
Molecular Characterization and Expression Profiling of Tomato GRF Transcription Factor Family Genes in Response to Abiotic Stresses and Phytohormones


 ,
,
Abstract
:1. Introduction
2. Results
2.1. Identification and Sequence Analysis of SlGRF Genes and Their Putative Proteins
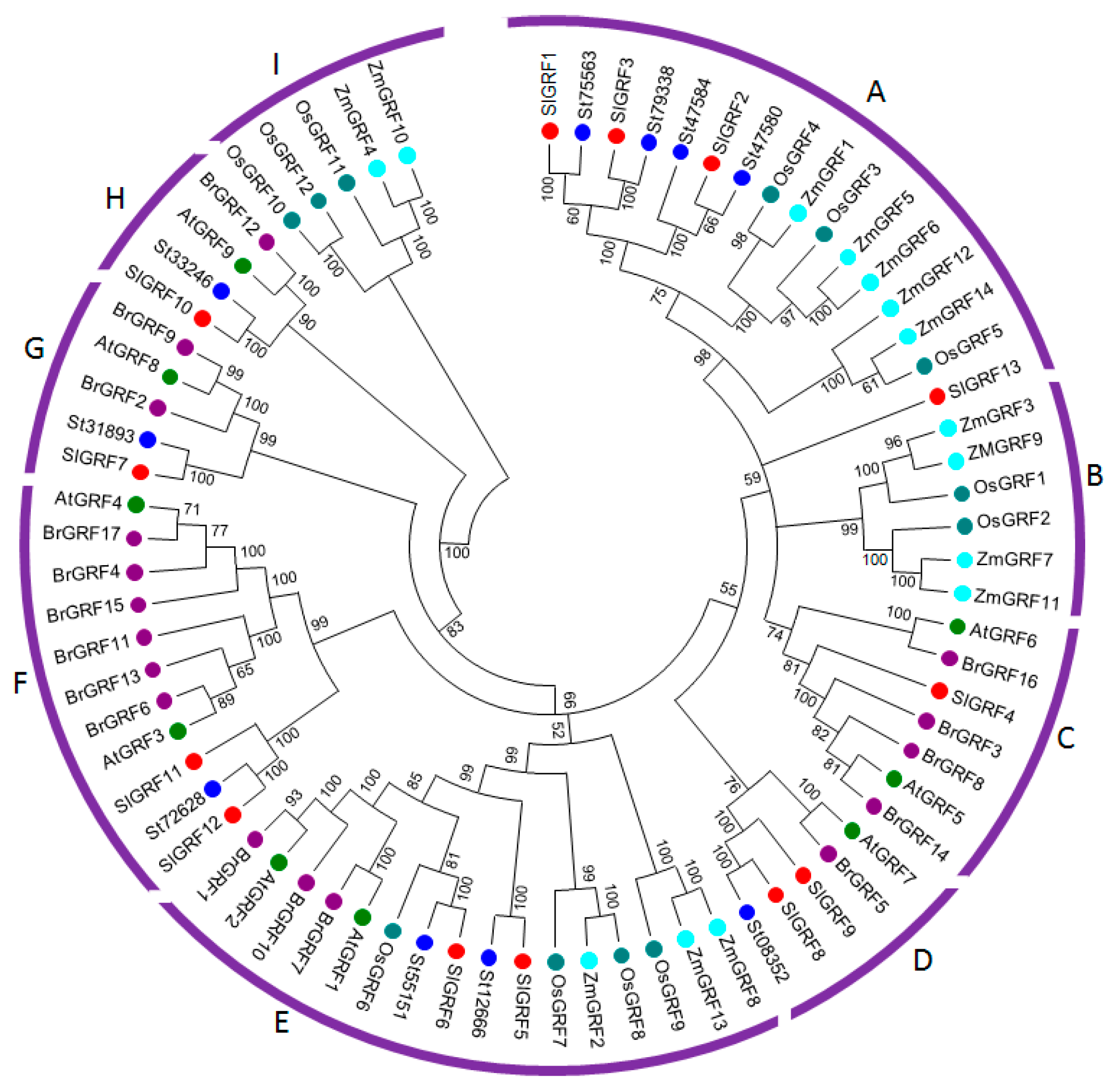
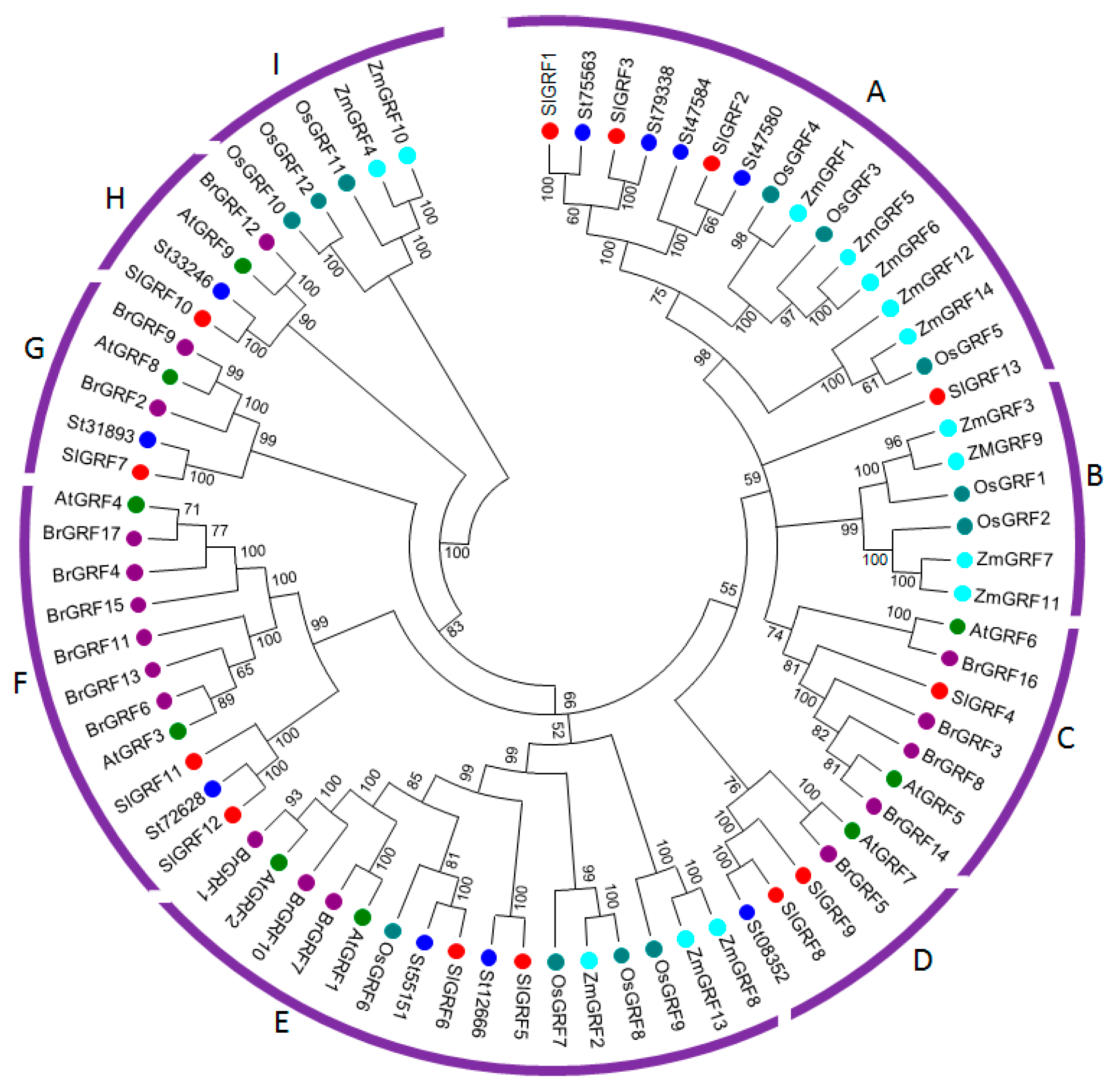
2.2. Evolutionary Analysis of SlGRF Proteins through Phylogenetic Classification
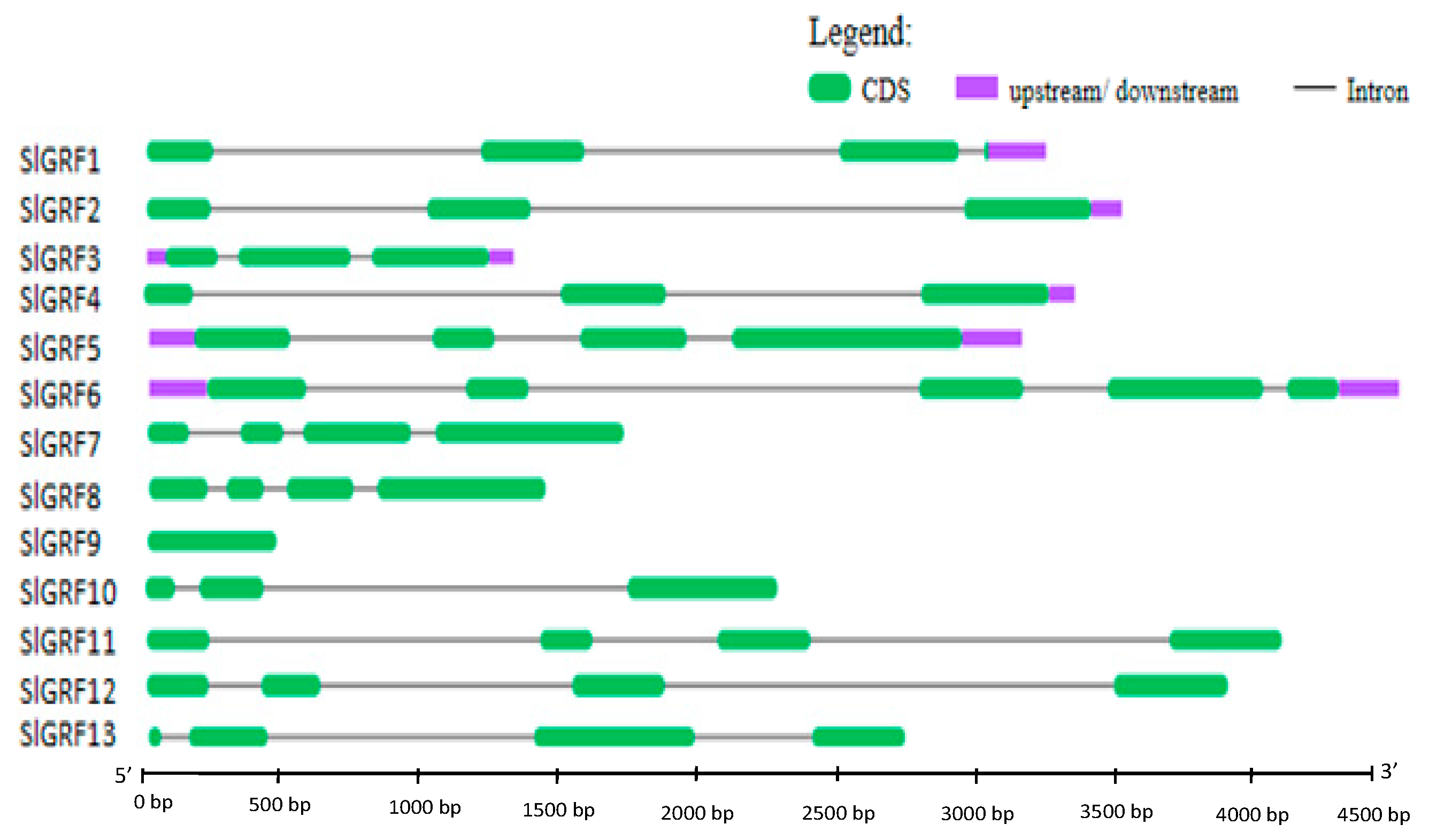
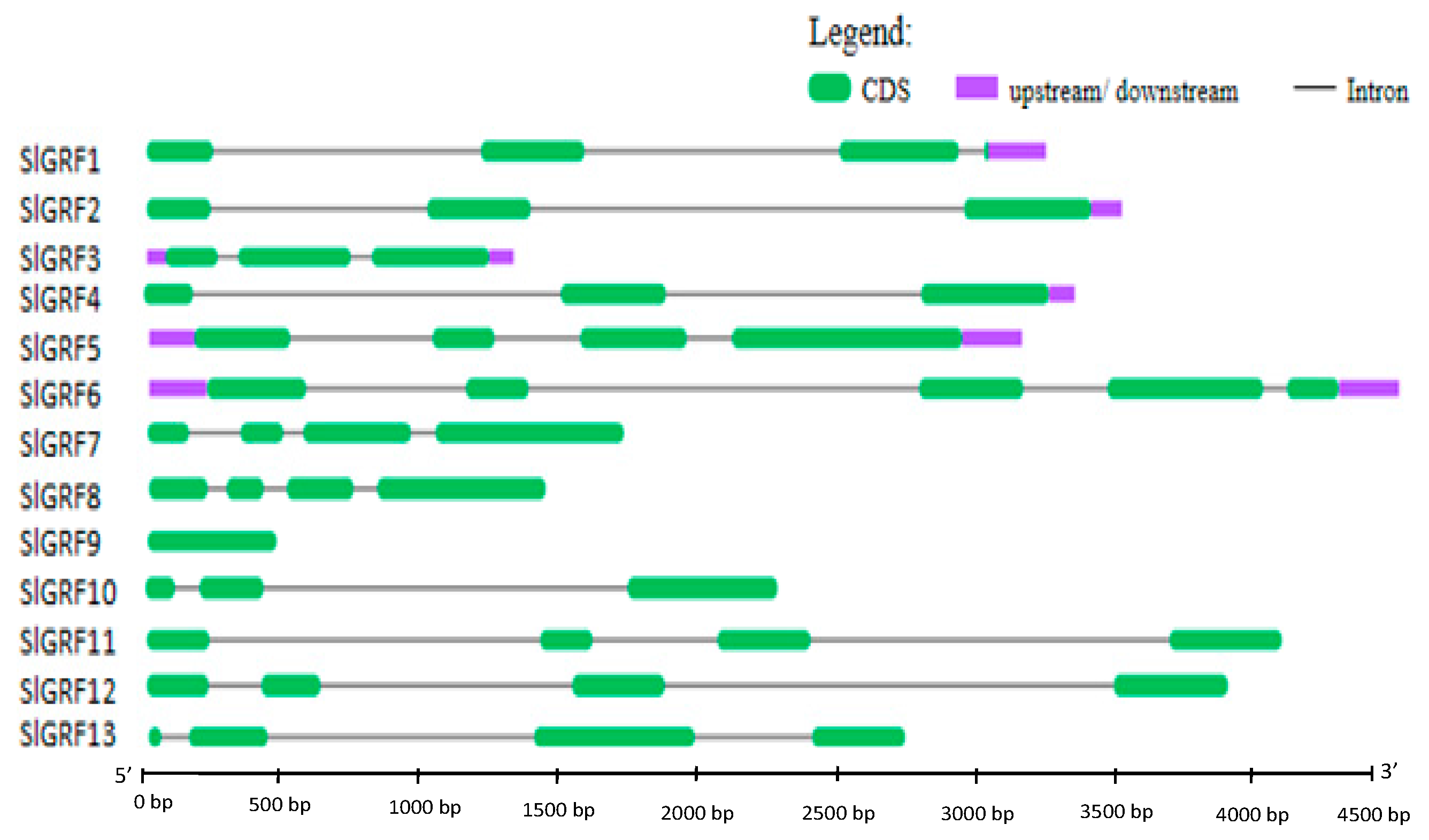
2.3. Structural Organization, and Chromosomal Location of SlGRF Genes
2.4. Putative Cis-Elements and Functional Analysis of SlGRF Genes
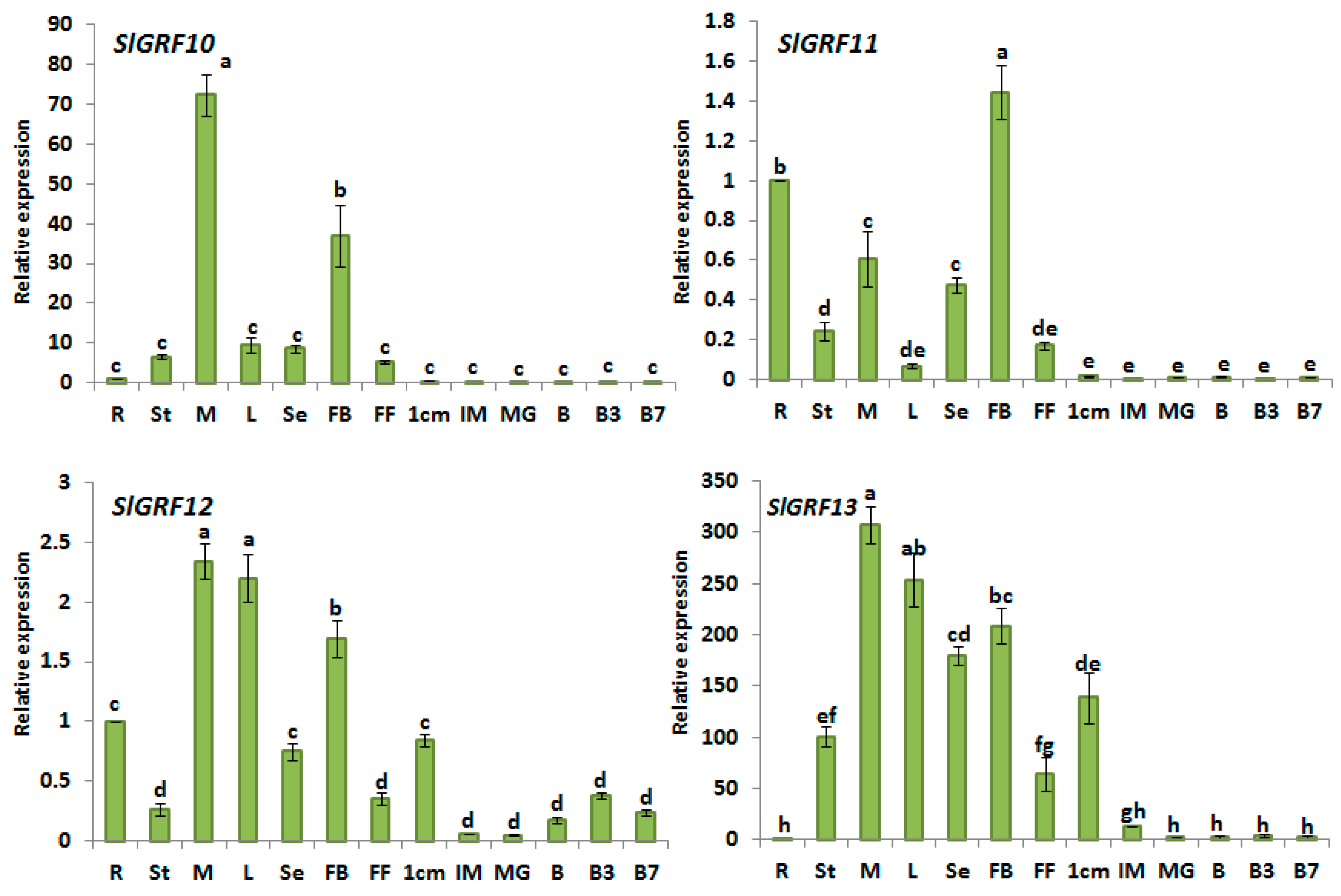
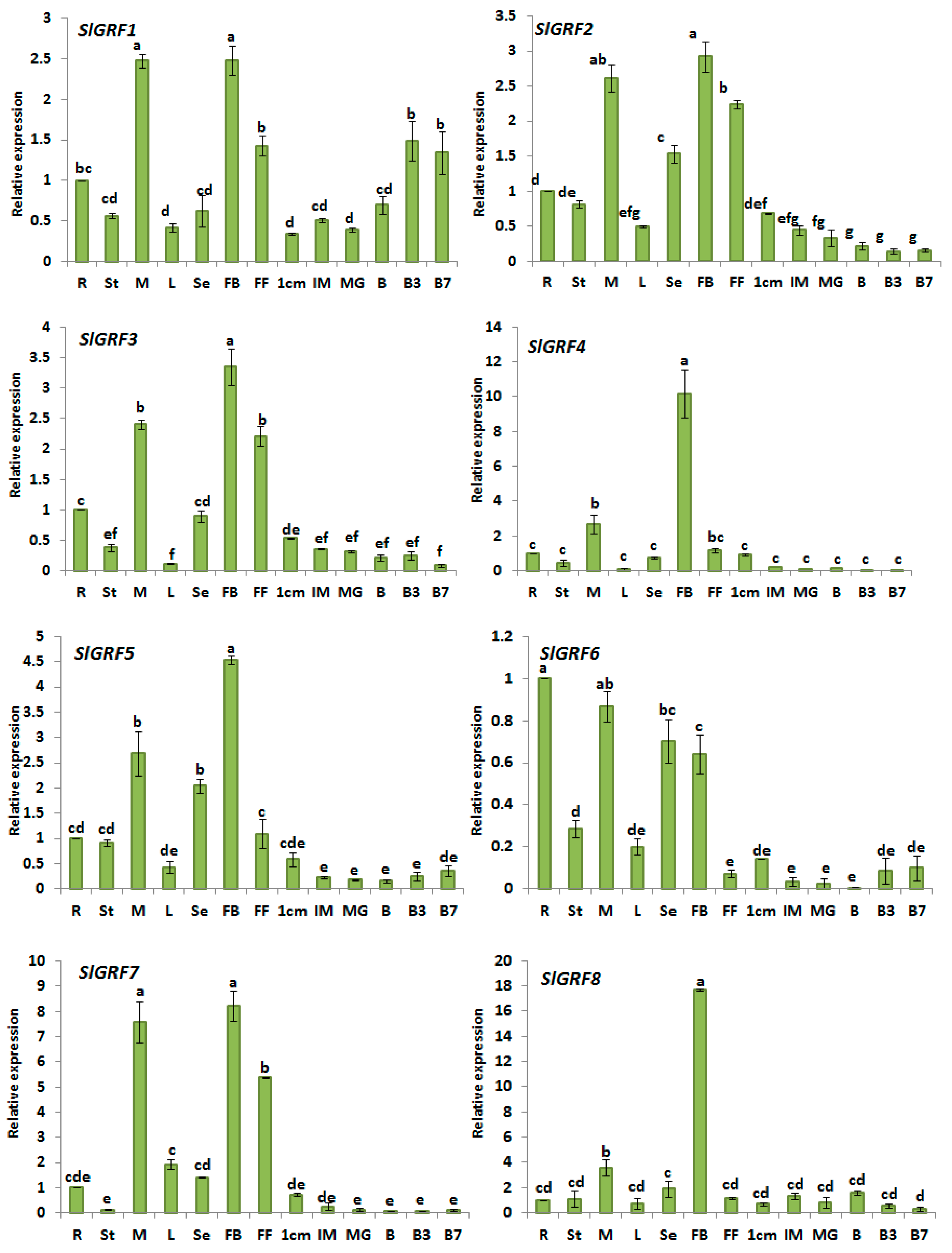
2.5. Expression Analysis of Tomato GRF Genes in Different Tissues
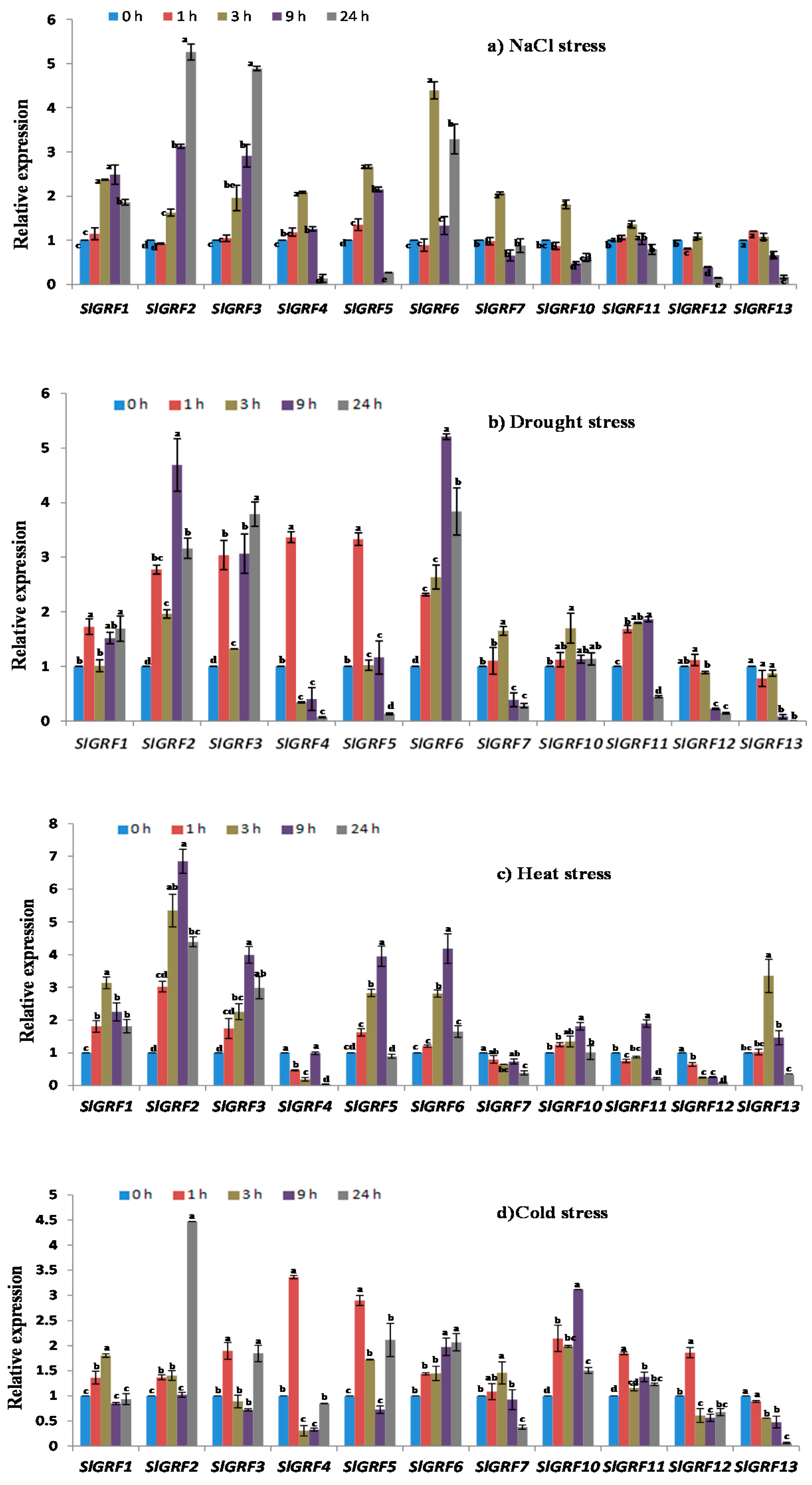
2.6. Expression Analysis of Tomato GRF Genes under Different Abiotic Stresses
2.6.1. NaCl Treatment
2.6.2. Drought Treatment
2.6.3. Heat Treatment
2.6.4. Cold Treatment
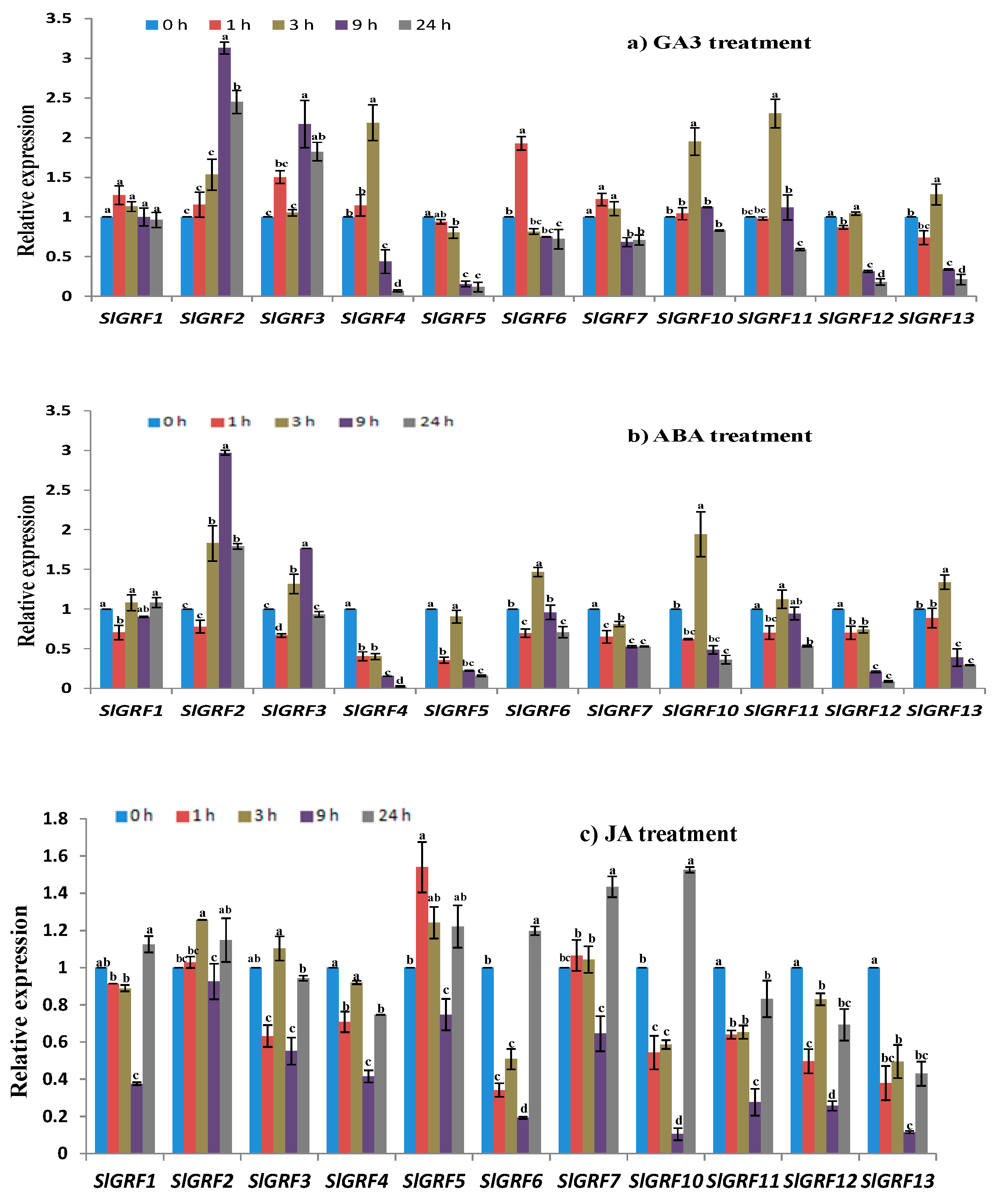
2.7. Expression Profiles of SlGRF Genes in Response to Phytohormone Treatments
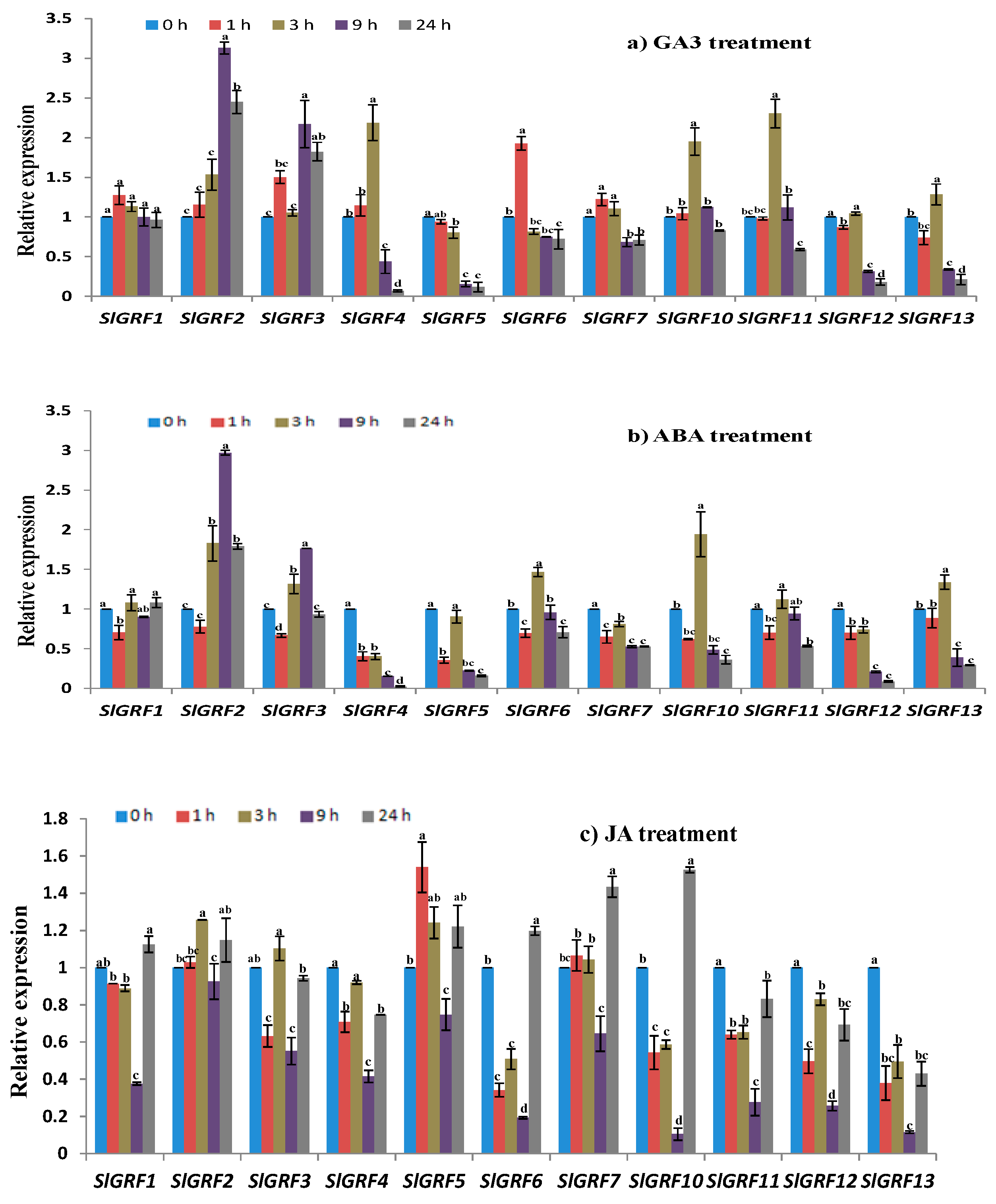
2.7.1. GA3 Treatment
2.7.2. Abscisic Acid Treatment
2.7.3. Jasmonic Acid Treatment
3. Discussion
4. Materials and Methods
4.1. Identification of GRF Family Genes and GRF Proteins in Tomato
4.2. Identification of Cis-Acting Elements and Chromosomal Position of Tomato GRF Genes
4.3. Functional Analysis of Tomato GRF Genes
4.4. Plant Sample Collection
4.5. RNA Extraction and cDNA Synthesis
4.6. qRT-PCR Expression Analysis
4.7. Statistical Analyses
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Kim, J.H.; Tsukaya, H. Regulation of plant growth and development by the GROWTH-REGULATING FACTOR and GRF-INTERACTING FACTOR duo. J. Exp. Bot. 2015, 66, 6093–6107. [Google Scholar] [CrossRef] [PubMed]
- Bao, M.; Bian, H.; Zha, Y.; Li, F.; Sun, Y.; Bai, B.; Chen, Z.; Wang, J.; Zhu, M.; Han, N. miR396a-mediated basic helix-loophelix transcription factor bHLH74 repression acts as a regulator for root growth in Arabidopsis seedlings. Plant Cell Physiol. 2014, 6, 1343–1353. [Google Scholar] [CrossRef] [PubMed]
