp. Med. Biol. 2014, 819, 21–39. [Google Scholar] [PubMed]
- Yasoshima, T.; Denno, R.; Kawaguchi, S.; Sato, N.; Okada, Y.; Ura, H.; Kikuchi, K.; Hirata, K. Establishment and characterization of human gastric carcinoma lines with high metastatic potential in the liver: Changes in integrin expression associated with the ability to metastasize in the liver of nude mice. Jpn. J. Cancer Res. 1996, 87, 153–160. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Wen, D.; Dong, L.; Tang, J.; Liu, D.; Liu, Y.; Tao, Z.; Gao, D.; Sun, H.; Cao, Y.; et al. Establishment of monoclonal HCC cell lines with organ site-specific tropisms. BMC Cancer 2015, 15, 678. [Google Scholar] [CrossRef] [PubMed]
- Meier, C.; Spitschak, A.; Abshagen, K.; Gupta, S.; Mor, J.M.; Wolkenhauer, O.; Haier, J.; Vollmar, B.; Alla, V.; Putzer, B.M. Association of RHAMM with E2F1 promotes tumour cell extravasation by transcriptional up-regulation of fibronectin. J. Pathol. 2014, 234, 351–364. [Google Scholar] [CrossRef] [PubMed]
- Conner, E.A.; Lemmer, E.R.; Omori, M.; Wirth, P.J.; Factor, V.M.; Thorgeirsson, S.S. Dual functions of E2F1 in a transgenic mouse model of liver carcinogenesis. Oncogene 2000, 19, 5054–5062. [Google Scholar] [CrossRef] [PubMed]
- Wang, S.N.; Wang, L.T.; Sun, D.P.; Chai, C.Y.; Hsi, E.; Kuo, H.T.; Yokoyama, K.K.; Hsu, S.H. Intestine-specific homeobox (ISX) upregulates E2F1 expression and related oncogenic activities in HCC. Oncotarget 2016, 7, 36924–36939. [Google Scholar] [CrossRef] [PubMed]
- Tian, H.; Ge, C.; Li, H.; Zhao, F.; Hou, H.; Chen, T.; Jiang, G.; Xie, H.; Cui, Y.; Yao, M.; et al. Ribonucleotide reductase M2B inhibits cell migration and spreading by early growth response protein 1-mediated phosphatase and tensin homolog/Akt1 pathway in hepatocellular carcinoma. Hepatology 2014, 59, 1459–1470. [Google Scholar] [CrossRef] [PubMed]
- Fang, T.; Cui, M.; Sun, J.; Ge, C.; Zhao, F.; Zhang, L.; Tian, H.; Zhang, L.; Chen, T.; Jiang, G.; et al. Orosomucoid 2 inhibits tumor metastasis and is upregulated by CCAAT/enhancer binding protein β in hepatocellular carcinomas. Oncotarget 2015, 6, 16106–16119. [Google Scholar] [CrossRef] [PubMed]


© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license ( http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, X.; Tian, H.; Li, H.; Ge, C.; Zhao, F.; Yao, M.; Li, J. Derivate Isocorydine (d-ICD) Suppresses Migration and Invasion of Hepatocellular Carcinoma Cell by Downregulating ITGA1 Expression. Int. J. Mol. Sci. 2017, 18, 514. https://doi.org/10.3390/ijms18030514
Liu X, Tian H, Li H, Ge C, Zhao F, Yao M, Li J. Derivate Isocorydine (d-ICD) Suppresses Migration and Invasion of Hepatocellular Carcinoma Cell by Downregulating ITGA1 Expression. International Journal of Molecular Sciences. 2017; 18(3):514. https://doi.org/10.3390/ijms18030514
Chicago/Turabian StyleLiu, Xiaoqin, Hua Tian, Hong Li, Chao Ge, Fangyu Zhao, Ming Yao, and Jinjun Li. 2017. "Derivate Isocorydine (d-ICD) Suppresses Migration and Invasion of Hepatocellular Carcinoma Cell by Downregulating ITGA1 Expression" International Journal of Molecular Sciences 18, no. 3: 514. https://doi.org/10.3390/ijms18030514