In-Depth Proteomic Analysis of the Hippocampus in a Rat Model after Cerebral Ischaemic Injury and Repair by Danhong Injection (DHI)

Abstract
:
1. Introduction
2. Results
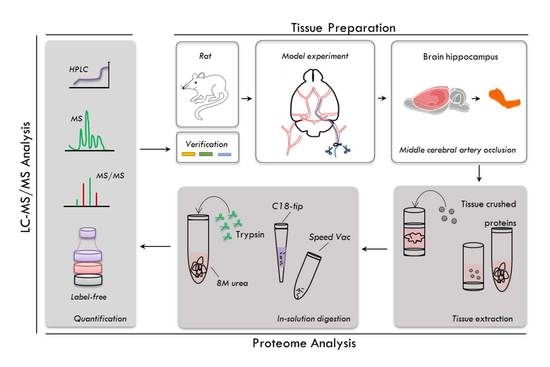
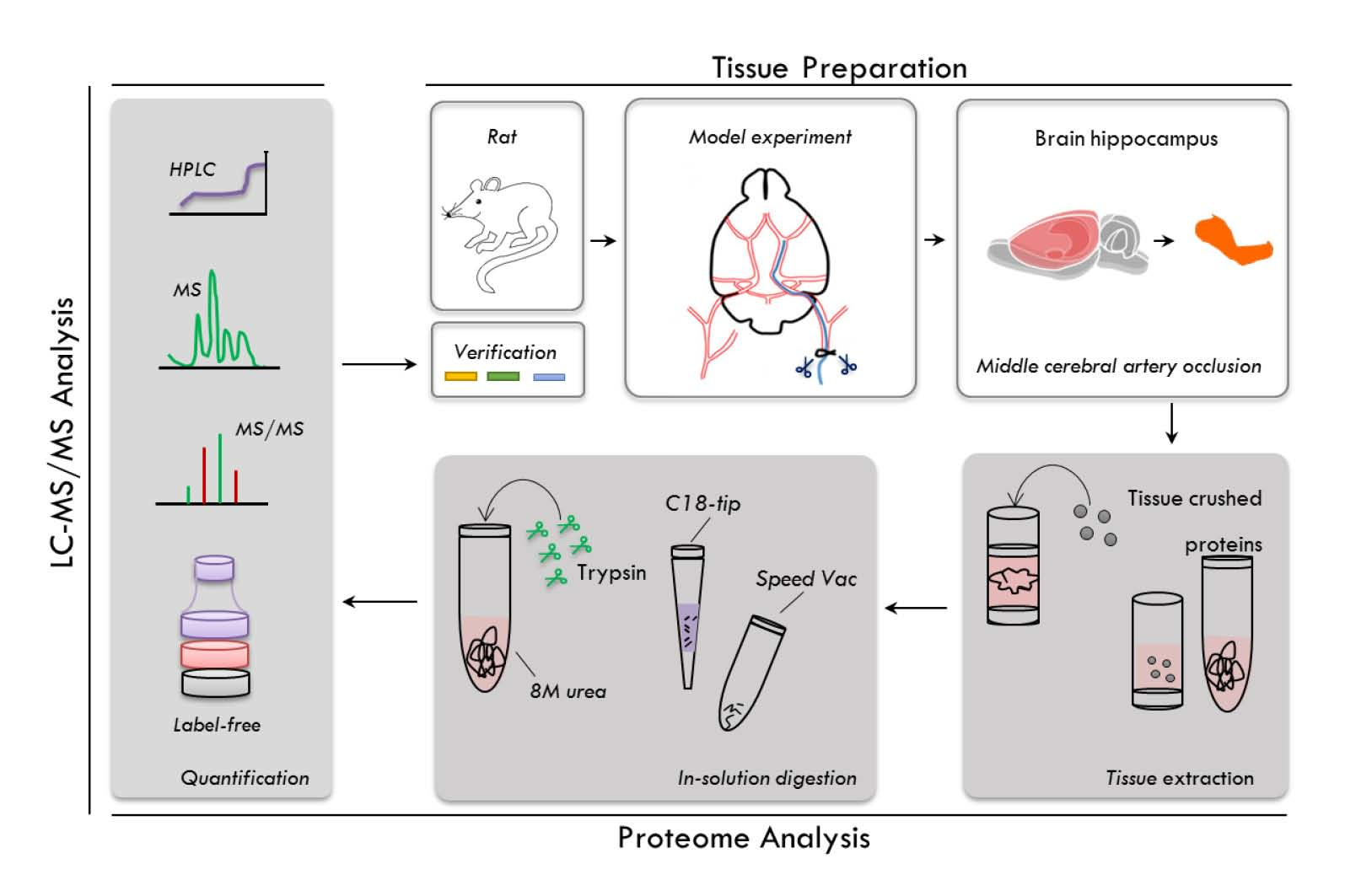
2.1. Experimental Workflow
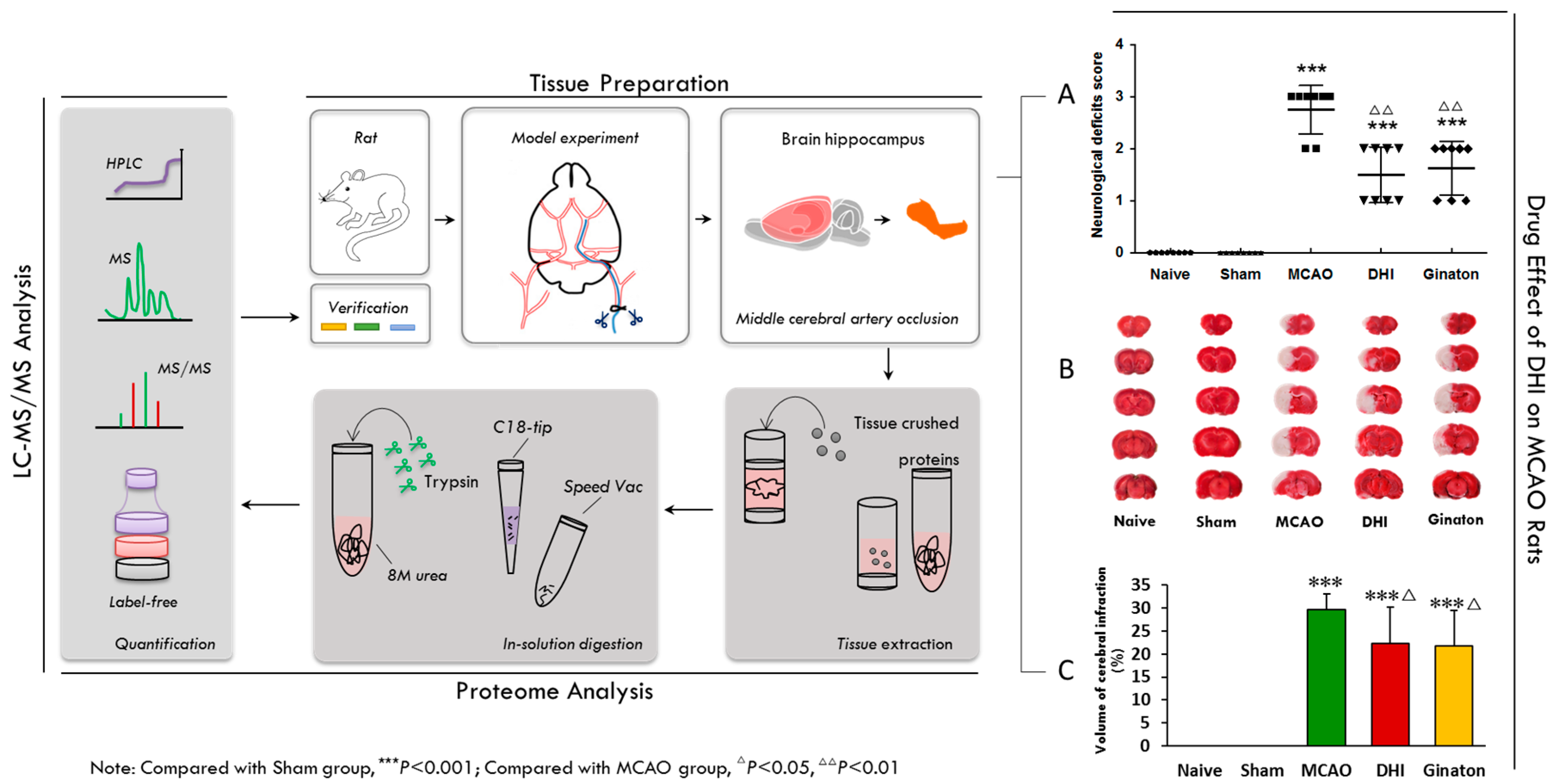
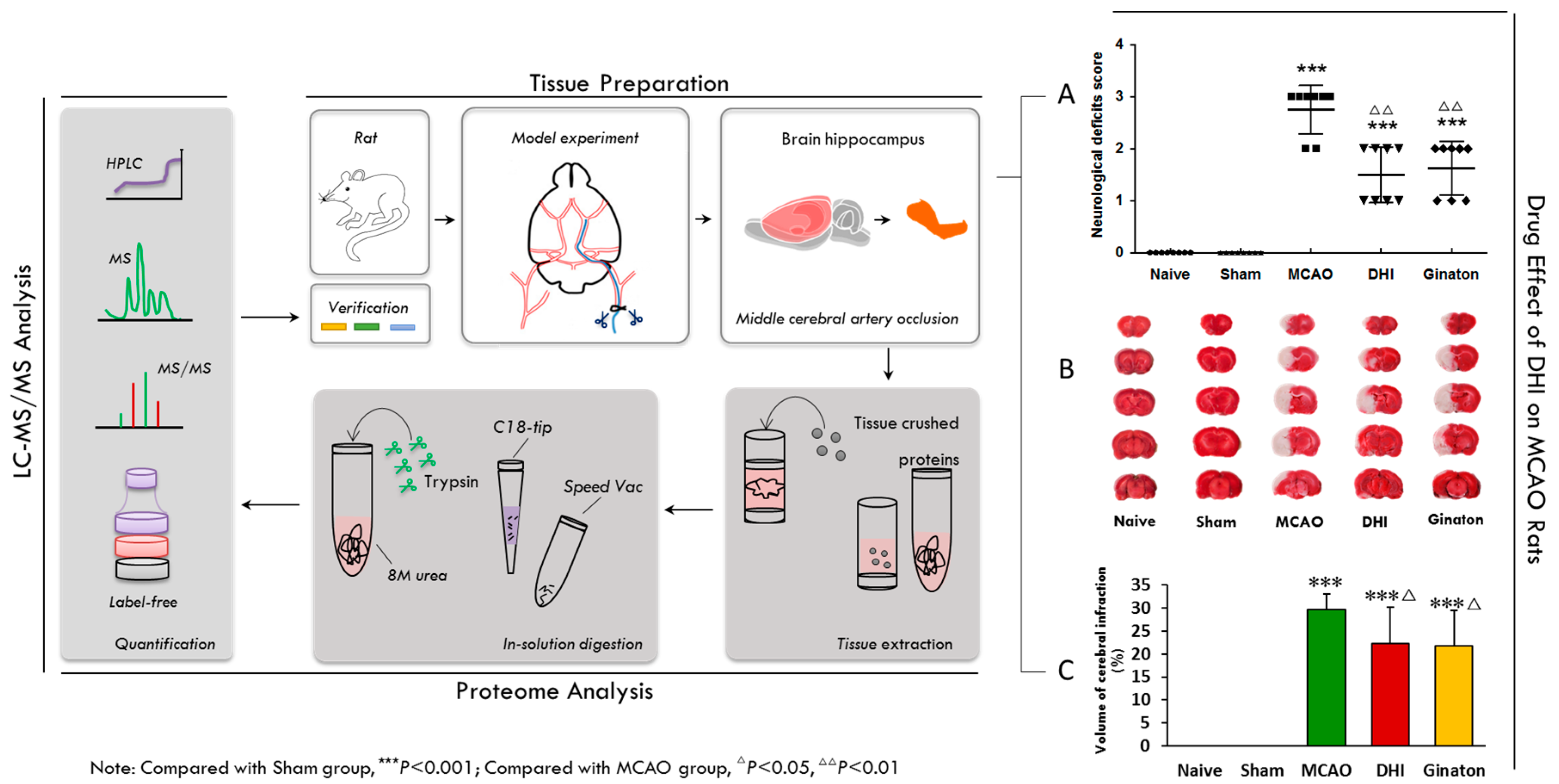
2.2. Evaluation of the Pharmacological Effects of DHI on MCAO Rats
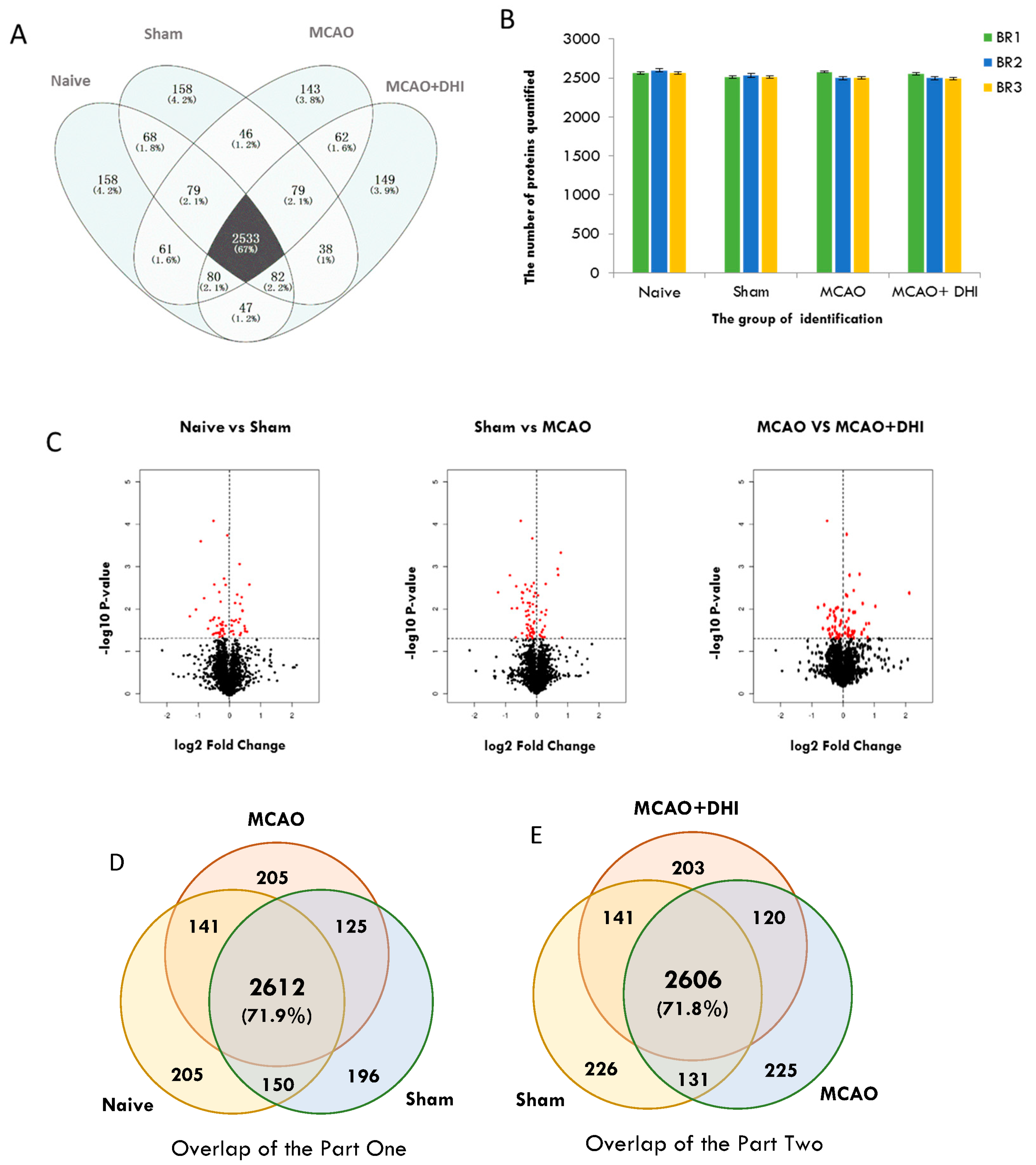
2.3. Proteome Profiling of the Hippocampus in Four Groups of Rats
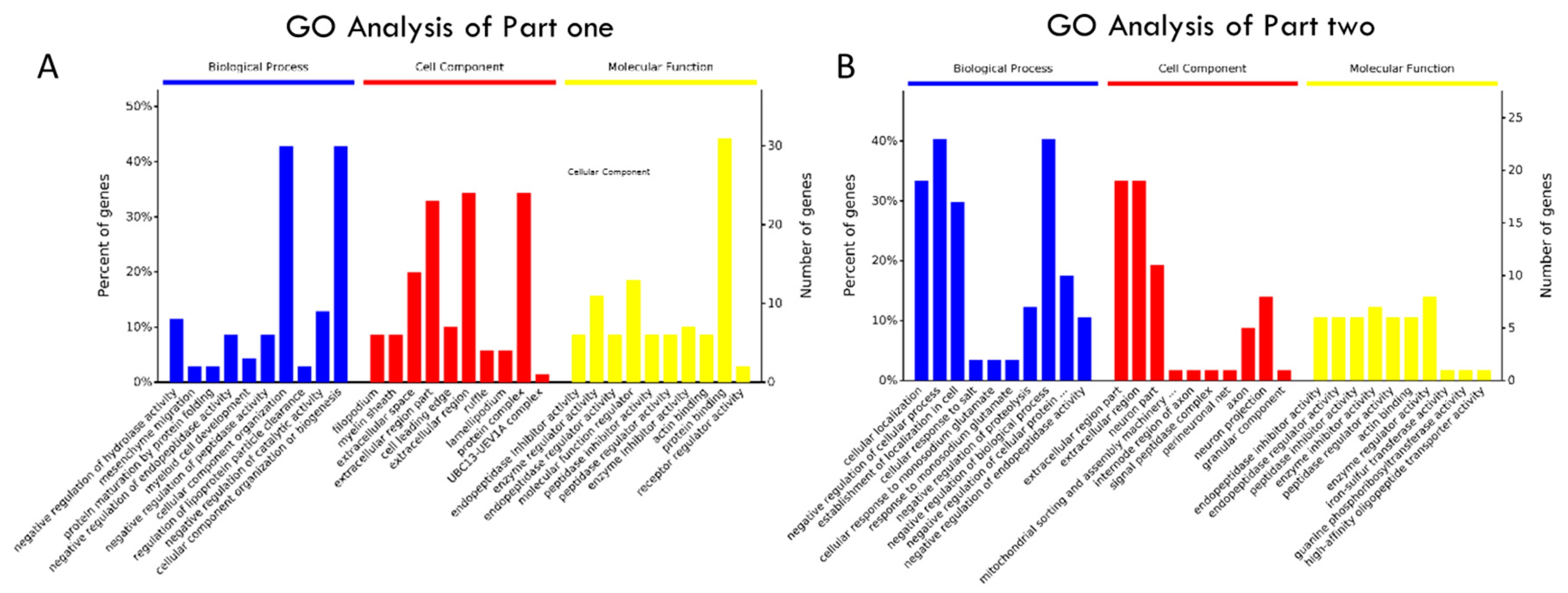
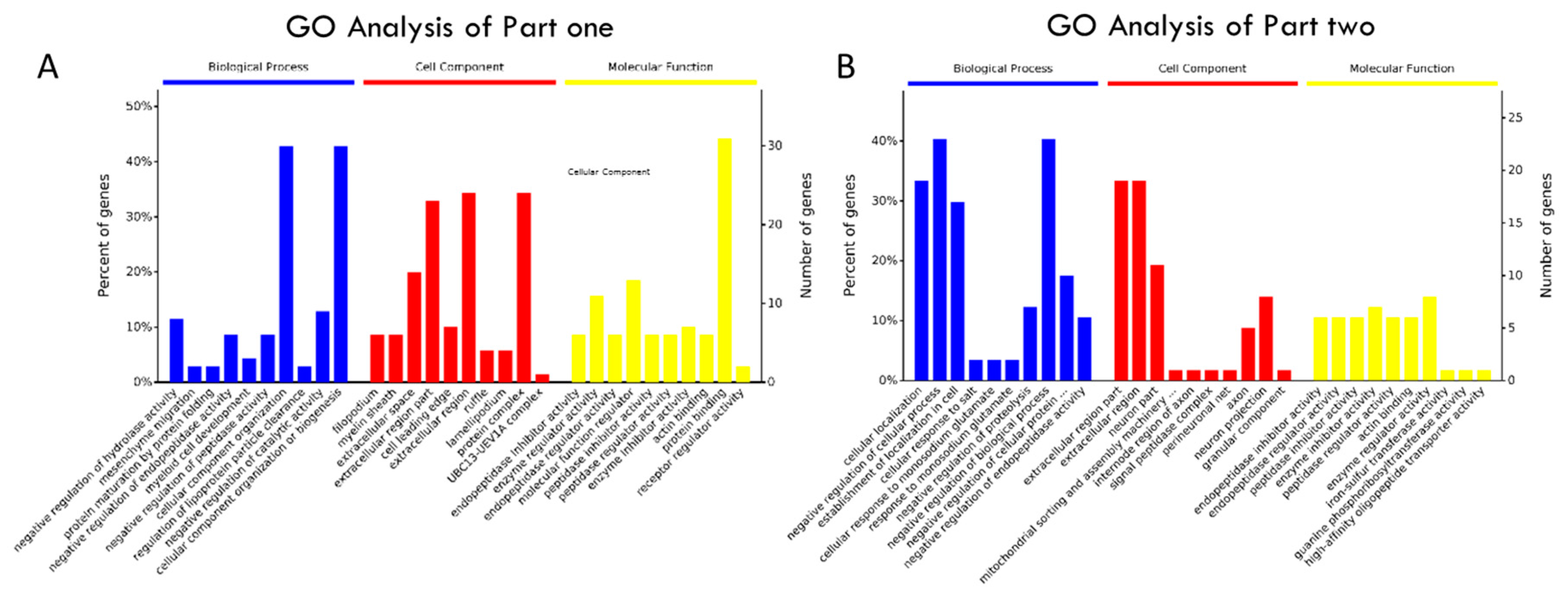
2.4. Annotation Map and Profiling of the Proteome
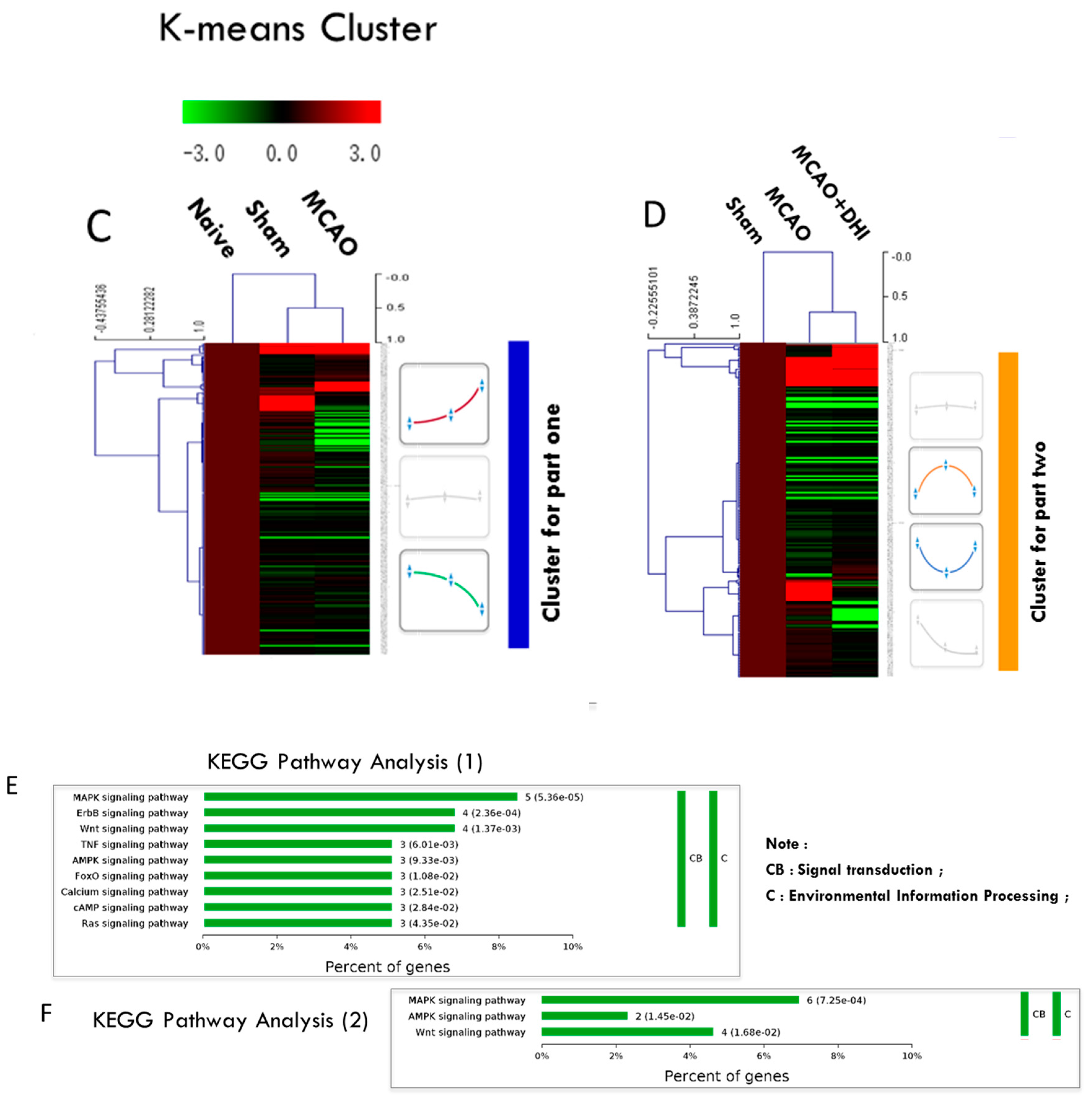
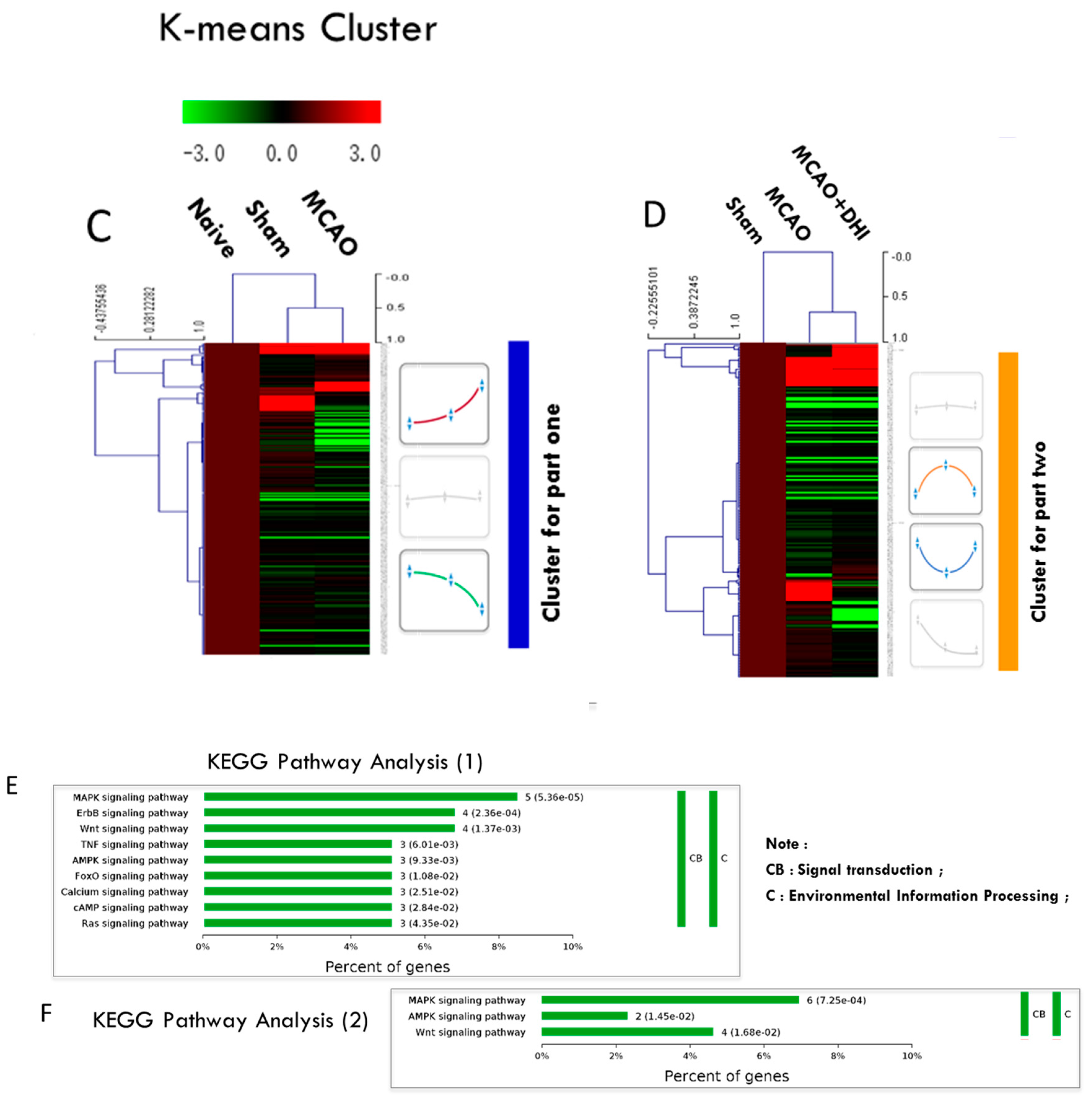
2.5. Hierarchical Clustering of the Significantly Altered Proteins
2.6. Pathway Enrichment of the Significantly Altered Proteins
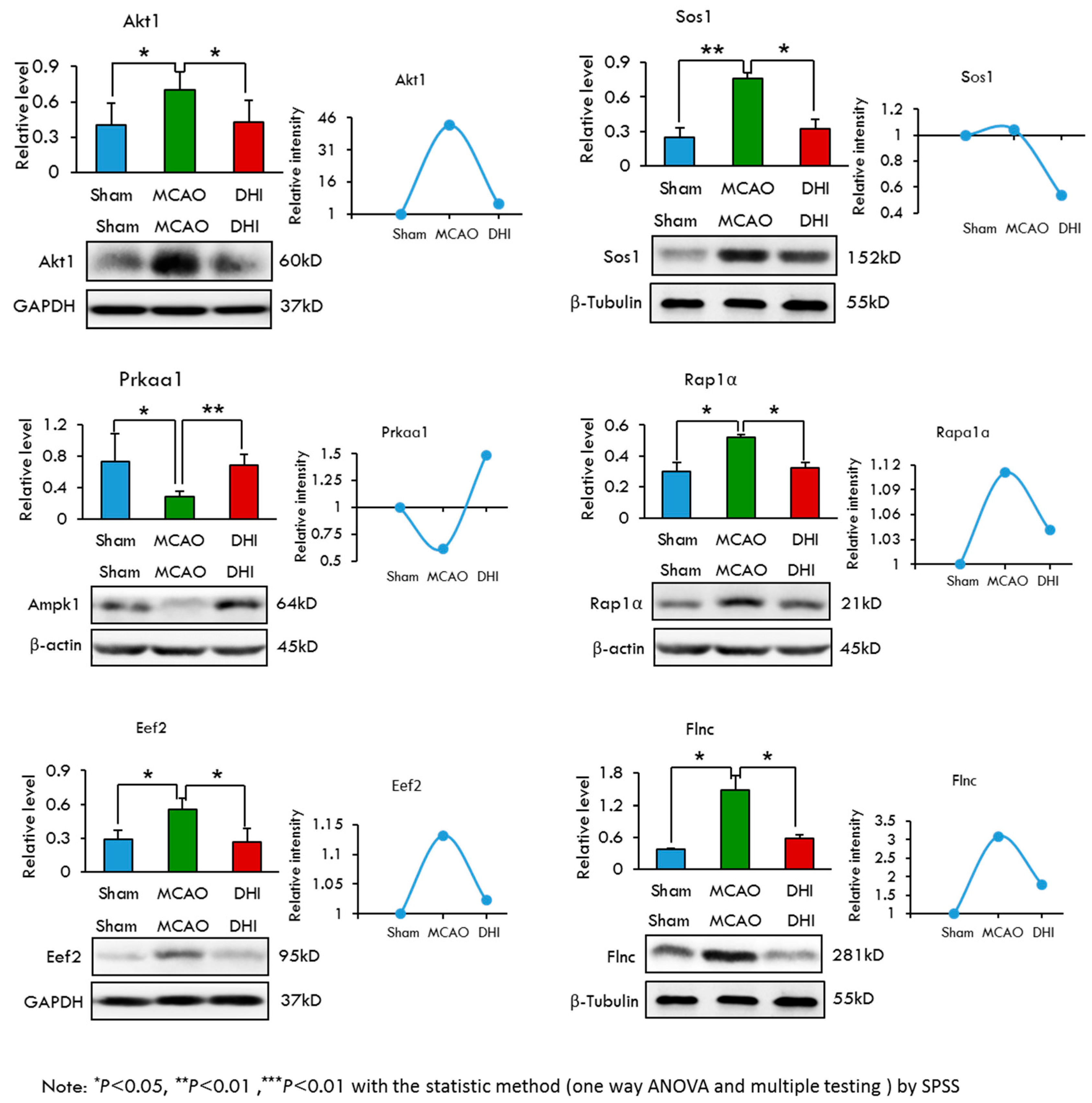
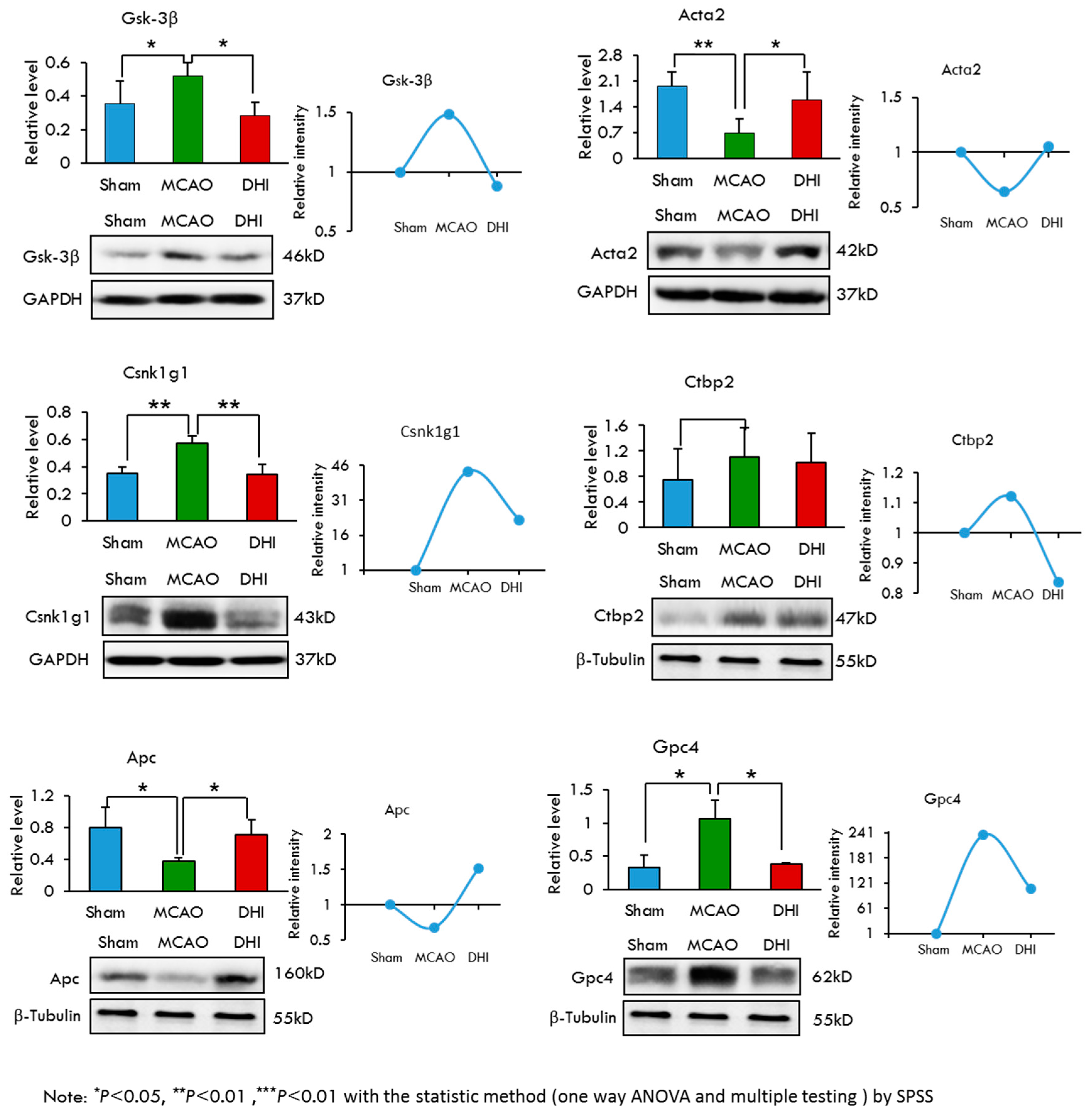
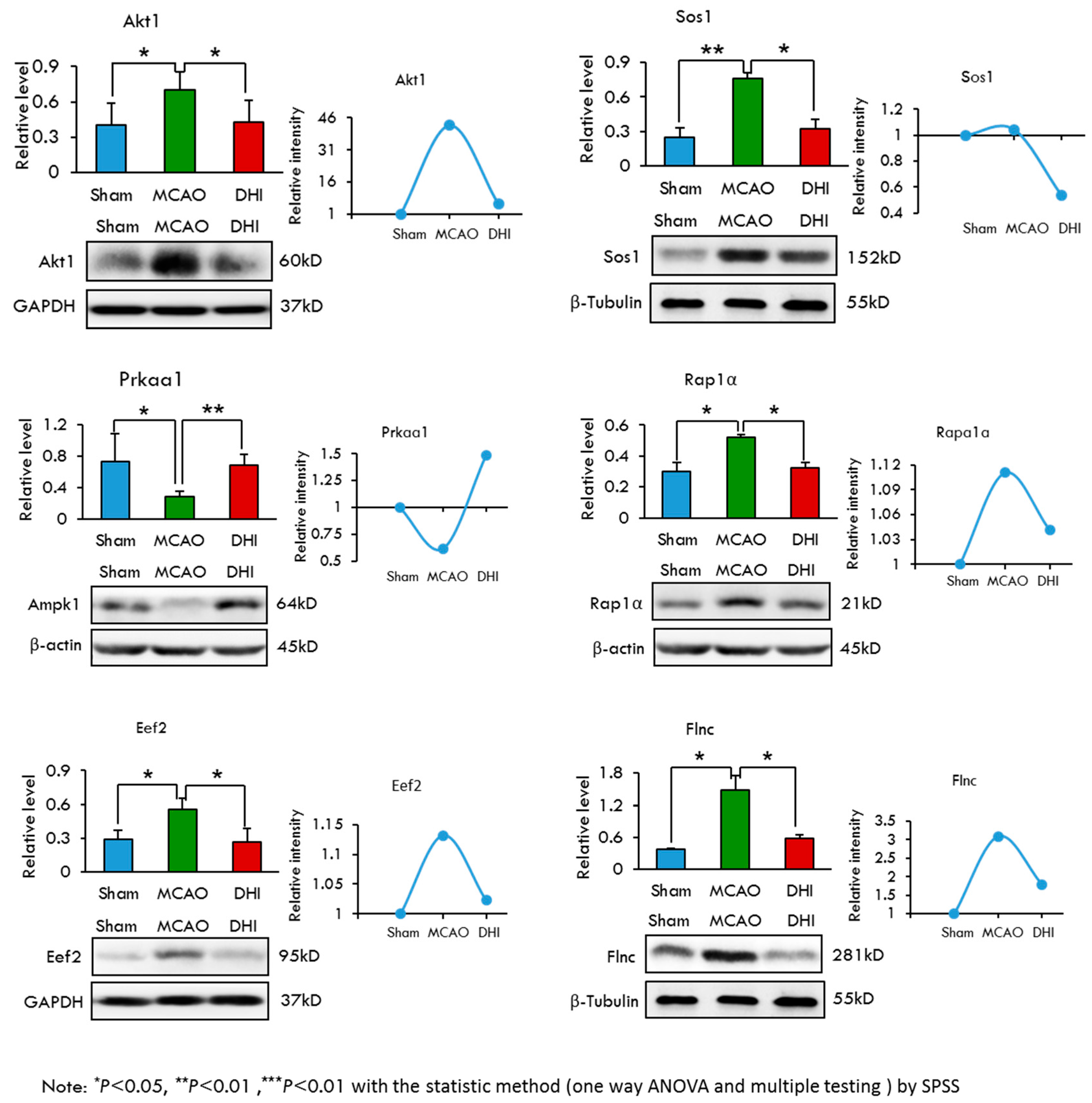
2.7. Selection and Validation of Candidate Proteins by Western Blot Analysis
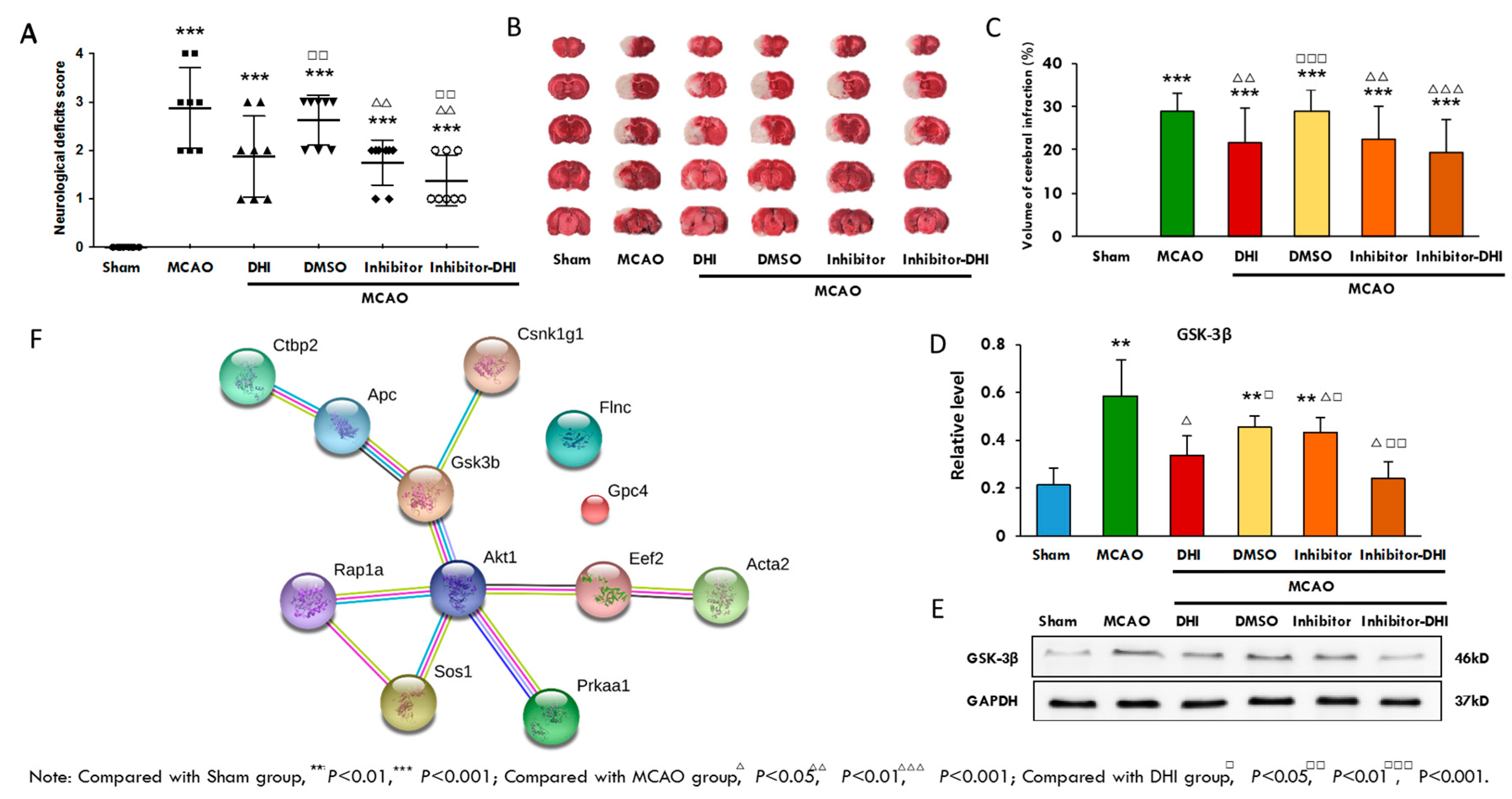
2.8. A Validation Experiment of the Protein Gsk-3β and its Inhibitor
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Treatment and Middle Cerebral Artery Occlusion
4.3. Gsk-3β Inhibition Validation Experiment
4.4. Evaluation of Neurological Deficit
4.5. Infarct Volume Measurement
4.6. Rat Hippocampus Sample Preparation
4.7. In-Solution Digestion of Hippocampal Proteins
4.8. MS Analysis for Peptide Identification
4.9. Protein Identification and Label-Free Quantitative Analysis
4.10. Bioinformatics and Statistical Analysis
4.11. Western Blotting Analysis
5. Conclusions
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Chen, X.; Du, Y.M.; Xu, F.; Liu, D.; Wang, Y.L. Propofol prevents hippocampal neuronal loss and memory impairment in cerebral ischemia injury through promoting pten degradation. J. Mol. Neurosci. 2016, 60, 63–70. [Google Scholar] [CrossRef] [PubMed]
- Kim, A.S.; Johnston, S.C. Temporal and geographic trends in the global stroke epidemic. Stroke 2013, 44, S123–S125. [Google Scholar] [CrossRef] [PubMed]
- Kuklina, E.V.; Tong, X.; George, M.G.; Bansil, P. Epidemiology and prevention of stroke: A worldwide perspective. Expert Rev. Neurother. 2012, 12, 199–208. [Google Scholar] [CrossRef] [PubMed]
- Lloyd-Jones, D.; Adams, R.; Carnethon, M.; de Simone, G.; Ferguson, T.B.; Flegal, K.; Ford, E.; Furie, K.; Go, A.; Greenlund, K.; et al. Heart disease and stroke statistics—2009 update: A report from the american heart association statistics committee and stroke statistics subcommittee. Circulation 2009, 119, 480–486. [Google Scholar] [PubMed]