- Debernardi, J.M.; Mecchia, M.A.; Vercruyssen, L.; Smaczniak, C.; Kaufmann, K.; Inze, D.; Rodriguez, R.E.; Palatnik, J.F. Post-transcriptional control of GRF transcription factors by microRNA miR396 and GIF co-activator affects leaf size and longevity. Plant J. 2014, 79, 413–426. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Guo, S.; Xu, Y.; Li, C.; Zhang, Z.; Zhang, D.; Xu, S.; Zhang, C.; Chong, K. OsmiR396d-regulated OsGRFs function in floral organogenesis in rice through binding to their targets OsJMJ706 and OsCR4. Plant Physiol. 2014, 165, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Omidbakhshfard, M.A.; Proost, S.; Fujikura, U.; Mueller-Roeber, B. Growth-regulating factors (GRFs): A small transcription factor family with important functions in plant biology. Mol. Plant. 2015, 8, 998–1010. [Google Scholar] [CrossRef] [PubMed]
- Van der Knaap, E.; Kim, J.H.; Kende, H. A novel gibberellins-induced gene from rice and its potential regulatory role in stem growth. Plant Physiol. 2000, 122, 695–704. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.F.; Li, B.; Jia, G.Q.; Zhang, T.F.; Dai, J.R.; Li, J.S.; Wang, S.C. Isolation and characterization of genes encoding GRF transcription factors and GIF transcriptional coactivators in Maize (Zea mays L.). Plant Sci. 2008, 175, 809–817. [Google Scholar] [CrossRef]
- Kim, J.H.; Choi, D.; Kende, H. The AtGRF family of putative transcription factors is involved in leaf and cotyledon growth in Arabidopsis. Plant J. 2003, 36, 94–104. [Google Scholar] [CrossRef] [PubMed]
- Treich, I.; Cairns, B.R.; de los Santos, T.; Brewster, E.; Carlson, M. SNF11, a new component of the yeast SNF-SWI complex that interacts with a conserved region of SNF2. Mol. Cell. Biol. 1995, 15, 4240–4248. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.H.; Kende, H. A transcriptional coactivator, AtGIF1, is involved in regulated leaf growth and morphology in Arabidopsis. Proc. Natl. Acad. Sci. USA 2004, 101, 13374–13379. [Google Scholar] [CrossRef] [PubMed]
- Choi, D.; Kim, J.H.; Kende, H. Whole genome analysis of the OsGRFgene family encoding plant-specific putative transcription activators in rice (Oryza sativa L.). Plant Cell Physiol. 2004, 45, 897–904. [Google Scholar] [CrossRef] [PubMed]
- Wang, F.; Qiu, N.; Ding, Q.; Li, J.; Zhang, Y.; Li, H.; Gao, J. Genome-wide identification and analysis of the growth-regulating factor family in Chinese cabbage (Brassica rapa L. ssp. pekinensis). BMC Genom. 2014, 15, 1. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.J.; Lee, B.H. Growth-regulating factor 4 of Arabidopsis thaliana is required for development of leaves, cotyledons, and shoot apical meristem. J. Plant Biol. 2006, 49, 463–468. [Google Scholar] [CrossRef]
