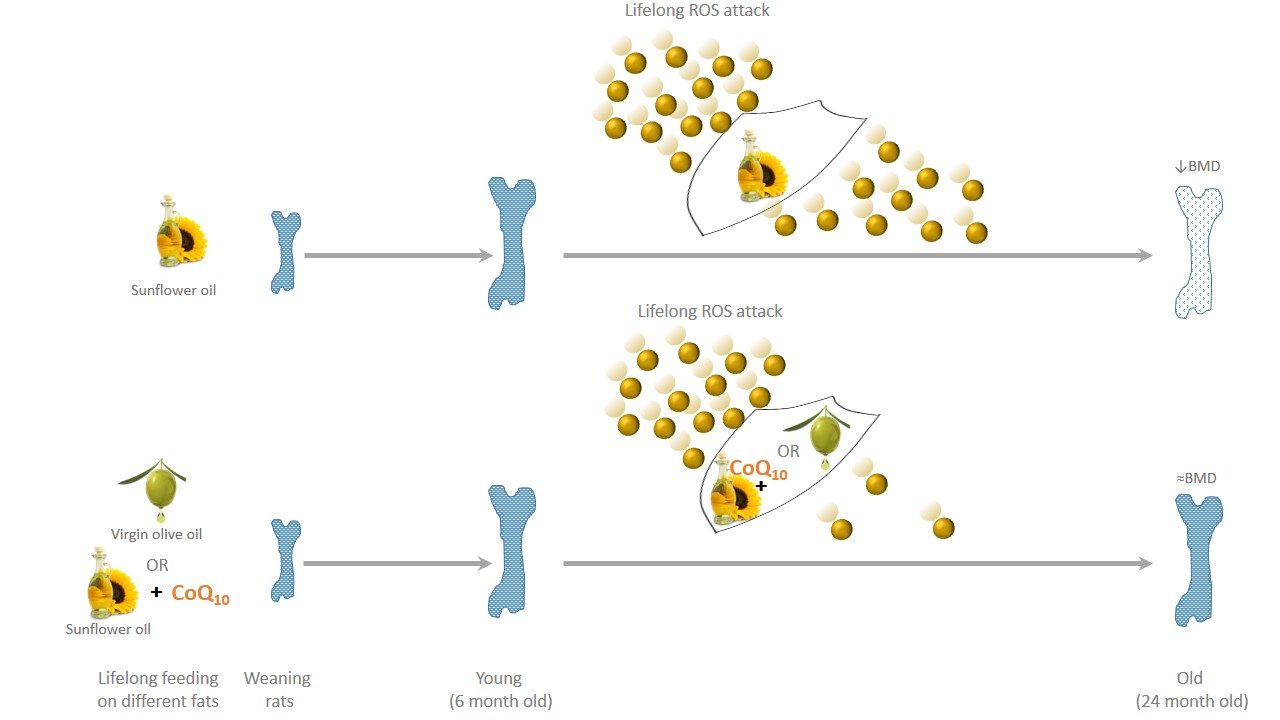
Loss of Bone Mineral Density Associated with Age in Male Rats Fed on Sunflower Oil Is Avoided by Virgin Olive Oil Intake or Coenzyme Q Supplementation

 , , , and
, , , and
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
2.1. Animal Weight
2.2. Plasma Fatty Acid Profile and Total Coenzyme Q Content
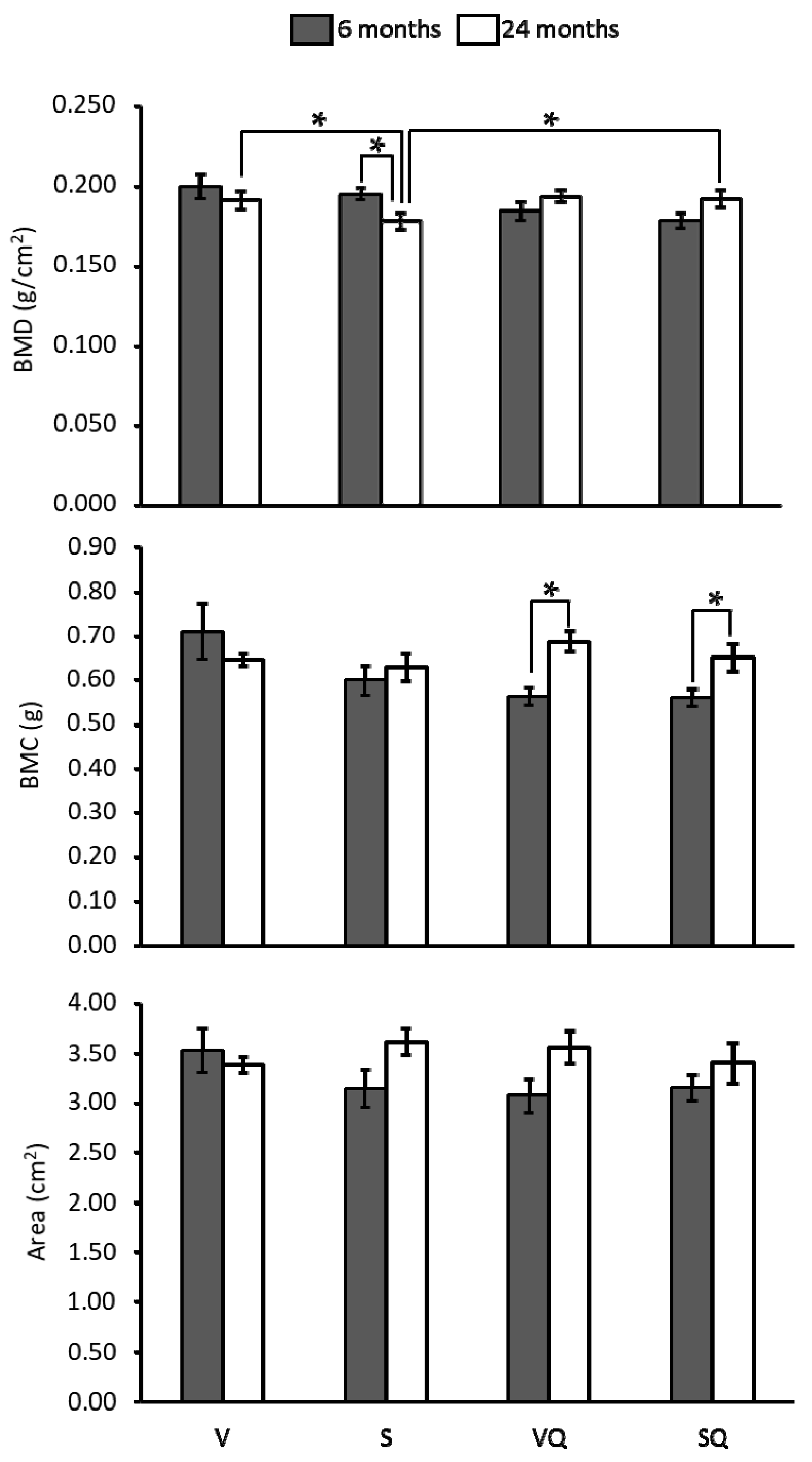
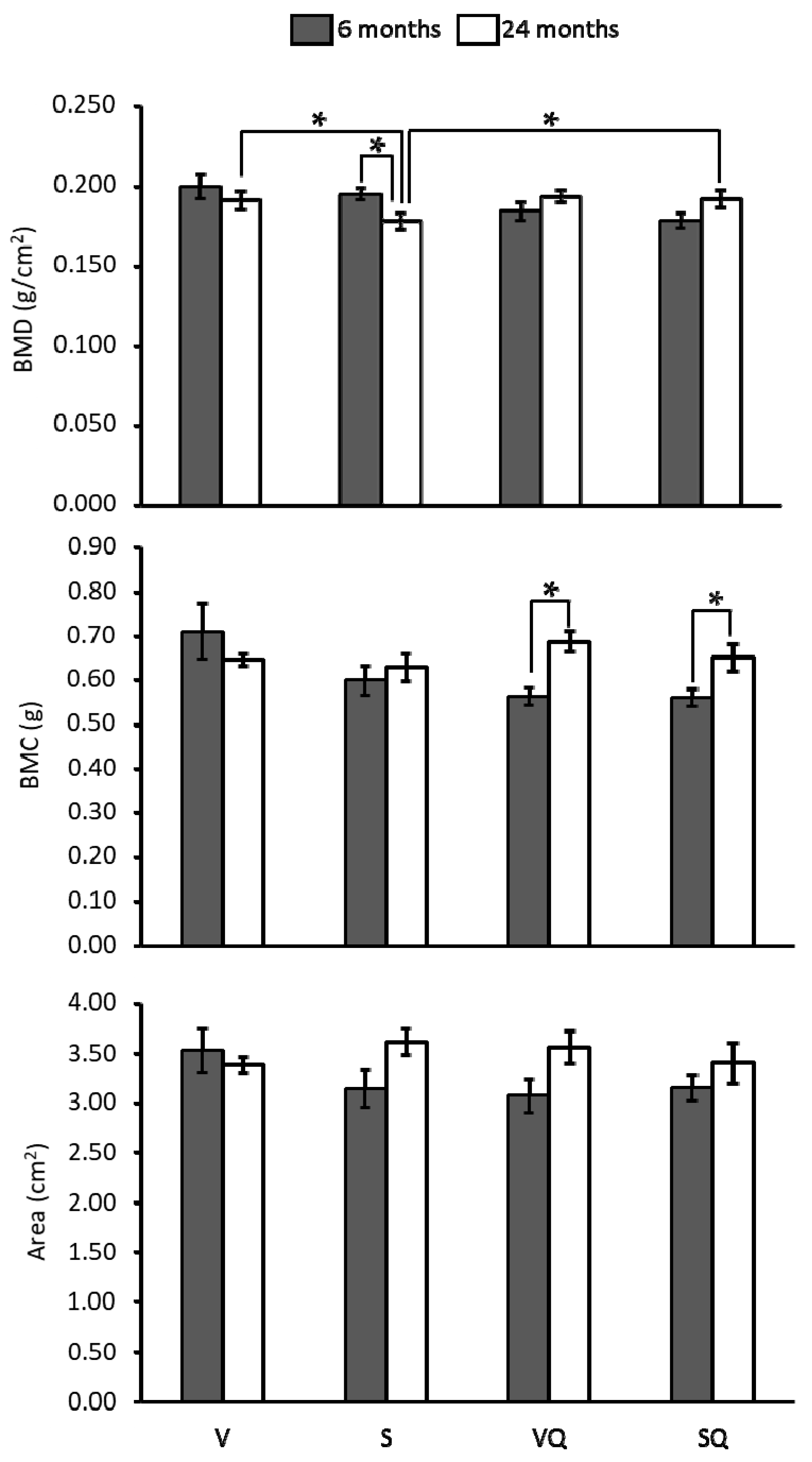
2.3. Bone Mineral Density, Bone Mineral Content (BMC) and Bone Areal Size
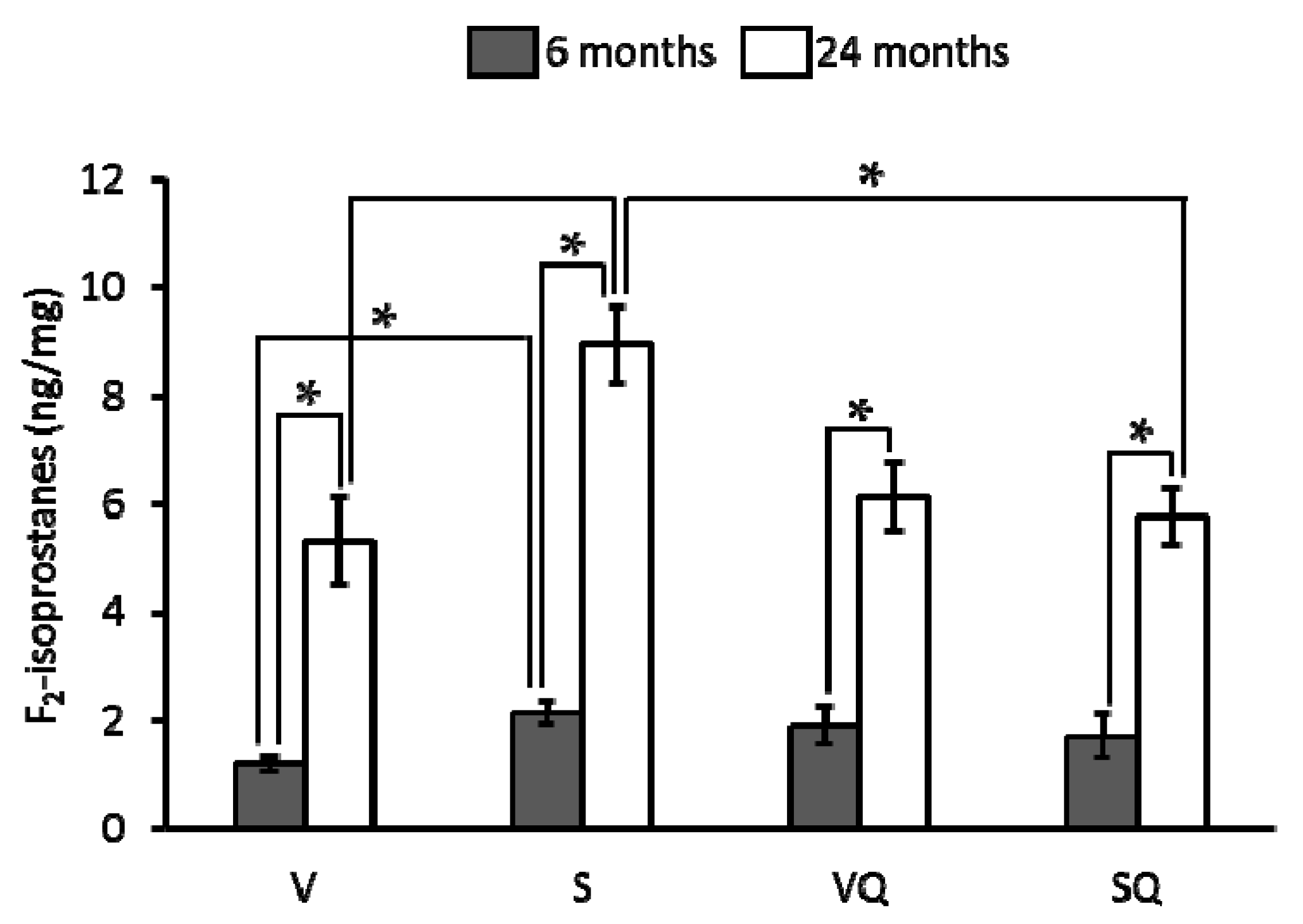
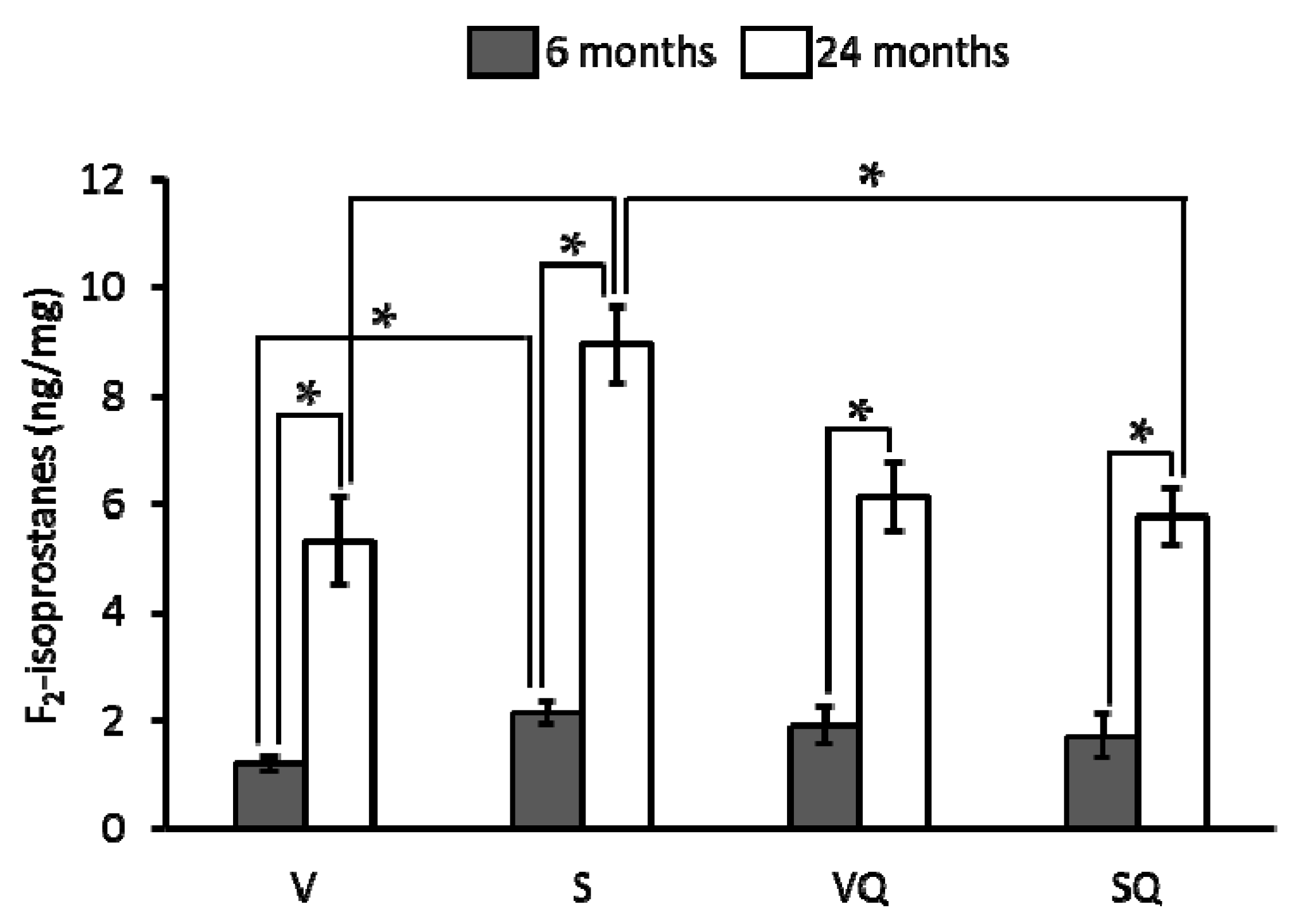
2.4. Urinary F2-Isoprostanes
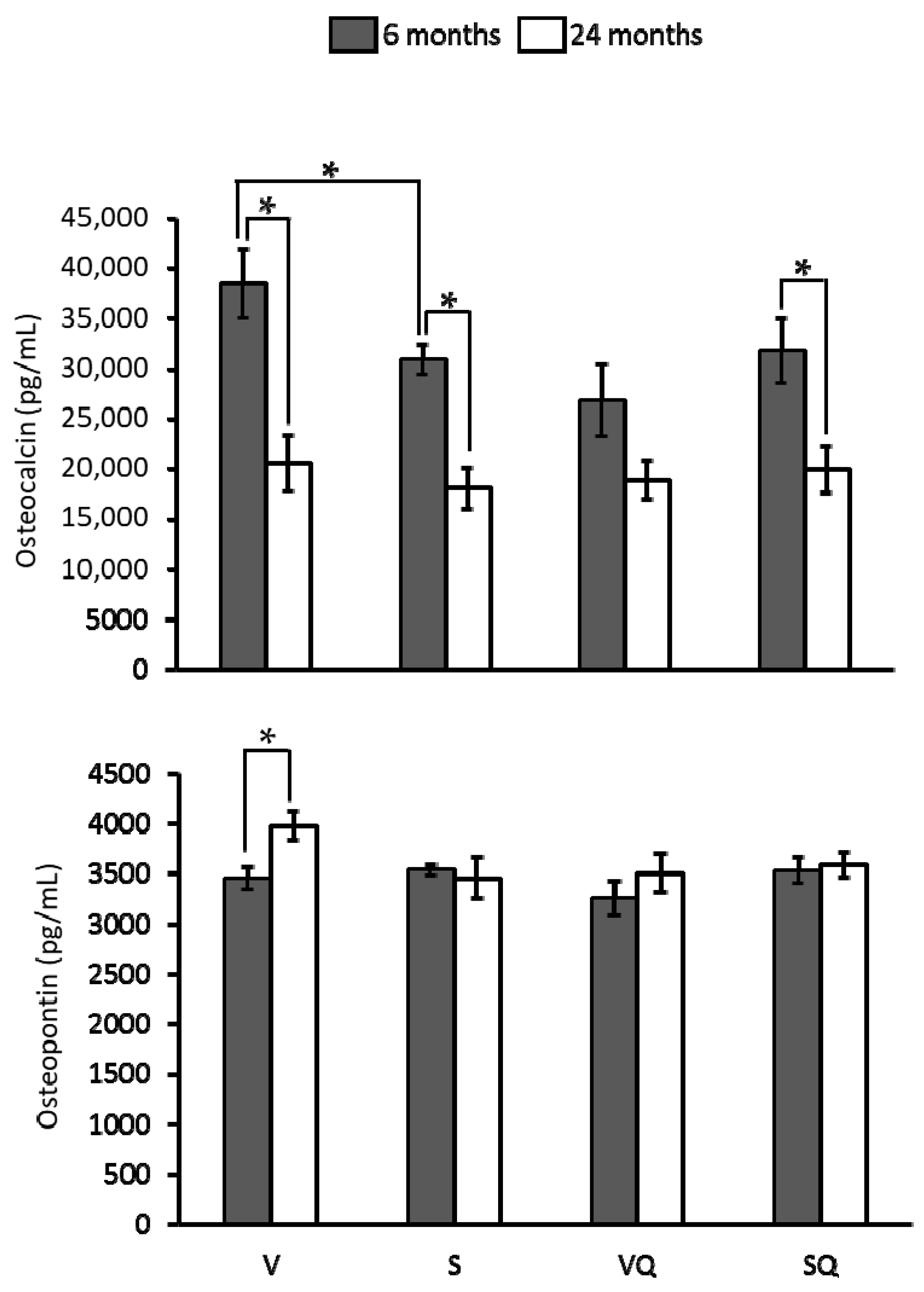
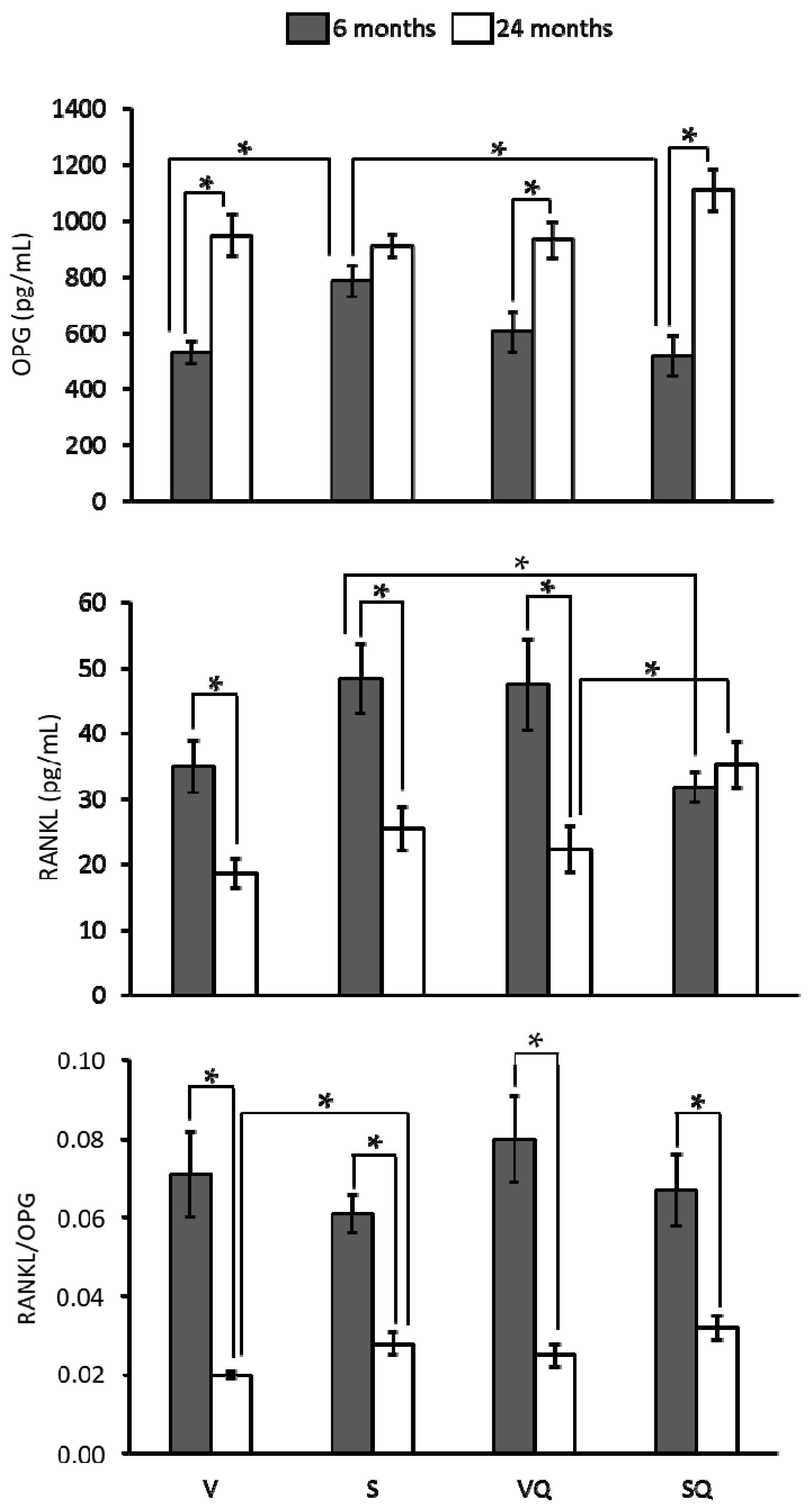
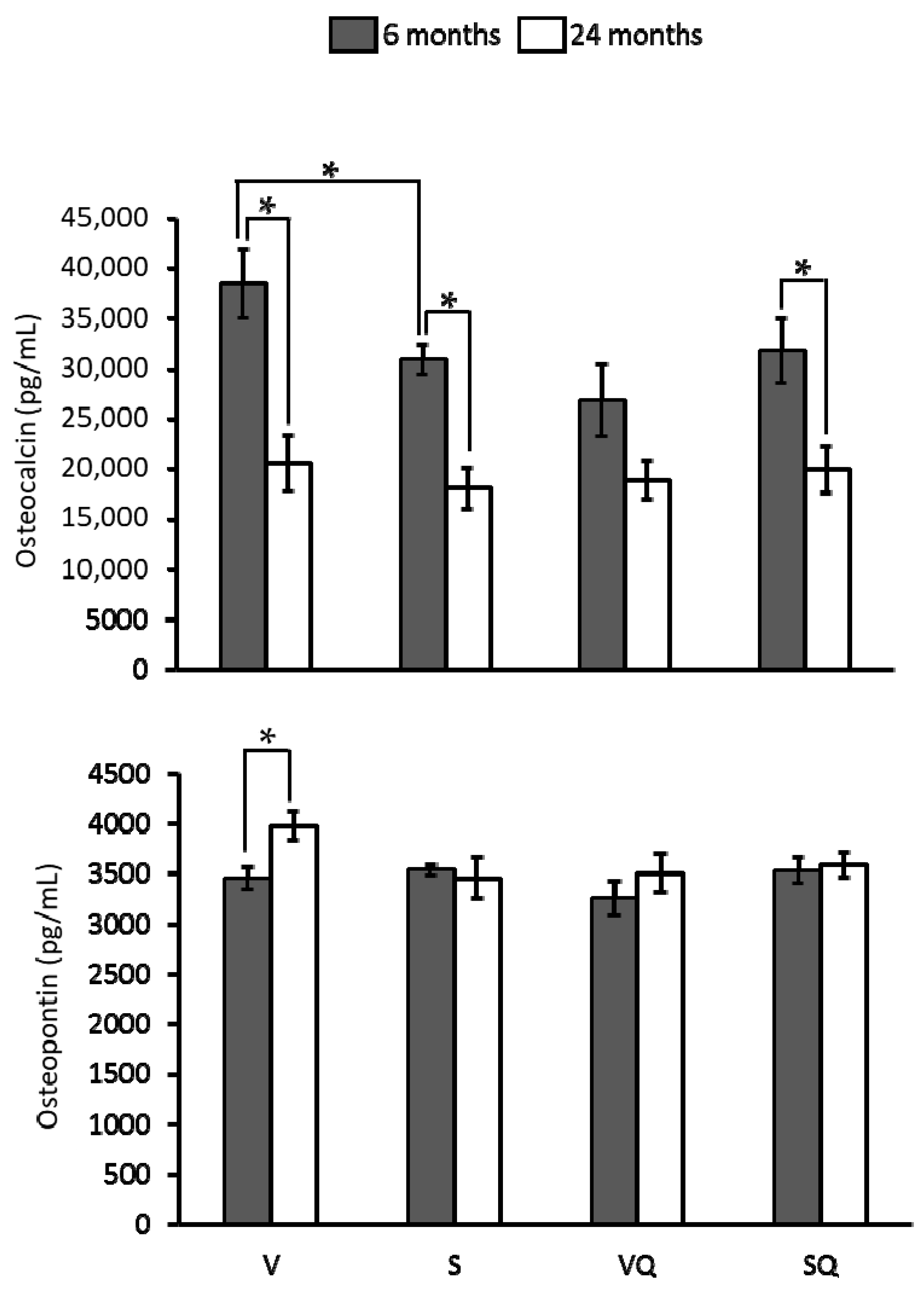
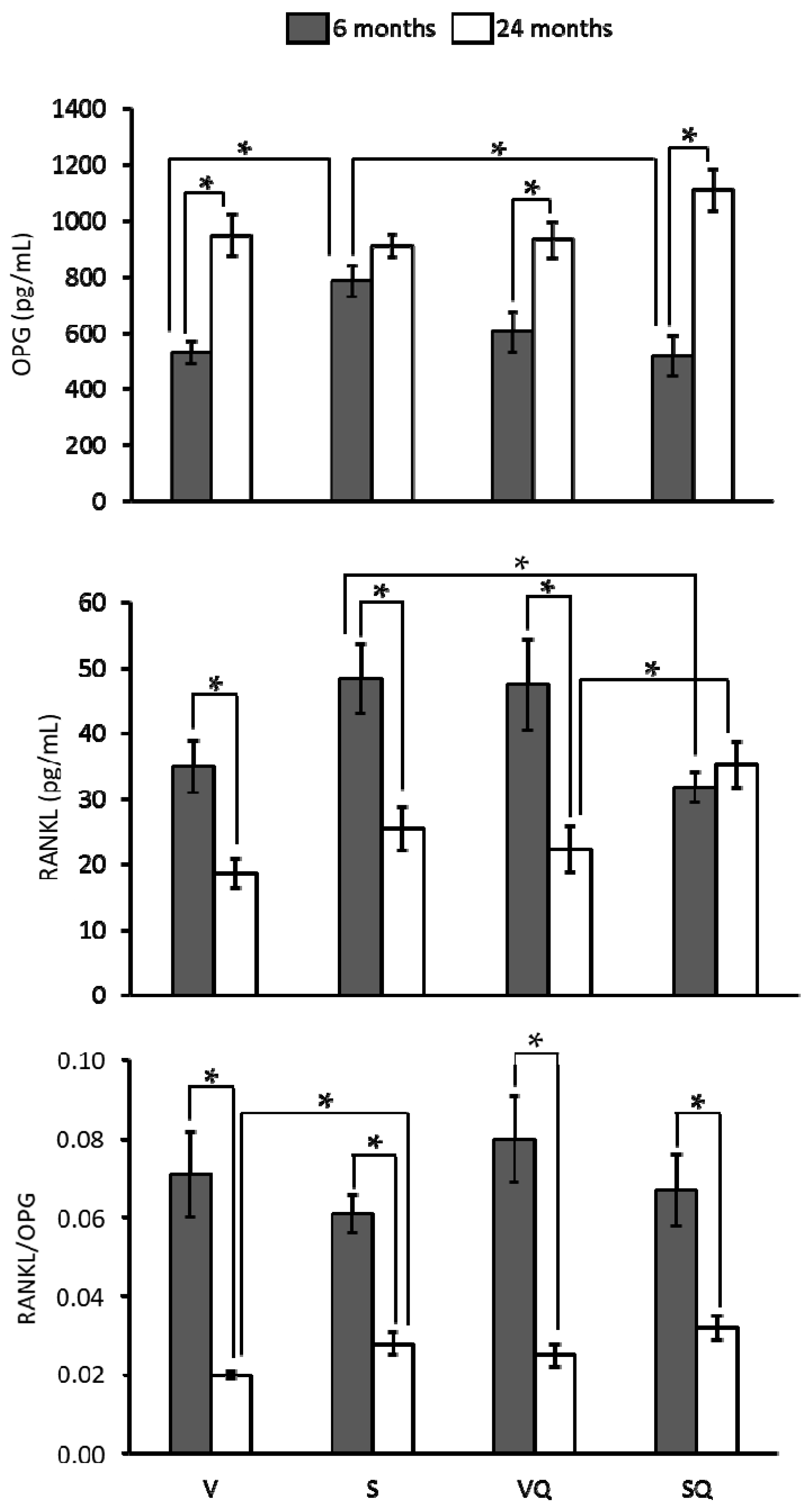
2.5. Circulating Levels of Ostecalcin and Osteopontin
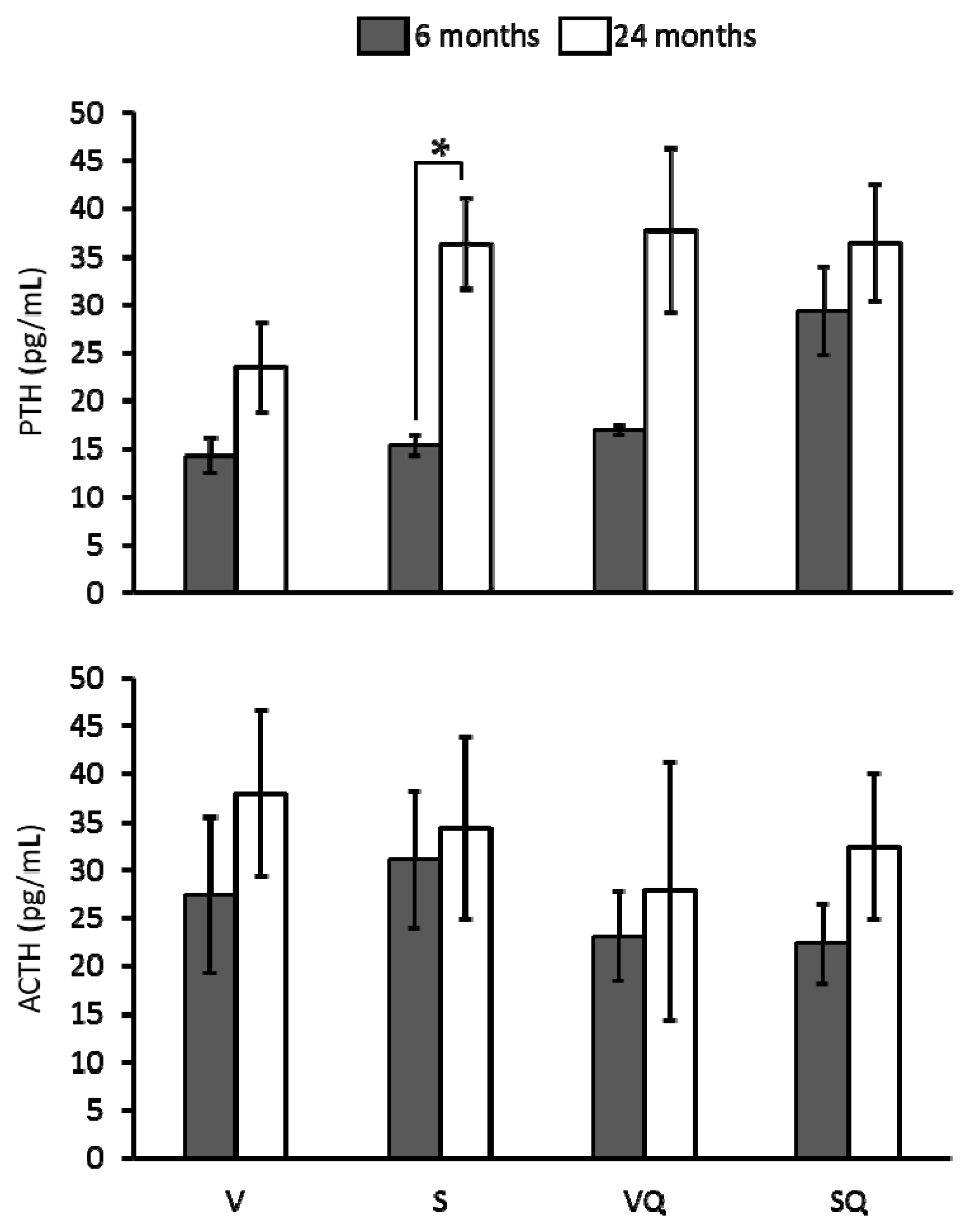
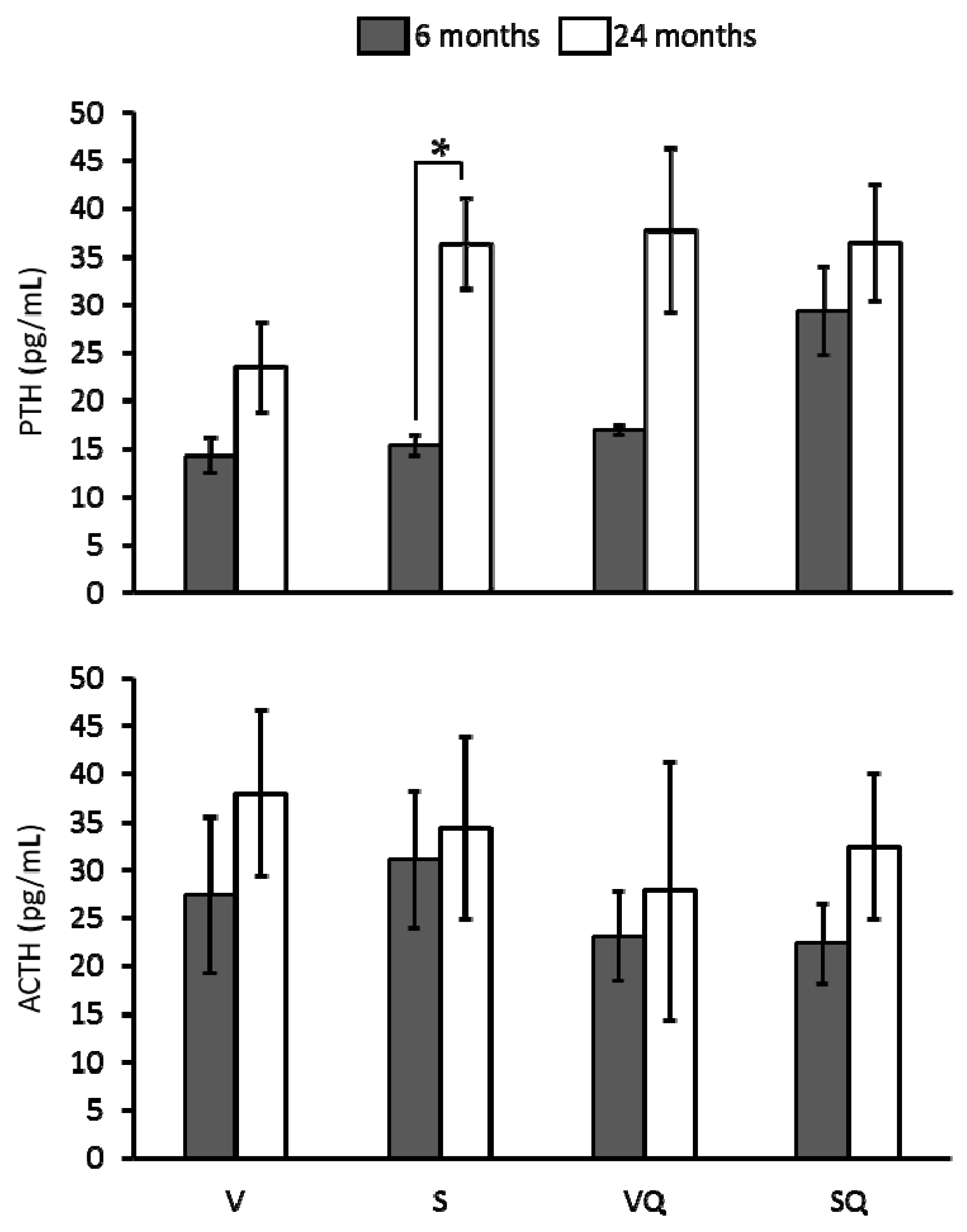
2.6. Circulating Levels of Parathyroid Hormone (PTH) and Adrenocorticotropin (ACTH)
3. Discussion
4. Materials and Methods
4.1. Experimental Design
4.2. Determination of Bone Mineral Density
4.3. Urinary F2-Isoprostanes Determination
4.4. Determination of Bone Metabolism Markers, RANKL, OPG, and Hormones’ Circulating Levels
4.5. Plasma Fatty Acid Profile
4.6. Plasma Coenzyme Q Determination
4.7. Statistical Analysis
Acknowledgments
Author Contributions
Conflicts of Interest
Abbreviations
| ACTH | Adrenocorticotropin |
| BMC | Bone mineral content |
| BMD | Bone mineral density |
| CoQ10 | Coenzyme Q10 |
| DXA | Dual energy X-ray absorptiometry |
| EDTA | Ethylenediaminetetraacetic acid |
| HED | Human equivalent dose |
| HPLC | High-performance liquid chromatography |
| MUFA | Monounsaturated fatty acids |
| n-3 PUFA | n-3 polyunsaturated fatty acids |
| n-6 PUFA | n-6 polyunsaturated fatty acids |
| NF-κB | Nuclear factor κB |
| OPG | Osteoprotegerin |
| PTH | Parathyroid hormone |
| PUFA | Polyunsaturated fatty acids |
| RANKL | Receptor activator of nuclear factor κB ligand |
| S | Sunflower oil group |
| SQ | Sunflower oil + CoQ10 group |
| V | Virgin olive oil group |