- Carota, A.; Neufeld, H.; Calabrese, P. Memory profiles after unilateral paramedian thalamic stroke infarction: A comparative study. Case Rep. Med. 2015, 2015, 430869. [Google Scholar] [CrossRef] [PubMed]
- Liu, W.; Wong, A.; Au, L.; Yang, J.; Wang, Z.; Leung, E.Y.; Chen, S.; Ho, C.L.; Mok, V.C. Influence of amyloid-β on cognitive decline after stroke/transient ischemic attack: Three-year longitudinal study. Stroke 2015, 46, 3074–3080. [Google Scholar] [CrossRef] [PubMed]
- Moskowitz, M.A.; Lo, E.H.; Iadecola, C. The science of stroke: Mechanisms in search of treatments. Neuron 2010, 67, 181–198. [Google Scholar] [CrossRef] [PubMed]
- Baudry, M.; Bi, X.; Gall, C.; Lynch, G. The biochemistry of memory: The 26 year journey of a “new and specific hypothesis”. Neurobiol. Learn. Mem. 2011, 95, 125–133. [Google Scholar] [CrossRef] [PubMed]
- Kirino, T. Delayed neuronal death in the gerbil hippocampus following ischemia. Brain Res. 1982, 239, 57–69. [Google Scholar] [CrossRef]
- Petito, C.K.; Feldmann, E.; Pulsinelli, W.A.; Plum, F. Delayed hippocampal damage in humans following cardiorespiratory arrest. Neurology 1987, 37, 1281–1286. [Google Scholar] [CrossRef] [PubMed]
- Globus, M.Y.; Busto, R.; Martinez, E.; Valdes, I.; Dietrich, W.D.; Ginsberg, M.D. Comparative effect of transient global ischemia on extracellular levels of glutamate, glycine, and gamma-aminobutyric acid in vulnerable and nonvulnerable brain regions in the rat. J. Neurochem. 1991, 57, 470–478. [Google Scholar] [CrossRef] [PubMed]
- Kirino, T.; Sano, K. Selective vulnerability in the gerbil hippocampus following transient ischemia. Acta Neuropathol. 1984, 62, 201–208. [Google Scholar] [CrossRef] [PubMed]
- Stoll, G.; Jander, S.; Schroeter, M. Inflammation and glial responses in ischemic brain lesions. Prog. Neurobiol. 1998, 56, 149–171. [Google Scholar] [CrossRef]
- White, B.C.; Sullivan, J.M.; DeGracia, D.J.; O’Neil, B.J.; Neumar, R.W.; Grossman, L.I.; Rafols, J.A.; Krause, G.S. Brain ischemia and reperfusion: Molecular mechanisms of neuronal injury. J. Neurol. Sci. 2000, 179, 1–33. [Google Scholar] [CrossRef]
- An, S.J.; Kang, T.C.; Park, S.K.; Hwang, I.K.; Cho, S.S.; Chung, M.H.; Won, M.H. Oxidative DNA damage and alteration of glutamate transporter expressions in the hippocampal ca1 area immediately after ischemic insult. Mol. Cells 2002, 13, 476–480. [Google Scholar] [PubMed]