- Yang, F.; Liang, G.; Liu, D.; Yu, D. Arabidopsis miR396 mediates the development of leaves and flowers in transgenic tobacco. J. Plant Biol. 2009, 52, 475–481. [Google Scholar] [CrossRef]
- Baucher, M.; Moussawi, J.; Vandeputte, O.M.; Monteyne, D.; Mol, A.; Pérez-Morga, D.; Jaziri, M.E.I. A role for the miR396/GRF network in specification of organ type during flower development, as supported by ectopic expression of Populus trichocarpa miR396c in transgenic tobacco. Plant Biol. 2013, 15, 892–898. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; He, H.; Li, Y.; Wang, F.; Yu, D. Molecular mechanism of miR396 mediating pistil development in Arabidopsis thaliana. Plant Physiol. 2014, 164, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Hua, W.; Yang, H.L.; Zhan, G.M.; Li, R.J.; Deng, L.B.; Wang, X.F.; Liu, G.H.; Wang, H.Z. The BnGRF2 gene (GRF2-like gene from Brassica napus) enhances seed oil production through regulating cell number and plant photosynthesis. J. Exp. Bot. 2012, 63, 3727–3740. [Google Scholar] [CrossRef] [PubMed]
- Wynn, A.N.; Rueschhoff, E.E.; Franks, R.G. Transcriptomic characterization of a synergistic genetic interaction during carpel margin meristem development in Arabidopsis thaliana. PLoS ONE 2011, 6, e26231. [Google Scholar] [CrossRef] [PubMed]
- Ye, R.; Yao, Q.H.; Xu, Z.H.; Xue, H.W. Development of an efficient method for the isolation of factors involved in gene transcription during rice embryo development. Plant J. 2004, 38, 348–357. [Google Scholar] [CrossRef] [PubMed]
- Kuijt, S.J.; Greco, R.; Agalou, A.; Shao, J.; CJ’t Hoen, C.O.; Verna, S.E.; Osnato, M.; Curiale, S.; Meynard, D.; van Gulik, R. Interaction between the growth-regulating factor and knotted1-like homeobox families of transcription factors. Plant Physiol. 2014, 164, 1952–1966. [Google Scholar] [CrossRef] [PubMed]
