
Emergence of CD26+ Cancer Stem Cells with Metastatic Properties in Colorectal Carcinogenesis

Abstract
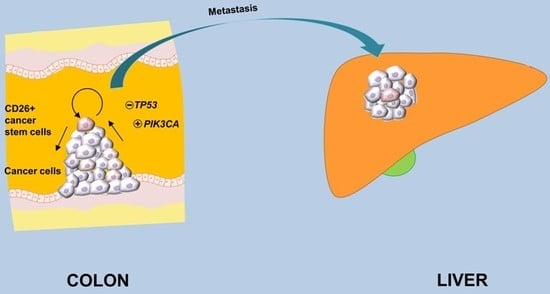
:
1. Introduction
2. Results
2.1. Primary Tumors with Confirmed or Suspected Metastases Harbor a Relatively High Proportion of CD26+ Cancer Stem Cell Subpopulation
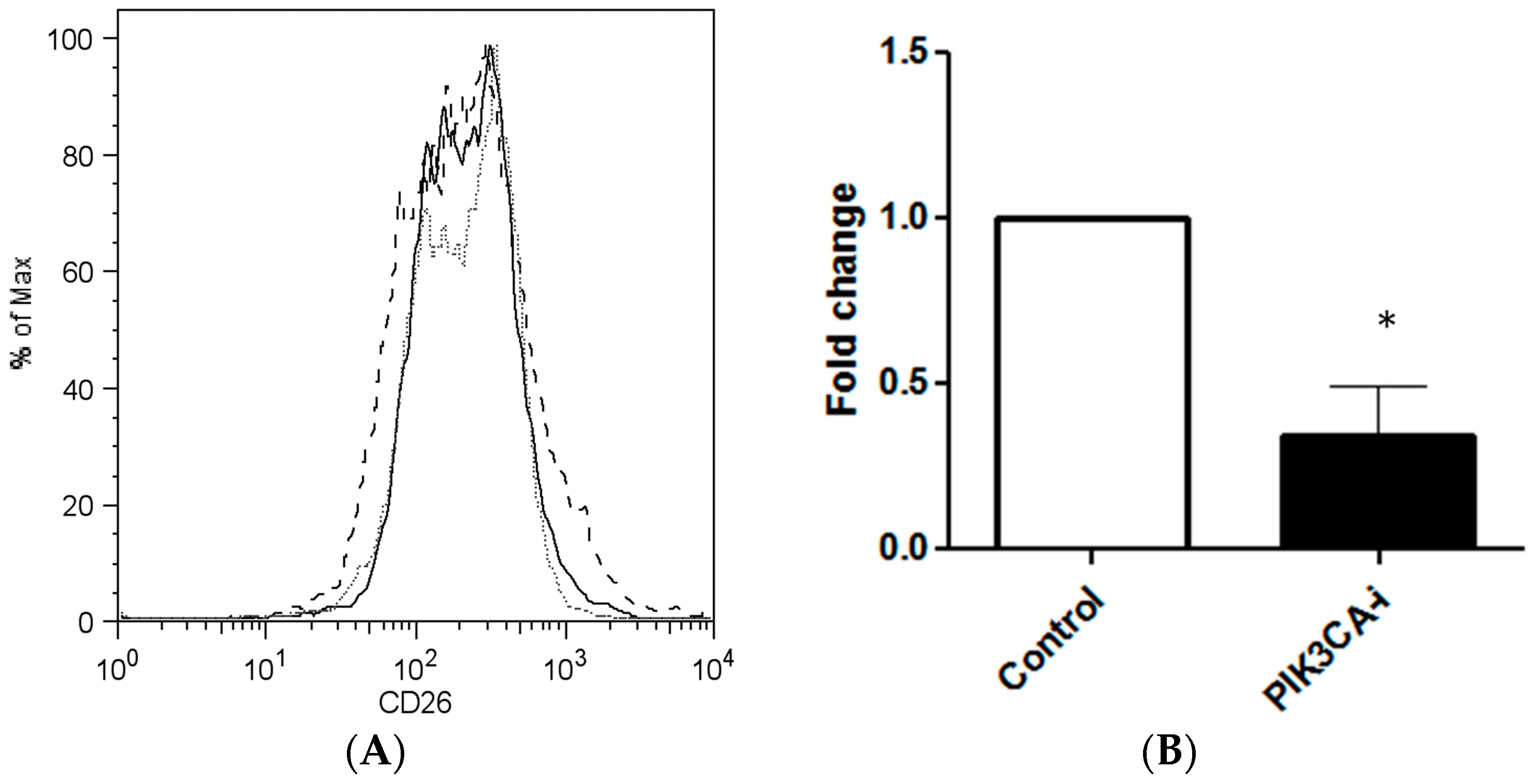
2.2. PIK3CA Inhibitor Decreases the Maintenance of CD26+ Cell Population in Culture
2.3. TP53 Knockdown Increases the Maintenance of CD26+ Cell Population in Culture
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Flow Cytometry and Fluorescent-Activated Cell Sorting (FACS)
4.3. PIK3CA Inhibitor Assay
4.4. Knockdown of P53
4.5. Clinical Specimens
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Conflicts of Interest
References
- Clevers, H. The cancer stem cell: Premises, promises and challenges. Nat. Med. 2011, 17, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Pang, R.; Law, W.L.; Chu, A.C.; Poon, J.T.; Lam, C.S.; Chow, A.K.; Ng, L.; Cheung, L.W.; Lan, X.R.; Lan, H.Y.; et al. A subpopulation of CD26+ cancer stem cells with metastatic capacity in human colorectal cancer. Cell Stem Cell 2010, 6, 603–615. [Google Scholar] [CrossRef] [PubMed]
- Chow, A.K.; Cheng, N.S.; Lam, C.S.; Wong, K.M.; Wan, M.H.; Man, H.W.; Cheung, H.K.; Yau, C.C.; Poon, T.C.; Law, W.-L.; et al. Preclinical analysis of the anti-tumor and anti-metastatic effects of Raf265 on colon cancer cells and CD26+ cancer stem cells in colorectal carcinoma. Mol. Cancer 2015, 14, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Lam, C.S.-C.; Cheung, A.H.-K.; Wong, S.K.-M.; Wan, T.M.-H.; Ng, L.; Chow, A.K.-M.; Cheng, N.S.-M.; Pak, R.C.-H.; Li, H.-S.; Man, J.H.-W.; et al. Prognostic significance of CD26 in patients with colorectal cancer. PLoS ONE 2014, 9, e98582. [Google Scholar] [CrossRef] [PubMed]
- Jones, S.; Chen, W.-D.; Parmigiani, G.; Diehl, F.; Beerenwinkel, N.; Antal, T.; Traulsen, A.; Nowak, M.A.; Siegel, C.; Velculescu, V.E.; et al. Comparative lesion sequencing provides insights into tumor evolution. Proc. Natl. Acad. Sci. USA 2008, 105, 4283–4288. [Google Scholar] [CrossRef] [PubMed]
- Bosman, F.T.; Carneiro, F.; Hruban, R.H.; Theise, N.D. WHO Classification of Tumours of the Digestive System; International Agency for Research on Cancer (IARC): Lyon, France, 2010. [Google Scholar]
- Ogino, S.; Nosho, K.; Kirkner, G.J.; Shima, K.; Irahara, N.; Kure, S.; Chan, A.T.; Engelman, J.A.; Kraft, P.; Cantley, L.C.; et al. PIK3CA mutation is associated with poor prognosis among patients with curatively resected colon cancer. J. Clin. Oncol. 2009, 27, 1477–1484. [Google Scholar] [CrossRef] [PubMed]