- Sun, M.; Zhang, J.J.; Shan, J.Z.; Zhang, H.; Jin, C.Y.; Xu, S.; Wang, Y.L. Clinical observation of Danhong injection (herbal TCM product from Radix Salviae miltiorrhizae and Flos Carthami tinctorii) in the treatment of traumatic intracranial hematoma. Phytomedicine 2009, 16, 683–689. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Wang, S.; Sun, A.; Huang, D.; Wang, W.; Zhang, C.; Shi, D.; Chen, K.; Zou, Y.; Ge, J. Danhong inhibits oxidized low-density lipoprotein-induced immune maturation of dentritic cells via a peroxisome proliferator activated receptor gamma-mediated pathway. J. Pharmacol. Sci. 2012, 119, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Guo, H.; Li, M.J.; Liu, Q.Q.; Guo, L.L.; Ma, M.M.; Wang, S.X.; Yu, B.; Hu, L.M. Danhong injection attenuates ischemia/reperfusion-induced brain damage which is associating with Nrf2 levels in vivo and in vitro. Neurochem. Res. 2014, 39, 1817–1824. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Yi, D.; Xie, Y.; Yang, W.; Yang, W.; Zhuang, Y.; Du, J. Analysis of clinical use of Danhong injection based on hospital information system. Zhongguo Zhong Yao Za Zhi 2011, 36, 2817–2820. [Google Scholar] [PubMed]
- Wei, J.; Zhang, Y.; Jia, Q.; Liu, M.; Li, D.; Zhang, Y.; Song, L.; Hu, Y.; Xian, M.; Yang, H.; et al. Systematic investigation of transcription factors critical in the protection against cerebral ischemia by Danhong injection. Sci. Rep. 2016, 6, 29823. [Google Scholar] [CrossRef] [PubMed]
- Aebersold, R.; Mann, M. Mass spectrometry-based proteomics. Nature 2003, 422, 198–207. [Google Scholar] [CrossRef] [PubMed]
- Bantscheff, M.; Schirle, M.; Sweetman, G.; Rick, J.; Kuster, B. Quantitative mass spectrometry in proteomics: A critical review. Anal. Bioanal. Chem. 2007, 389, 1017–1031. [Google Scholar] [CrossRef] [PubMed]
- Schulze, W.X.; Usadel, B. Quantitation in mass-spectrometry-based proteomics. Annu. Rev. Plant Biol. 2010, 61, 491–516. [Google Scholar] [CrossRef] [PubMed]
- Wasinger, V.C.; Zeng, M.; Yau, Y. Current status and advances in quantitative proteomic mass spectrometry. Int. J. Proteomics 2013, 2013, 180605. [Google Scholar] [CrossRef] [PubMed]
- Longa, E.Z.; Weinstein, P.R.; Carlson, S.; Cummins, R. Reversible middle cerebral artery occlusion without craniectomy in rats. Stroke 1989, 20, 84–91. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.; Chen, C.; Chen, J.X.; Wen, L.M.; Yang, G.L.; Duan, F.P.; Huang, Z.Y.; Li, D.F.; Yu, D.R.; Yang, H.J.; et al. Synergism and rules of the new combination drug Yiqijiedu formulae (YQJD) on ischemic stroke based on amino acids (AAs) metabolism. Sci. Rep. 2014, 4, 5149. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.L.; Zhang, S.P.; Zhu, H.B.; Jian, H.; Tian, J.W. Cornin ameliorates cerebral infarction in rats by antioxidant action and stabilization of mitochondrial function. Phytother. Res. 2010, 24, 547–552. [Google Scholar] [CrossRef] [PubMed]