| VQ | Virgin olive oil + CoQ10 group |
| WHO | World Health Organization |
References
- Sowers, M. Clinical epidemiology and osteoporosis. Measures and their interpretation. Endocrinol. Metab. Clin. 1997, 26, 219–231. [Google Scholar] [CrossRef]
- Szulc, P.; Seeman, E.; Duboeuf, F.; Sornay-Rendu, E.; Delmas, P.D. Bone fragility: Failure of periosteal apposition to compensate for increased endocortical resorption in postmenopausal women. J. Bone Miner. Res. 2006, 21, 1856–1863. [Google Scholar] [CrossRef] [PubMed]
- Lauretani, F.; Bandinelli, S.; Griswold, M.E.; Maggio, M.; Semba, R.; Guralnik, J.M.; Ferrucci, L. Longitudinal changes in BMD and bone geometry in a population-based study. J. Bone Miner. Res. 2008, 23, 400–408. [Google Scholar] [CrossRef] [PubMed]
- Edwards, M.H.; Dennison, E.M.; Aihie Sayer, A.; Fielding, R.; Cooper, C. Osteoporosis and sarcopenia in older age. Bone 2015, 80, 126–130. [Google Scholar] [CrossRef] [PubMed]
- National Institute of Health. Consensus development panel on osteoporosis prevention, diagnosis, and therapy osteoporosis prevention, diagnosis, and therapy. J. Am. Med. Assoc. 2001, 285, 785–795. [Google Scholar]
- Willson, T.; Nelson, S.D.; Newbold, J.; Nelson, R.E.; LaFleur, J. The clinical epidemiology of male osteoporosis: A review of the recent literature. Clin. Epidemiol. 2015, 7, 65–76. [Google Scholar] [PubMed]
- Chin, K.-Y.; Ima-Nirwana, S. Olives and bone: A green osteoporosis prevention option. Int. J. Environ. Res. Public Health 2016, 13, E755. [Google Scholar] [CrossRef] [PubMed]
- Sacco, S.M.; Horcajada, M.-N.; Offord, E. Phytonutrients for bone health during ageing. Br. J. Clin. Pharmacol. 2013, 75, 697–707. [Google Scholar] [CrossRef] [PubMed]
- Penrod, J.D.; Litke, A.; Hawkes, W.G.; Magaziner, J.; Doucette, J.T.; Koval, K.J.; Silberzweig, S.B.; Egol, K.A.; Siu, A.L. The association of race, gender, and comorbidity with mortality and function after hip fracture. J. Gerontol. A Biol. Sci. Med. Sci. 2008, 63, 867–872. [Google Scholar] [CrossRef] [PubMed]