- Hewezi, T.; Maier, T.R.; Nettleton, D.; Baum, T.J. The Arabidopsis microRNA396-GRF1/GRF3 regulatory module acts as a developmental regulator in the reprogramming of root cells during cyst nematode infection. Plant Physiol. 2012, 159, 321-35. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Fu, F.; Zhang, H.; Song, F. Genome-wide systematic characterization of the bZIP transcriptional factor family in tomato (Solanum lycopersicum L.). BMC Genom. 2015, 16, 1. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Mizoi, J.; Kidokoro, S.; Maruyama, K.; Nakajima, J.; Nakashima, K.; Mitsuda, N.; Takiguchi, Y.; ohmeoTakagi, M.; Kondou, Y.; et al. Arabidopsis GROWTH-REGULATING FACTOR7 functions as a transcriptional repressor of abscisic acid and osmotic stress–responsive genes, including DREB2A. Plant Cell. 2012, 24, 3393–3405. [Google Scholar] [CrossRef] [PubMed]
- Cao, D.; Wang, J.; Ju, Z.; Liu, Q.; Li, S.; Tian, H.; Fu, D.; Zhu, H.; Luo, Y.; Zhu, B. Regulations on growth and development in tomato cotyledon, flower and fruit via destruction of miR396 with short tandem target mimic. Plant Sci. 2016, 247, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Horiguchi, G.; Kim, G.T.; Tsukaya, H. The transcription factor AtGRF5 and the transcription coactivator AN3 regulate cell proliferation in leaf primordia of Arabidopsis thaliana. Plant J. 2005, 43, 68–78. [Google Scholar] [CrossRef] [PubMed]
- Ahmadi, J.; Noormohammadi, N.; Ourang, S.F. Identification of conserved domains and motifs for GRF gene family. Genome 2014, 1271, 9999895. [Google Scholar]
- Koutinas, N.; Pepelyankov, G.; Lichev, V. Flower induction and flower bud development in apple and sweet cherry. Biotechnol. Biotechnol. Equip. 2010, 24, 1549–1558. [Google Scholar] [CrossRef]
- Luo, A.D.; Liu, l.; Tang, Z.S.; Bai, X.Q.; Cao, S.Y.; Chu, C.C. Down-regulation of OsGrF1 gene in rice rhd1 mutant results in reduced heading date. J. Integr. Plant Biol. 2005, 47, 745–752. [Google Scholar] [CrossRef]
- Karlova, R.; Chapman, N.; David, K.; Angenent, G.C.; Seymour, G.B.; de Maagd, R.A. Transcriptional control of fleshy fruit development and ripening. J. Exp. Bot. 2014, 65, 4527–4541. [Google Scholar] [CrossRef] [PubMed]
- Pesaresi, P.; Mizzotti, C.; Colombo, M.; Masiero, S. Genetic regulation and structural changes during tomato fruit development and ripening. Front. Plant Sci. 2014, 5, 92. [Google Scholar] [CrossRef] [PubMed]
- De Jong, M.; Mariani, C.; Vriezen, W.H. The role of auxin and gibberellin in tomato fruit set. J. Exp. Bot. 2009, 60, 1523–1532. [Google Scholar] [CrossRef] [PubMed]