- Astrup, J.; Siesjo, B.K.; Symon, L. Thresholds in cerebral ischemia—The ischemic penumbra. Stroke 1981, 12, 723–725. [Google Scholar] [CrossRef] [PubMed]
- Memezawa, H.; Smith, M.L.; Siesjo, B.K. Penumbral tissues salvaged by reperfusion following middle cerebral artery occlusion in rats. Stroke 1992, 23, 552–559. [Google Scholar] [CrossRef] [PubMed]
- Paciaroni, M.; Caso, V.; Agnelli, G. The concept of ischemic penumbra in acute stroke and therapeutic opportunities. Eur. Neurol. 2009, 61, 321–330. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wan, H.; Du, Y.; Bie, X.; Zhao, T.; Fu, W.; Xing, P. Protective effect of Danhong injection on cerebral ischemia-reperfusion injury in rats. J. Ethnopharmacol. 2012, 144, 387–394. [Google Scholar] [CrossRef] [PubMed]
- Loh, K.P.; Huang, S.H.; Tan, B.K.; Zhu, Y.Z. Cerebral protection of purified herba leonuri extract on middle cerebral artery occluded rats. J. Ethnopharmacol. 2009, 125, 337–343. [Google Scholar] [CrossRef] [PubMed]
- Mi, H.; Muruganujan, A.; Casagrande, J.T.; Thomas, P.D. Large-scale gene function analysis with the panther classification system. Nat. Protoc. 2013, 8, 1551–1566. [Google Scholar] [CrossRef] [PubMed]
- Eichelbaum, K.; Winter, M.; Diaz, M.B.; Herzig, S.; Krijgsveld, J. Selective enrichment of newly synthesized proteins for quantitative secretome analysis. Nat. Biotechnol. 2012, 30, 984–990. [Google Scholar] [CrossRef] [PubMed]
- Dale, T.C. Signal transduction by the Wnt family of ligands. Biochem. J. 1998, 329 Pt 2, 209–223. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, S.; Chuang, D.M. Neuroprotective effects of chronic lithium on focal cerebral ischemia in rats. Neuroreport 1998, 9, 2081–2084. [Google Scholar] [CrossRef] [PubMed]
- Hashimoto, M.; Sagara, Y.; Langford, D.; Everall, I.P.; Mallory, M.; Everson, A.; Digicaylioglu, M.; Masliah, E. Fibroblast growth factor 1 regulates signaling via the glycogen synthase kinase-3β pathway. Implications for neuroprotection. J. Biol. Chem. 2002, 277, 32985–32991. [Google Scholar] [CrossRef] [PubMed]
- Chen, G.; Bower, K.A.; Ma, C.; Fang, S.; Thiele, C.J.; Luo, J. Glycogen synthase kinase 3β (GSK3β) mediates 6-hydroxydopamine-induced neuronal death. FASEB J. 2004, 18, 1162–1164. [Google Scholar] [PubMed]
- Nusse, R. Wnt signaling in disease and in development. Cell Res. 2005, 15, 28–32. [Google Scholar] [CrossRef] [PubMed]
- Van Amerongen, R.; Nusse, R. Towards an integrated view of Wnt signaling in development. Development 2009, 136, 3205–3214. [Google Scholar] [CrossRef] [PubMed]