- Endo, Y.; Aharonoff, G.B.; Zuckerman, J.D.; Egol, K.A.; Koval, K.J. Gender differences in patients with hip fracture: A greater risk of morbidity and mortality in men. J. Orthop. Trauma 2005, 19, 29–35. [Google Scholar] [CrossRef] [PubMed]
- Pasco, J.A.; Henry, M.J.; Wilkinson, L.K.; Nicholson, G.C.; Schneider, H.G.; Kotowicz, M.A. Antioxidant vitamin supplements and markers of bone turnover in a community sample of nonsmoking women. J. Women’s Health 2006, 15, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Sanders, K.M.; Kotowicz, M.A.; Nicholson, G.C. Potential role of the antioxidant N-acetylcysteine in slowing bone resorption in early post-menopausal women: A pilot study. Transl. Res. J. Lab. Clin. Med. 2007, 150, 215. [Google Scholar] [CrossRef] [PubMed]
- Corwin, R.L.; Hartman, T.J.; Maczuga, S.A.; Graubard, B.I. Dietary saturated fat intake is inversely associated with bone density in humans: Analysis of NHANES III. J. Nutr. 2006, 136, 159–165. [Google Scholar] [PubMed]
- Kato, I.; Toniolo, P.; Zeleniuch-Jacquotte, A.; Shore, R.E.; Koenig, K.L.; Akhmedkhanov, A.; Riboli, E. Diet, smoking and anthropometric indices and postmenopausal bone fractures: A prospective study. Int. J. Epidemiol. 2000, 29, 85–92. [Google Scholar] [CrossRef] [PubMed]
- Järvinen, R.; Tuppurainen, M.; Erkkilä, A.T.; Penttinen, P.; Kärkkäinen, M.; Salovaara, K.; Jurvelin, J.S.; Kröger, H. Associations of dietary polyunsaturated fatty acids with bone mineral density in elderly women. Eur. J. Clin. Nutr. 2012, 66, 496–503. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.C.; Claassen, N.; Smuts, C.M.; Potgieter, H.C. Correlation between essential fatty acids and parameters of bone formation and degradation. Asia Pac. J. Clin. Nutr. 1997, 6, 235–238. [Google Scholar] [PubMed]
- Kruger, M.C.; Schollum, L.M. Is docosahexaenoic acid more effective than eicosapentaenoic acid for increasing calcium bioavailability? Prostaglandins Leukot. Essent. Fatty Acids 2005, 73, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Macdonald, H.M.; New, S.A.; Golden, M.H.N.; Campbell, M.K.; Reid, D.M. Nutritional associations with bone loss during the menopausal transition: Evidence of a beneficial effect of calcium, alcohol, and fruit and vegetable nutrients and of a detrimental effect of fatty acids. Am. J. Clin. Nutr. 2004, 79, 155–165. [Google Scholar] [PubMed]