- Le, D.T.; Nishiyama, R.; Watanabe, Y.; Vankova, R.; Tanaka, M.; Seki, M.; Tran, L.S.P. Identification and expression analysis of cytokinin metabolic genes in soybean under normal and drought conditions in relation to cytokinin levels. PLoS ONE 2012, 7, 324–325. [Google Scholar] [CrossRef] [PubMed]
- Walther, D.; Brunnemann, R.; Selbig, J. The regulatory code for transcriptional response diversity and its relation to genome structural properties in A. thaliana. PLoS Genet. 2007, 3, 216–229. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Quackenbush, J. TM4 Microarray software suite. Method Enzymol. 2006, 411, 134–193. [Google Scholar]
- Liu, X.; Guo, L.X.; Jin, L.F.; Liu, Y.Z.; Liu, T.; Fan, Y.H.; Peng, S.A. Identification and transcript profiles of citrus growth-regulating factor genes involved in the regulation of leaf and fruit development. Mol. Biol. Rep. 2016, 43, 1059–1067. [Google Scholar] [CrossRef] [PubMed]
- Heidel, A.J.; Clarke, J.D.; Antonovics, J.; Dong, X. Fitness costs of mutations affecting the systemic acquired resistance pathway in Arabidopsis thaliana. Genetics 2004, 168, 2197–2206. [Google Scholar] [CrossRef] [PubMed]
- Sakuma, Y.; Maruyama, K.; Osakabe, Y.; Qin, F.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Functional analysis of an Arabidopsis transcription factor, DREB2A, involved in drought-responsive gene expression. Plant Cell. 2006, 18, 1292–1309. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Tian, X.; Li, Y.; Wu, C.; Zheng, C. Microarray-based analysis of stress-regulated microRNAs in Arabidopsis thaliana. RNA 2008, 14, 836–843. [Google Scholar] [CrossRef] [PubMed]
- Casadevall, R.; Rodriguez, R.; Debernardi, J.; Palatnik, J.; Casati, P. Repression of growth regulating factors by the microRNA396 inhibits cell proliferation by UV-B radiation in Arabidopsis leaves. Plant Cell. 2013, 25, 3570–3583. [Google Scholar] [CrossRef] [PubMed]
- Casati, P. Analysis of UV-B regulated miRNAs and their targets in maize leaves. Plant Signal. Behav. 2013, 8, e26758. [Google Scholar] [CrossRef]
- Liu, J.; Rice, J.H.; Chen, N.; Baum, T.J.; Hewezi, T. Synchronization of developmental processes and defense signaling by growth regulating transcription factors. PLoS ONE 2014, 9, e98477. [Google Scholar] [CrossRef] [PubMed]
- Büyük, İ.; Aras, S. Genome-wide in silico identification, characterization and transcriptional analysis of the family of growth-regulating factors in common bean (Phaseolus vulgaris L.) subjected to PEG-induced drought stress. Arch. Biol. Sci. 2016, 69, 5–14. [Google Scholar] [CrossRef]