- Angers, S.; Moon, R.T. Proximal events in Wnt signal transduction. Nat. Rev. Mol. Cell Biol. 2009, 10, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Goessling, W.; North, T.E.; Loewer, S.; Lord, A.M.; Lee, S.; Stoick-Cooper, C.L.; Weidinger, G.; Puder, M.; Daley, G.Q.; Moon, R.T.; et al. Genetic interaction of PGE2 and Wnt signaling regulates developmental specification of stem cells and regeneration. Cell 2009, 136, 1136–1147. [Google Scholar] [CrossRef] [PubMed]
- Wassink, T.H.; Piven, J.; Vieland, V.J.; Huang, J.; Swiderski, R.E.; Pietila, J.; Braun, T.; Beck, G.; Folstein, S.E.; Haines, J.L.; et al. Evidence supporting WNT2 as an autism susceptibility gene. Am. J. Med. Gene. 2001, 105, 406–413. [Google Scholar] [CrossRef] [PubMed]
- Cotter, D.; Kerwin, R.; al-Sarraji, S.; Brion, J.P.; Chadwich, A.; Lovestone, S.; Anderton, B.; Everall, I. Abnormalities of Wnt signalling in schizophrenia—Evidence for neurodevelopmental abnormality. Neuroreport 1998, 9, 1379–1383. [Google Scholar] [CrossRef] [PubMed]
- Miyaoka, T.; Seno, H.; Ishino, H. Increased expression of Wnt-1 in schizophrenic brains. Schizophr. Res. 1999, 38, 1–6. [Google Scholar] [CrossRef]
- Dean, B. Understanding the pathology of schizophrenia: Recent advances from the study of the molecular architecture of postmortem cns tissue. Postgrad. Med. J. 2002, 78, 142–148. [Google Scholar] [CrossRef] [PubMed]
- Howng, S.L.; Wu, C.H.; Cheng, T.S.; Sy, W.D.; Lin, P.C.; Wang, C.; Hong, Y.R. Differential expression of Wnt genes, β-catenin and E-cadherin in human brain tumors. Cancer Lett. 2002, 183, 95–101. [Google Scholar] [CrossRef]
- Willert, K.; Nusse, R. α-catenin: A key mediator of Wnt signaling. Curr. Opin. Genet. Dev. 1998, 8, 95–102. [Google Scholar] [CrossRef]
- Chen, R.H.; Ding, W.V.; McCormick, F. Wnt signaling to β-catenin involves two interactive components. Glycogen synthase kinase-3β inhibition and activation of protein kinase C. J. Biol. Chem. 2000, 275, 17894–17899. [Google Scholar] [CrossRef] [PubMed]
- Fukumoto, S.; Hsieh, C.M.; Maemura, K.; Layne, M.D.; Yet, S.F.; Lee, K.H.; Matsui, T.; Rosenzweig, A.; Taylor, W.G.; Rubin, J.S.; et al. Akt participation in the Wnt signaling pathway through dishevelled. J. Biol. Chem. 2001, 276, 17479–17483. [Google Scholar] [CrossRef] [PubMed]
- Grimes, C.A.; Jope, R.S. Creb DNA binding activity is inhibited by glycogen synthase kinase-3β and facilitated by lithium. J. Neurochem. 2001, 78, 1219–1232. [Google Scholar] [CrossRef] [PubMed]
- Veugelers, M.; Vermeesch, J.; Watanabe, K.; Yamaguchi, Y.; Marynen, P.; David, G. GPC4, the gene for human K-glypican, flanks GPC3 on Xq26: Deletion of the GPC3–GPC4 gene cluster in one family with simpson-golabi-behmel syndrome. Genomics 1998, 53, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Jack, B.H.; Pearson, R.C.; Crossley, M. C-terminal binding protein: A metabolic sensor implicated in regulating adipogenesis. Int. J. Biochem. Cell Biol. 2011, 43, 693–696. [Google Scholar] [CrossRef] [PubMed]