- Trichopoulou, A.; Georgiou, E.; Bassiakos, Y.; Lipworth, L.; Lagiou, P.; Proukakis, C.; Trichopoulos, D. Energy intake and monounsaturated fat in relation to bone mineral density among women and men in Greece. Prev. Med. 1997, 26, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Harris, M.; Farrell, V.; Houtkooper, L.; Going, S.; Lohman, T. Associations of polyunsaturated fatty acid intake with bone mineral density in postmenopausal women. J. Osteoporos. 2015, 2015, 737521. [Google Scholar] [CrossRef] [PubMed]
- Poulsen, R.C.; Kruger, M.C. Detrimental effect of eicosapentaenoic acid supplementation on bone following ovariectomy in rats. Prostaglandins Leukot. Essent. Fatty Acids 2006, 75, 419–427. [Google Scholar] [CrossRef] [PubMed]
- Muraki, S.; Yamamoto, S.; Ishibashi, H.; Oka, H.; Yoshimura, N.; Kawaguchi, H.; Nakamura, K. Diet and lifestyle associated with increased bone mineral density: Cross-sectional study of Japanese elderly women at an osteoporosis outpatient clinic. J. Orthop. Sci. 2007, 12, 317–320. [Google Scholar] [CrossRef] [PubMed]
- Virtanen, J.K.; Mozaffarian, D.; Cauley, J.A.; Mukamal, K.J.; Robbins, J.; Siscovick, D.S. Fish consumption, bone mineral density, and risk of hip fracture among older adults: The cardiovascular health study. J. Bone Miner. Res. 2010, 25, 1972–1979. [Google Scholar] [CrossRef] [PubMed]
- Almeida, M.; Han, L.; Martin-Millan, M.; Plotkin, L.I.; Stewart, S.A.; Roberson, P.K.; Kousteni, S.; O’Brien, C.A.; Bellido, T.; Parfitt, A.M.; et al. Skeletal involution by age-associated oxidative stress and its acceleration by loss of sex steroids. J. Biol. Chem. 2007, 282, 27285–27297. [Google Scholar] [CrossRef] [PubMed]
- Liu, H.; Huang, H.; Li, B.; Wu, D.; Wang, F.; Zheng, X.H.; Chen, Q.; Wu, B.; Fan, X. Olive oil in the prevention and treatment of osteoporosis after artificial menopause. Clin. Interv. Aging 2014, 9, 2087–2095. [Google Scholar] [CrossRef] [PubMed]