- Wolters, H.; Jurgens, G. Survival of the flexible: Hormonal growth control and adaptation in plant development. Nat. Rev. Genet. 2009, 10, 305–317. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.; Wu, W.; Abrams, S.R.; Cutler, A.J. The relationship of drought-related gene expression in Arabidopsis thaliana to hormonal and environmental factors. J. Expt. Bot. 2008, 59, 2991–3007. [Google Scholar] [CrossRef] [PubMed]
- Fujita, M.; Fujita, Y.; Noutoshi, Y.; Takahashi, F.; Narusaka, Y.; Yamaguchi-Shinozaki, K.; Shinozaki, K. Crosstalk between abiotic and biotic stress responses: A current view from the points of convergence in the stress signaling networks. Curr. Opin. Plant Biol. 2006, 9, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Singh, D.P.; Jermakow, A.M.; Swain, S.M. Gibberellins are required for seed development and pollen tube growth in Arabidopsis. Plant Cell. 2002, 14, 3133–3147. [Google Scholar] [CrossRef] [PubMed]
- Tuteja, N. Abscisic acid and abiotic stress signaling. Plant Signal. Behav. 2007, 2, 135–138. [Google Scholar] [CrossRef] [PubMed]
- Sakamoto, T.; Kamiya, N.; Ueguchi-Tanaka, M.; Iwahori, S.; Matsuoka, M. KNOX homeodomain protein directly suppresses the expression of a gibberellin biosynthetic gene in the tobacco shoot apical meristem. Genes Dev. 2001, 15, 581–590. [Google Scholar] [CrossRef] [PubMed]
- Browse, J. Jasmonate: Preventing the maize tassel from getting in touch with his feminine side. Sci. Signal. 2009, 2, pe9. [Google Scholar] [CrossRef] [PubMed]
- Moreno, J.E.; Tao, Y.; Chory, J.; Ballare, C.L. Ecological modulation of plant defense via phytochrome control of jasmonates sensitivity. Proc. Natl. Acad. Sci. USA 2009, 106, 106–4940. [Google Scholar] [CrossRef] [PubMed]
- Nafie, E.; Hathout, T.; Al, S.; Al, M. Jasmonic acid elicits oxidative defense and detoxification systems in Cucumis melo L. cells. Braz. J. Plant Physiol. 2011, 23, 161–174. [Google Scholar] [CrossRef]
- Cipollini, D. Constitutive expression of methyl jasmonates inducible responses delays reproduction and constrains fitness responses to nutrients in Arabidopsis thaliana. Evol. Ecol. 2010, 24, 59–68. [Google Scholar] [CrossRef]
- Solgenomics database. Available online: https://solgenomics.net/ (accessed on 24 August 2017).