- Ueyama, H.; Bruns, G.; Kanda, N. Assignment of the vascular smooth muscle actin gene ACTSA to human chromosome 10. Jpn. J. Hum. Genet. 1990, 35, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Wan, D.; Zhou, Y.; Wang, K.; Hou, Y.; Hou, R.; Ye, X. Resveratrol provides neuroprotection by inhibiting phosphodiesterases and regulating the cAMP/AMPK/SIRT1 pathway after stroke in rats. Brain Res. Bull. 2016, 121, 255–262. [Google Scholar] [CrossRef] [PubMed]
- An, J.Y.; Zhou, L.L.; Sun, P.; Pang, H.G.; Li, D.D.; Li, Y.; Zhang, M.; Song, J.N. Role of the AMPK signaling pathway in early brain injury after subarachnoid hemorrhage in rats. Acta Neurochir. 2015, 157, 781–792. [Google Scholar] [CrossRef] [PubMed]
- Sun, J.; Nan, G. The mitogen-activated protein kinase (MAPK) signaling pathway as a discovery target in stroke. J. Mol. Neurosci. 2016, 59, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Ansar, S.; Eftekhari, S.; Waldsee, R.; Nilsson, E.; Nilsson, O.; Saveland, H.; Edvinsson, L. MAPK signaling pathway regulates cerebrovascular receptor expression in human cerebral arteries. BMC Neurosci. 2013, 14, 12. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Watson, C.R.; Emson, P.C. AChE-stained horizontal sections of the rat brain in stereotaxic coordinates. J. Neurosci. Methods 1980, 3, 129–149. [Google Scholar] [CrossRef]
- Saraf, M.K.; Prabhakar, S.; Anand, A. Neuroprotective effect of bacopa monniera on ischemia induced brain injury. Pharmacol. Biochem. Behav. 2010, 97, 192–197. [Google Scholar] [CrossRef] [PubMed]
- Alessandrini, A.; Namura, S.; Moskowitz, M.A.; Bonventre, J.V. MEK1 protein kinase inhibition protects against damage resulting from focal cerebral ischemia. Proc. Natl. Acad. Sci. USA 1999, 96, 12866–12869. [Google Scholar] [CrossRef] [PubMed]
- Wisniewski, J.R.; Zougman, A.; Nagaraj, N.; Mann, M. Universal sample preparation method for proteome analysis. Nat. Methods 2009, 6, 359–362. [Google Scholar] [CrossRef] [PubMed]
- Boersema, P.J.; Geiger, T.; Wisniewski, J.R.; Mann, M. Quantification of the N-glycosylated secretome by super-SILAC during breast cancer progression and in human blood samples. Mol. Cell Proteomics 2013, 12, 158–171. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Jiang, J.; Zhao, X.; Zhao, Y.; Cao, Q.; Zhao, Q.; Han, H.; Wang, J.; Yu, Z.; Peng, B.; et al. In-depth analysis of secretome and N-glycosecretome of human hepatocellular carcinoma metastatic cell lines shed light on metastasis correlated proteins. Oncotarget 2016, 7, 22031. [Google Scholar] [PubMed]