- Bullon, P.; Battino, M.; Varela-Lopez, A.; Perez-Lopez, P.; Granados-Principal, S.; Ramirez-Tortosa, M.C.; Ochoa, J.J.; Cordero, M.D.; Gonzalez-Alonso, A.; Ramirez-Tortosa, C.L.; et al. Diets based on virgin olive oil or fish oil but not on sunflower oil prevent age-related alveolar bone resorption by mitochondrial-related mechanisms. PLoS ONE 2013, 8, e74234. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.L.; Ochoa, J.J.; Ramirez-Tortosa, C.; Battino, M.; Huertas, J.R.; Martín, Y.; Mataix, J. Dietary fat type (virgin olive vs. sunflower oils) affects age-related changes in DNA double-strand-breaks, antioxidant capacity and blood lipids in rats. Exp. Gerontol. 2004, 39, 1189–1198. [Google Scholar] [CrossRef] [PubMed]
- Varela-Lopez, A.; Bullon, P.; Battino, M.; Ramirez-Tortosa, M.C.; Ochoa, J.J.; Cordero, M.D.; Ramirez-Tortosa, C.L.; Rubini, C.; Zizzi, A.; Quiles, J.L. Coenzyme Q protects against age-related alveolar bone loss associated to n-6 polyunsaturated fatty acid rich-diets by modulating mitochondrial mechanisms. J. Gerontol. Ser. A 2016, 71, 593–600. [Google Scholar] [CrossRef] [PubMed]
- Ochoa, J.J.; Quiles, J.L.; López-Frías, M.; Huertas, J.R.; Mataix, J. Effect of lifelong coenzyme Q10 supplementation on age-related oxidative stress and mitochondrial function in liver and skeletal muscle of rats fed on a polyunsaturated fatty acid (PUFA)-rich diet. J. Gerontol. Ser. A 2007, 62, 1211–1218. [Google Scholar] [CrossRef]
- Quiles, J.L.; Pamplona, R.; Ramirez-Tortosa, M.C.; Naudí, A.; Portero-Otin, M.; Araujo-Nepomuceno, E.; López-Frías, M.; Battino, M.; Ochoa, J.J. Coenzyme Q addition to an n-6 PUFA-rich diet resembles benefits on age-related mitochondrial DNA deletion and oxidative stress of a MUFA-rich diet in rat heart. Mech. Ageing Dev. 2010, 131, 38–47. [Google Scholar] [CrossRef] [PubMed]
- Farina, E.K.; Kiel, D.P.; Roubenoff, R.; Schaefer, E.J.; Cupples, L.A.; Tucker, K.L. Protective effects of fish intake and interactive effects of long-chain polyunsaturated fatty acid intakes on hip bone mineral density in older adults: The framingham osteoporosis study. Am. J. Clin. Nutr. 2011, 93, 1142–1151. [Google Scholar] [CrossRef] [PubMed]