- Locus in Solgenomics database. Available online: https://solgenomics.net/search/locus (accessed on 30 August 2017).
- iTAK-Plant Transcription factor and Protein Kinase Identifier and Classifier. Available online: http://bioinfo.bti.cornell.edu/cgi-bin/itak/index.cgi (accessed on 01 September 2017).
- Tomato genomic Resources database. Available online: http://59.163.192.91/tomato2/getTF_family.php?trans_fac_family=zf-HD (accessed on 01 September 2017).
- SMART conserved domain search tool. Available online: http://smart.embl-heidelberg.de/ (accessed on 01 September 2017).
- Pfam database. Available online: http://www.sanger.ac.uk/science/tools/pfam (accessed on 05 September 2017).
- NCBI database. Available online: https://www.ncbi.nlm.nih.gov/orffinder/ (accessed on 05 September 2017).
- ProtParam tool. Available online: http://web.expasy.org/protparam/ (accessed on 30 August 2017).
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [PubMed]
- The Gene Structure display Server. Available online: http://gsds.cbi.pku.edu.cn/ (accessed on 30 August 2017).
- Genedoc multiple sequence alignment tool. Available online: https://www.nrbsc.org/gfx/genedoc/ebinet.htm (accessed on 30 August 2017).
- Multiple EM for Motif Elicitation (MEME) web tool. Available online: http://meme-suite.org/ (accessed on 30 August 2017).
- PlantCARE database. Available online: http://bioinformatics.psb.ugent.be/webtools/plantcare/html/ (accessed on 10 September 2017).
- MapGene2Chrom web v2 software. Available online: http://mg2c.iask.in/ (accessed on 10 September 2017).
- Blast2GO functional annotation and genomics software. Available online: https://www.blast2go.com/ (accessed on 15 October 2017).
- Khatun, K.; Robin, A.H.K.; Park, J.I.; Kim, C.K.; Kim, K.B.; Lim, M.B.; Lee, D.J.; Nou, I.S.; Chung, M.Y. Genome-wide identification, characterization and expression profiling of ADF family genes in Solanum lycopersicum L. Genes 2016, 7, 79. [Google Scholar] [CrossRef] [PubMed]
- Khatun, K.; Robin, A.H.K.; Park, J.I.; Ahmed, N.U.; Kim, C.K.; Lim, K.B.; Kim, M.B.; Lee, D.J.; Nou, I.S.; Chung, M.Y. Genome-wide identification, characterization and expression profiling of LIM family genes in Solanum lycopersicum L. Plant Physiol. Biochem. 2016, 108, 177–190. [Google Scholar] [CrossRef] [PubMed]
- Primer3 software. Available online: http://bioinfo.ut.ee/primer3-0.4.0/primer3/input.htm (accessed on 01 September 2017).