- Tyanova, S.; Temu, T.; Sinitcyn, P.; Carlson, A.; Hein, M.Y.; Geiger, T.; Mann, M.; Cox, J. The perseus computational platform for comprehensive analysis of (prote) omics data. Nat. Methods 2016, 13, 731–740. [Google Scholar] [CrossRef] [PubMed]
- Karagiannis, G.S.; Pavlou, M.P.; Saraon, P.; Musrap, N.; Xie, A.; Batruch, I.; Prassas, I.; Dimitromanolakis, A.; Petraki, C.; Diamandis, E.P. In-depth proteomic delineation of the colorectal cancer exoproteome: Mechanistic insight and identification of potential biomarkers. J. Proteomics 2014, 103, 121–136. [Google Scholar] [CrossRef] [PubMed]
- Saeed, A.I.; Bhagabati, N.K.; Braisted, J.C.; Liang, W.; Sharov, V.; Howe, E.A.; Li, J.; Thiagarajan, M.; White, J.A.; Quackenbush, J. TM4 microarray software suite. Methods Enzymol. 2006, 411, 134–193. [Google Scholar] [PubMed]
- Ashburner, M.; Ball, C.A.; Blake, J.A.; Botstein, D.; Butler, H.; Cherry, J.M.; Davis, A.P.; Dolinski, K.; Dwight, S.S.; Eppig, J.T.; et al. Gene ontology: Tool for the unification of biology. The gene ontology consortium. Nat. Genet. 2000, 25, 25–29. [Google Scholar] [CrossRef] [PubMed]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using david bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef] [PubMed]
- Sun, N.; Sun, W.; Li, S.; Yang, J.; Yang, L.; Quan, G.; Gao, X.; Wang, Z.; Cheng, X.; Li, Z.; et al. Proteomics analysis of cellular proteins co-immunoprecipitated with nucleoprotein of influenza a virus (H7N9). Int. J. Mol. Sci. 2015, 16, 25982–25998. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Xu, X.; Zhou, B.; He, Z.; Zhai, Q. Gene expression profile change and associated physiological and pathological effects in mouse liver induced by fasting and refeeding. PloS ONE 2011, 6, e27553. [Google Scholar]
- Zhu, Y.M.; Wang, C.C.; Chen, L.; Qian, L.B.; Ma, L.L.; Yu, J.; Zhu, M.H.; Wen, C.Y.; Yu, L.N.; Yan, M. Both PI3K/Akt and ERK1/2 pathways participate in the protection by dexmedetomidine against transient focal cerebral ischemia/reperfusion injury in rats. Brain Res. 2013, 1494, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Shen, C.; Yu, Y.; Li, H.; Yan, G.; Liu, M.; Shen, H.; Yang, P. Global profiling of proteolytically modified proteins in human metastatic hepatocellular carcinoma cell lines reveals CAPN2 centered network. Proteomics 2012, 12, 1917–1927. [Google Scholar] [CrossRef] [PubMed]
- Mann, M.; Kulak, N.A.; Nagaraj, N.; Cox, J. The coming age of complete, accurate, and ubiquitous proteomes. Mol. Cell 2013, 49, 583–590. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Groups | Mean Score |
|---|---|
| Naive | 0.00 ± 0.00 |
| Sham | 0.00 ± 0.00 |
| MCAO | 2.75 ± 0.43 *** |
| DHI | 1.50 ± 0.50 ***,∆∆ |
| Ginaton | 1.62 ± 0.48 ***,∆∆ |