- Weiss, L.A.; Barrett-Connor, E.; von Mühlen, D. Ratio of n-6 to n-3 fatty acids and bone mineral density in older adults: The rancho bernardo study. Am. J. Clin. Nutr. 2005, 81, 934–938. [Google Scholar] [PubMed]
- Kanis, J.A.; Reginster, J.-Y. European guidance for the diagnosis and management of osteoporosis in postmenopausal women—What is the current message for clinical practice? Pol. Arch. Med. Wewn. 2008, 118, 538–540. [Google Scholar] [PubMed]
- Zhang, X.-X.; Qian, K.-J.; Zhang, Y.; Wang, Z.-J.; Yu, Y.-B.; Liu, X.-J.; Cao, X.-T.; Liao, Y.-H.; Zhang, D.-Y. Efficacy of coenzyme Q10 in mitigating spinal cord injury-induced osteoporosis. Mol. Med. Rep. 2015, 12, 3909–3915. [Google Scholar] [CrossRef] [PubMed]
- Varela-López, A.; Giampieri, F.; Battino, M.; Quiles, J.L. Coenzyme Q and its role in the dietary therapy against aging. Molecules 2016, 21, 373. [Google Scholar] [CrossRef] [PubMed]
- Das, U.N. Essential fatty acids and osteoporosis. Nutrition 2000, 16, 386–390. [Google Scholar] [CrossRef]
- Iwadate, H.; Kobayashi, H.; Kanno, T.; Asano, T.; Saito, R.; Sato, S.; Suzuki, E.; Watanabe, H.; Ohira, H. Plasma osteopontin is correlated with bone resorption markers in rheumatoid arthritis patients. Int. J. Rheum Dis. 2014, 17, 50–56. [Google Scholar] [CrossRef] [PubMed]
- Gravallese, E.M. Osteopontin: A bridge between bone and the immune system. J. Clin. Investig. 2003, 112, 147–149. [Google Scholar] [CrossRef] [PubMed]
- Chen, A.B.; Minami, K.; Raposo, J.F.; Matsuura, N.; Koizumi, M.; Yokota, H.; Ferreira, H.G. Transient modulation of calcium and parathyroid hormone stimulates bone formation. Endocrine 2016, 54, 232–240. [Google Scholar] [CrossRef] [PubMed]
- Nakagawa, N.; Kinosaki, M.; Yamaguchi, K.; Shima, N.; Yasuda, H.; Yano, K.; Morinaga, T.; Higashio, K. RANK is the essential signaling receptor for osteoclast differentiation factor in osteoclastogenesis. Biochem. Biophys. Res. Commun. 1998, 253, 395–400. [Google Scholar] [CrossRef] [PubMed]