- Grady, W.M.; Carethers, J.M. Genomic and epigenetic instability in colorectal cancer pathogenesis. Gastroenterology 2008, 135, 1079–1099. [Google Scholar] [CrossRef] [PubMed]
- Reya, T.; Morrison, S.J.; Clarke, M.F.; Weissman, I.L. Stem cells, cancer, and cancer stem cells. Nature 2001, 414, 105–111. [Google Scholar] [CrossRef] [PubMed]
- Ricci-Vitiani, L.; Lombardi, D.G.; Pilozzi, E.; Biffoni, M.; Todaro, M.; Peschle, C.; De Maria, R. Identification and expansion of human colon-cancer-initiating cells. Nature 2007, 445, 111–115. [Google Scholar] [CrossRef] [PubMed]
- Diehn, M.; Majeti, R. Metastatic cancer stem cells: An opportunity for improving cancer treatment? Cell Stem Cell 2010, 6, 502–503. [Google Scholar] [CrossRef] [PubMed]
- Pisco, A.; Huang, S. Non-genetic cancer cell plasticity and therapy-induced stemness in tumour relapse: ‘What does not kill me strengthens me’. Br. J. Cancer 2015, 112, 1725–1732. [Google Scholar] [CrossRef] [PubMed]
- Chaffer, C.L.; Brueckmann, I.; Scheel, C.; Kaestli, A.J.; Wiggins, P.A.; Rodrigues, L.O.; Brooks, M.; Reinhardt, F.; Su, Y.; Polyak, K.; et al. Normal and neoplastic nonstem cells can spontaneously convert to a stem-like state. Proc. Natl. Acad. Sci. USA 2011, 108, 7950–7955. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Quan, Y.; Wang, W.; Fu, Q.; Wu, J.; Mei, T.; Li, J.; Tang, Y.; Luo, C.; Ouyang, Q.; et al. Dynamic equilibrium between cancer stem cells and non-stem cancer cells in human SW620 and MCF-7 cancer cell populations. Br. J. Cancer 2012, 106, 1512–1519. [Google Scholar] [CrossRef] [PubMed]
- Cho, K.R.; Vogelstein, B. Genetic alterations in the adenoma–carcinoma sequence. Cancer 1992, 70, 1727–1731. [Google Scholar] [CrossRef]
- Leslie, A.; Carey, F.; Pratt, N.; Steele, R. The colorectal adenoma–carcinoma sequence. Br. J. Surg. 2002, 89, 845–860. [Google Scholar] [CrossRef]
- Fearon, E.R.; Vogelstein, B. A genetic model for colorectal tumorigenesis. Cell 1990, 61, 759–767. [Google Scholar] [CrossRef]
- Subarsky, P.; Hill, R.P. The hypoxic tumour microenvironment and metastatic progression. Clin. Exp. Metastasis 2003, 20, 237–250. [Google Scholar] [CrossRef]
- Mantovani, A. Cancer: Inflaming metastasis. Nature 2009, 457, 36–37. [Google Scholar] [CrossRef] [PubMed]
- Dallas, N.A.; Xia, L.; Fan, F.; Gray, M.J.; Gaur, P.; van Buren, G.; Samuel, S.; Kim, M.P.; Lim, S.J.; Ellis, L.M. Chemoresistant colorectal cancer cells, the cancer stem cell phenotype, and increased sensitivity to insulin-like growth factor-I receptor inhibition. Cancer Res. 2009, 69, 1951–1957. [Google Scholar] [CrossRef] [PubMed]