| Groups | Cerebral Infarct Volume (%) |
|---|---|
| Sham | 0.00 ± 0.00 |
| MCAO | 29.71 ± 3.47 *** |
| Ginaton-treated | 21.77 ± 7.71 ***,∆ |
| Danhong-treated | 22.25 ± 7.94 ***,∆ |
| Name | Groups | Protein Relative Level(Mean ± SD) |
|---|---|---|
| Gsk-3β | Sham | 0.352 ± 0.135 |
| MCAO | 0.421 ± 0.163 * | |
| DHI | 0.359 ± 0.169 * | |
| Acta2 | Sham | 1.693 ± 0.670 |
| MCAO | 0.925 ± 0.527 * | |
| DHI | 1.338 ± 0.762 * | |
| Csk1g1 | Sham | 0.350 ± 0.048 |
| MCAO | 0.575 ± 0.048 ** | |
| DHI | 0.344 ± 0.077 ** | |
| Apc | Sham | 0.804 ± 0.250 |
| MCAO | 0.371 ± 0.051 * | |
| DHI | 0.743 ± 0.163 * | |
| Gpc4 | Sham | 0.330 ± 0.188 |
| MCAO | 1.073 ± 0.257 * | |
| DHI | 0.379 ± 0.023 * | |
| Ctbp2 | Sham | 0.749 ± 0.480 |
| MCAO | 1.103 ± 0.463 ** | |
| DHI | 1.018 ± 0.460 | |
| Akt1 | Sham | 0.404 ± 0.187 |
| MCAO | 0.700 ± 0.155 * | |
| DHI | 0.430 ± 0.182 * | |
| Sos1 | Sham | 0.247 ± 0.083 |
| MCAO | 0.765 ± 0.050 ** | |
| DHI | 0.322 ± 0.085 * | |
| Prkaa1 | Sham | 0.733 ± 0.353 |
| MCAO | 0.282 ± 0.075 * | |
| DHI | 0.684 ± 0.134 ** | |
| Eef2 | Sham | 0.288 ± 0.084 |
| MCAO | 0.558 ± 0.091 * | |
| DHI | 0.265 ± 0.120 * | |
| Flnc | Sham | 0.370 ± 0.021 |
| MCAO | 1.477 ± 0.276 * | |
| DHI | 0.587 ± 0.069 * | |
| Rap1a | Sham | 0.302 ± 0.058 |
| MCAO | 0.521 ± 0.020 * | |
| DHI | 0.322 ± 0.038 * |
| Groups | Mean Score |
|---|---|
| Sham | 0.00 ± 0.00 |
| MCAO | 2.88 ± 0.83 *** |
| DHI | 1.88 ± 0.78 *** |
| MCAO + DMSO | 2.62 ± 0.48 ***,□□ |
| MCAO + Inhibitor | 1.75 ± 0.43 ***,∆∆ |
| MCAO + Inhibitor + DHI | 1.38 ± 0.48 ***,∆∆□□ |
| Groups | Cerebral Infarct Volume (%) |
|---|---|
| Sham | 0.00 ± 0.00 |
| MCAO | 28.78 ± 3.92 *** |
| DHI | 21.79 ± 4.54 ***,∆∆ |
| MCAO + DMSO | 28.75 ± 3.28 ***,□□□ |
| MCAO + inhibitor | 22.62 ± 4.26 ***,∆∆ |
| MCAO + inhibitor + DHI | 19.42 ± 3.48 ***,△△△ |
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cui, Y.; Liu, X.; Li, X.; Yang, H. In-Depth Proteomic Analysis of the Hippocampus in a Rat Model after Cerebral Ischaemic Injury and Repair by Danhong Injection (DHI). Int. J. Mol. Sci. 2017, 18, 1355. https://doi.org/10.3390/ijms18071355
Cui Y, Liu X, Li X, Yang H. In-Depth Proteomic Analysis of the Hippocampus in a Rat Model after Cerebral Ischaemic Injury and Repair by Danhong Injection (DHI). International Journal of Molecular Sciences. 2017; 18(7):1355. https://doi.org/10.3390/ijms18071355
Chicago/Turabian StyleCui, Yiran, Xin Liu, Xianyu Li, and Hongjun Yang. 2017. "In-Depth Proteomic Analysis of the Hippocampus in a Rat Model after Cerebral Ischaemic Injury and Repair by Danhong Injection (DHI)" International Journal of Molecular Sciences 18, no. 7: 1355. https://doi.org/10.3390/ijms18071355