- Robin, A.H.K.; Yi, G.E.; Laila, R.; Yang, K.; Park, J.I.; Kim, H.R.; Nou, I.S. Expression profiling of glucosinolate biosynthetic genes in Brassica oleracea L. var. capitata inbred lines reveals their association with glucosinolate content. Molecules 2016, 21, 787. [Google Scholar] [CrossRef]
- Aoki, K.; Yano, K.; Suzuki, A.; Kawamura, S.; Sakurai, N.; Suda, K.; Ooga, K. Large-scale analysis of full length cDNAs from the tomato (Solanum lycopersicum) cultivar Micro-Tom, a reference system for the Solanaceae genomics. BMC Genom. 2010, 11, 210. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative CT method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Serial No. | Gene Name | Locus Name | ORF (bp) | Chrom. (strand) | No. of Introns | Protein | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Length (aa) | Domain (Start–End) | MW (kDa) | pI | |||||||
| QLQ | WRC | |||||||||
| 1 | SlGRF1 | Solyc12g096070 | 1038 | C12(+) | 3 | 345 | 9–45 | 72–114 | 38.5907 | 7.13 |
| 2 | SlGRF2 | Solyc08g005430 | 1071 | C08(+) | 2 | 356 | 30–66 | 93–135 | 39.1162 | 8.63 |
| 3 | SlGRF3 | Solyc08g075950 | 1086 | C08(−) | 2 | 361 | 27–63 | 90–132 | 39.8591 | 8.43 |
| 4 | SlGRF4 | Solyc07g041640 | 1035 | C07(−) | 2 | 344 | 10–46 | 79–121 | 39.59 | 8.99 |
| 5 | SlGRF5 | Solyc04g077510 | 1788 | C04(+) | 3 | 595 | 136–172 | 205–247 | 64.1118 | 8.3 |
| 6 | SlGRF6 | Solyc02g092070 | 1704 | C02(+) | 4 | 568 | 139–175 | 210–252 | 61.3674 | 8.51 |
| 7 | SlGRF7 | Solyc08g083230 | 1197 | C08(+) | 3 | 398 | 74–108 | 125–167 | 43.7076 | 7.92 |
| 8 | SlGRF8 | Solyc03g082430 | 1377 | C03(−) | 3 | 458 | 58–94 | 117–159 | 50.4247 | 5.97 |
| 9 | SlGRF9 | Solyc08g068760 | 468 | C08(+) | 0 | 155 | 46–82 | 106–143 | 17.3958 | 8.84 |
| 10 | SlGRF10 | Solyc01g091540 | 1263 | C01(+) | 2 | 420 | 57–93 | 122–163 311–352 | 46.5821 | 9.18 |
| 11 | SlGRF11 | Solyc09g009200 | 1146 | C09(−) | 3 | 381 | 79–115 | 150–192 | 42.0369 | 9.08 |
| 12 | SlGRF12 | Solyc10g083510 | 1170 | C10(−) | 3 | 389 | 81–115 | 157–199 | 42.3790 | 9.18 |
| 13 | SlGRF13 | Solyc08g079800 | 657 | C08(−) | 3 | 218 | 49–83 | 112–154 | 24.7856 | 8.95 |
| Observation on | Key Findings and Predicted Specific Function (s) Based on Expression Data | |
|---|---|---|
| Structural characteristics | 13 GRF genes were identified from Solanum lycopersicum. All of them contained functional QLQ and WRC domain and diverge C-terminal region rich in Pro, Gln, Ser/Thr that are frequently found in transcription factors. Besides, some of SlGRF contained C-terminal FFD and TQL motifs. The structural characteristics suggested that SlGRF proteins function as transcriptional regulators. | |
| Phylogenetic classification | The GRF proteins from tomato, potato, Arabidopsis, Chinese cabbage, rice and maize phylogenitically classified into nine subfamilies which indicated their ancestral evolution and tomato GRF proteins are more closely related to potato suggested their evolution from common ancestor. | |
| Relative expression in different organs/tissues | Among the 13 SlGRF genes one gene; SlGRF9 was undetectable in all organ studied and suggested as pseudogene. The organ expression analysis revealed that most of the genes predominantly expressed in flower bud indicating possible function in flower bud (i.e., reproductive organ development) in tomato. The SlGRF genes also showed differential expression in meristem, full blooming flower, leaf, stem, root, seedling and in different fruit developmental stages suggesting the important function in growth and development of tomato. The relatively higher expression of 12 SlGRF genes in different organs are listed below: | |
| SlGRF1: | Flower bud, meristem and ripening fruit | |
| SlGRF2: | Flower bud, meristem, flower blooming and small green fruit | |
| SlGRF3: | Flower bud, meristem, flower blooming and small green fruit | |
| SlGRF4: | Flower bud | |
| SlGRF5: | Flower bud, meristem and small green fruit | |
| SlGRF7: | Flower bud and meristem | |
| SlGRF8: | Flower bud | |
| SlGRF10: | Flower bud, meristem and ripening fruit | |
| SlGRF11: | Flower bud and meristem | |
| SlGRF12: | Meristem, leaf, flower bud, small green fruit | |
| SlGRF13: | Meristem, leaf, flower bud, seedling, stem, and small green fruit | |
| Relative expression under abiotic stresses and phytohormone treatments | Four abiotic stresses—NaCl, drought, heat, cold and three phytohormones (GA3, ABA, and JA) treatments—were studied where the following genes were (up/down) regulated by abiotic and phytohormone treatments at different time points: | |
| SlGRF1: | NaCl, drought, heat, cold, JA | |
| SlGRF2: | NaCl, drought, heat, cold, ABA, GA3, JA | |
| SlGRF3: | NaCl, drought, heat, cold, ABA, GA3, JA | |
| SlGRF4: | NaCl, drought, heat, cold, ABA, GA3, JA | |
| SlGRF5: | NaCl, drought, heat, cold, ABA, GA3 | |
| SlGRF6: | NaCl, drought, heat, cold, ABA, GA3, JA | |
| SlGRF7: | NaCl, drought, ABA, GA3, JA | |
| SlGRF10: | Drought, heat, cold, ABA, JA | |
| SlGRF11: | Drought, heat, cold, JA | |
| SlGRF12: | Drought, heat, cold, ABA, JA | |
| SlGRF13: | Drought, heat, cold, ABA, JA | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khatun, K.; Robin, A.H.K.; Park, J.-I.; Nath, U.K.; Kim, C.K.; Lim, K.-B.; Nou, I.S.; Chung, M.-Y. Molecular Characterization and Expression Profiling of Tomato GRF Transcription Factor Family Genes in Response to Abiotic Stresses and Phytohormones. Int. J. Mol. Sci. 2017, 18, 1056. https://doi.org/10.3390/ijms18051056
Khatun K, Robin AHK, Park J-I, Nath UK, Kim CK, Lim K-B, Nou IS, Chung M-Y. Molecular Characterization and Expression Profiling of Tomato GRF Transcription Factor Family Genes in Response to Abiotic Stresses and Phytohormones. International Journal of Molecular Sciences. 2017; 18(5):1056. https://doi.org/10.3390/ijms18051056
Chicago/Turabian StyleKhatun, Khadiza, Arif Hasan Khan Robin, Jong-In Park, Ujjal Kumar Nath, Chang Kil Kim, Ki-Byung Lim, Ill Sup Nou, and Mi-Young Chung. 2017. "Molecular Characterization and Expression Profiling of Tomato GRF Transcription Factor Family Genes in Response to Abiotic Stresses and Phytohormones" International Journal of Molecular Sciences 18, no. 5: 1056. https://doi.org/10.3390/ijms18051056