- Yasuda, H.; Shima, N.; Nakagawa, N.; Yamaguchi, K.; Kinosaki, M.; Goto, M.; Mochizuki, S.I.; Tsuda, E.; Morinaga, T.; Udagawa, N.; et al. A novel molecular mechanism modulating osteoclast differentiation and function. Bone 1999, 25, 109–113. [Google Scholar] [CrossRef]
- Liu, J.M.; Zhao, H.Y.; Ning, G.; Zhao, Y.J.; Chen, Y.; Zhang, Z.; Sun, L.H.; Xu, M.-Y.; Chen, J.L. Relationships between the changes of serum levels of OPG and RANKL with age, menopause, bone biochemical markers and bone mineral density in Chinese women aged 20–75. Calcif. Tissue Int. 2005, 76, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Altindag, O.; Erel, O.; Soran, N.; Celik, H.; Selek, S. Total oxidative/anti-oxidative status and relation to bone mineral density in osteoporosis. Rheumatol. Int. 2008, 28, 317–321. [Google Scholar] [CrossRef] [PubMed]
- Sharma, T.; Islam, N.; Ahmad, J.; Akhtar, N.; Beg, M. Correlation between bone mineral density and oxidative stress in postmenopausal women. Indian J. Endocrinol. Metab. 2015, 19, 491–497. [Google Scholar] [PubMed]
- Fatokun, A.A.; Stone, T.W.; Smith, R.A. Responses of differentiated MC3T3-E1 osteoblast-like cells to reactive oxygen species. Eur. J. Pharmacol. 2008, 587, 35–41. [Google Scholar] [CrossRef] [PubMed]
- Ha, H.; Kwak, H.B.; Lee, S.W.; Jin, H.M.; Kim, H.-M.; Kim, H.-H.; Lee, Z.H. Reactive oxygen species mediate RANK signaling in osteoclasts. Exp. Cell Res. 2004, 301, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.-J.; Ko, W.-K.; Han, S.W.; Kim, D.-S.; Hwang, Y.-S.; Park, H.-K.; Kwon, I.K. Antioxidants, like coenzyme Q10, selenite, and curcumin, inhibited osteoclast differentiation by suppressing reactive oxygen species generation. Biochem. Biophys. Res. Commun. 2012, 418, 247–253. [Google Scholar] [CrossRef] [PubMed]
- Moon, H.-J.; Ko, W.-K.; Jung, M.-S.; Kim, J.H.; Lee, W.-J.; Park, K.-S.; Heo, J.-K.; Bang, J.B.; Kwon, I.K. Coenzyme Q10 regulates osteoclast and osteoblast differentiation. J. Food Sci. 2013, 78, H785–H891. [Google Scholar] [CrossRef] [PubMed]
- Bello, R.I.; Gómez-Díaz, C.; Burón, M.I.; Alcaín, F.J.; Navas, P.; Villalba, J.M. Enhanced anti-oxidant protection of liver membranes in long-lived rats fed on a coenzyme Q10-supplemented diet. Exp. Gerontol. 2005, 40, 694–706. [Google Scholar] [CrossRef] [PubMed]