- Bitarte, N.; Bandres, E.; Boni, V.; Zarate, R.; Rodriguez, J.; Gonzalez-Huarriz, M.; Lopez, I.; Javier Sola, J.; Alonso, M.M.; Fortes, P.; et al. MicroRNA-451 is involved in the self-renewal, tumorigenicity, and chemoresistance of colorectal cancer stem cells. Stem Cells 2011, 29, 1661–1671. [Google Scholar] [CrossRef] [PubMed]
- Abdullah, L.N.; Chow, E.K.-H. Mechanisms of chemoresistance in cancer stem cells. Clin. Trans. Med. 2013, 2, 1. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Patient | Gender and Age | CD26+ Cells % | Stage at Surgery | Tumour Size (cm) | Degree of Differentiation | Metastasis Diagnosed or Suspected | Survival Status | Metastasis-Free Survival (Months) | Overall Survival (Months) |
|---|---|---|---|---|---|---|---|---|---|
| 1 | M/81 | 0.2 | III | 9 | Moderate | Liver | Deceased | 5.40 | 9.70 |
| 2 | F/80 | 0.3 | I | 5 | Moderate | None | Alive | 39.23 | 39.23 |
| 3 | F/68 | 0.4 | IIA | 10 | Moderate | None | Alive | 36.20 | 36.20 |
| 4 | M/71 | 0.5 | IIIC | 5 | Moderate | Peritoneum and bone | Deceased | 13.97 | 19.83 |
| 5 | M/79 | 0.6 | IIA | 11 | Poor | None | Alive | 33.17 | 33.17 |
| 6 | M/83 | 3.3 | IV | 12 | Moderate | Lung | Deceased | NA | 19.47 |
| 7 | M/74 | 8.2 | IV | 5 | Poor | Liver | Deceased | NA | 51.97 |
| 8 | F/56 | 9.6 | IV | 5.5 | Moderate | Liver | Deceased | NA | 29.67 |
| 9 | M/68 | 9.9 | IV | 9 | Moderate | Lung and liver | Alive | NA | 47.50 |
| 10 | F/84 | 12.7 | IIA | 10 | Moderate | Suspected lung metastasis | Alive | 21.30 | 31.23 |
| 11 | M/78 | 13.2 | IIA | 15 | Moderate | Liver | Deceased | 5.10 | 5.17 |
| Number of Patients | Confirmed or Suspected Metastases | ||
|---|---|---|---|
| Absent | Present | ||
| CD26+ cell proportion | low | 3 | 2 |
| high | 0 | 6 | |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cheung, A.H.-K.; Iyer, D.N.; Lam, C.S.-C.; Ng, L.; Wong, S.K.M.; Lee, H.-S.; Wan, T.; Man, J.; Chow, A.K.M.; Poon, R.T.; et al. Emergence of CD26+ Cancer Stem Cells with Metastatic Properties in Colorectal Carcinogenesis. Int. J. Mol. Sci. 2017, 18, 1106. https://doi.org/10.3390/ijms18061106
Cheung AH-K, Iyer DN, Lam CS-C, Ng L, Wong SKM, Lee H-S, Wan T, Man J, Chow AKM, Poon RT, et al. Emergence of CD26+ Cancer Stem Cells with Metastatic Properties in Colorectal Carcinogenesis. International Journal of Molecular Sciences. 2017; 18(6):1106. https://doi.org/10.3390/ijms18061106
Chicago/Turabian StyleCheung, Alvin Ho-Kwan, Deepak Narayanan Iyer, Colin Siu-Chi Lam, Lui Ng, Sunny K. M. Wong, Hung-Sing Lee, Timothy Wan, Johnny Man, Ariel K. M. Chow, Ronnie T. Poon, and et al. 2017. "Emergence of CD26+ Cancer Stem Cells with Metastatic Properties in Colorectal Carcinogenesis" International Journal of Molecular Sciences 18, no. 6: 1106. https://doi.org/10.3390/ijms18061106