- Reeves, P.G. Components of the AIN-93 diets as improvements in the AIN-76A diet. J. Nutr. 1997, 127, 838S–841S. [Google Scholar] [PubMed]
- Quiles, J.L.; Huertas, J.R.; Mañas, M.; Ochoa, J.J.; Battino, M.; Mataix, J. Oxidative stress induced by exercise and dietary fat modulates the coenzyme Q and vitamin A balance between plasma and mitochondria. Int. J. Vitam. Nutr. Res. 1999, 69, 243–249. [Google Scholar] [CrossRef] [PubMed]
- Quiles, J.L.; Ochoa, J.J.; Ramirez-Tortosa, M.C.; Linde, J.; Bompadre, S.; Battino, M.; Narbona, E.; Maldonado, J.; Mataix, J. Coenzyme Q concentration and total antioxidant capacity of human milk at different stages of lactation in mothers of preterm and full-term infants. Free Radic. Res. 2006, 40, 199–206. [Google Scholar] [CrossRef] [PubMed]
© 2017 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Varela-López, A.; Ochoa, J.J.; Llamas-Elvira, J.M.; López-Frías, M.; Planells, E.; Speranza, L.; Battino, M.; Quiles, J.L. Loss of Bone Mineral Density Associated with Age in Male Rats Fed on Sunflower Oil Is Avoided by Virgin Olive Oil Intake or Coenzyme Q Supplementation. Int. J. Mol. Sci. 2017, 18, 1397. https://doi.org/10.3390/ijms18071397
Varela-López A, Ochoa JJ, Llamas-Elvira JM, López-Frías M, Planells E, Speranza L, Battino M, Quiles JL. Loss of Bone Mineral Density Associated with Age in Male Rats Fed on Sunflower Oil Is Avoided by Virgin Olive Oil Intake or Coenzyme Q Supplementation. International Journal of Molecular Sciences. 2017; 18(7):1397. https://doi.org/10.3390/ijms18071397
Chicago/Turabian StyleVarela-López, Alfonso, Julio J. Ochoa, José M. Llamas-Elvira, Magdalena López-Frías, Elena Planells, Lorenza Speranza, Maurizio Battino, and José L. Quiles. 2017. "Loss of Bone Mineral Density Associated with Age in Male Rats Fed on Sunflower Oil Is Avoided by Virgin Olive Oil Intake or Coenzyme Q Supplementation" International Journal of Molecular Sciences 18, no. 7: 1397. https://doi.org/10.3390/ijms18071397