Estimation of Whole Plant Photosynthetic Rate of Irwin Mango under Artificial and Natural Lights Using a Three-Dimensional Plant Model and Ray-Tracing

Abstract
:
1. Introduction
2. Results
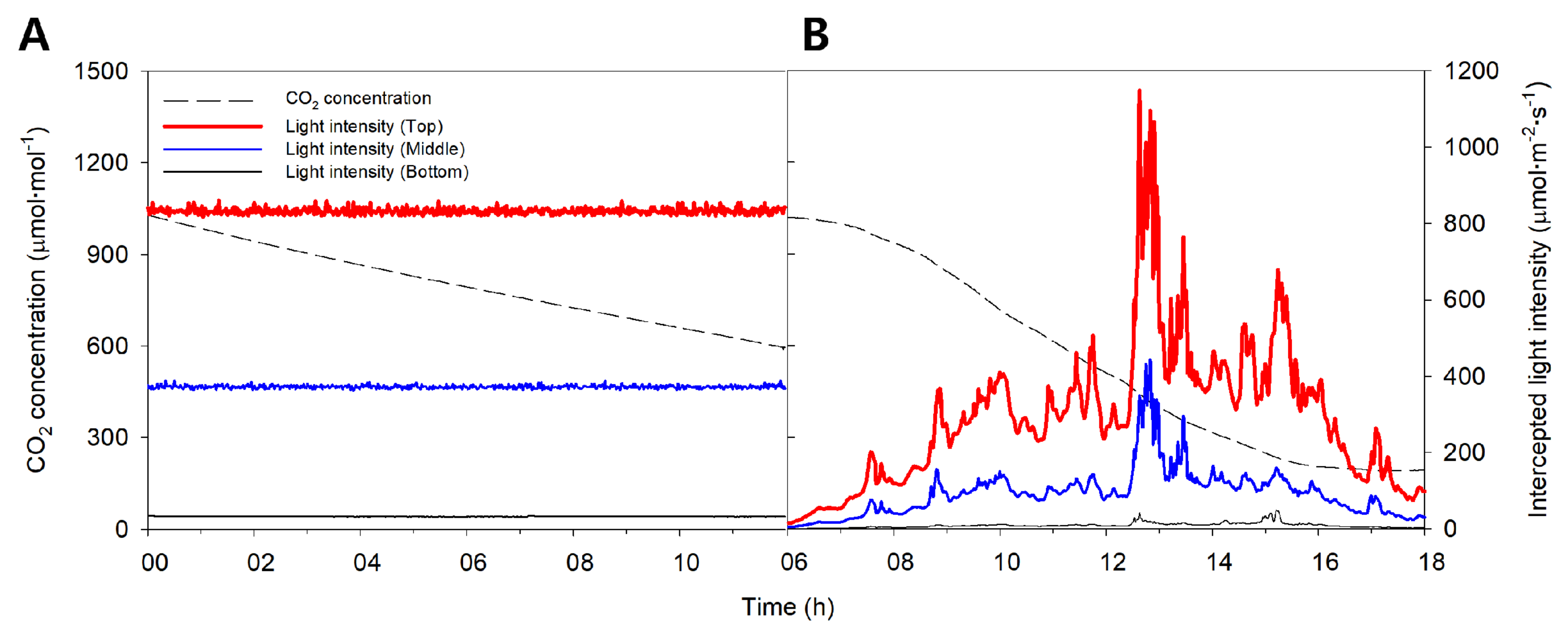
2.1. Actual Distribution of Intercepted Light Intensity and Measured Whole Plant Photosynthetic Rate
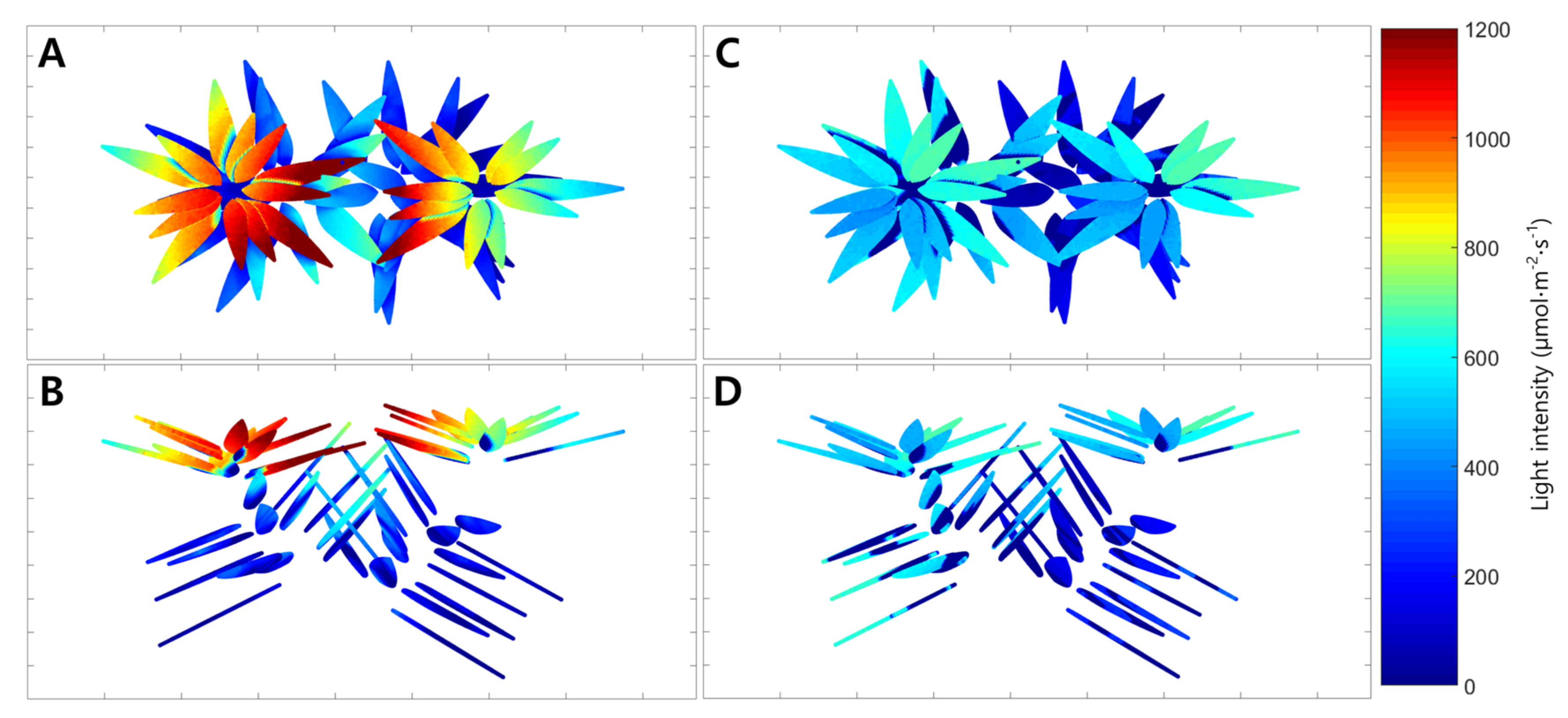
2.2. Simulated Distribution of Intercepted Light Intensity and Estimated Whole Plant Photosynthetic Rate
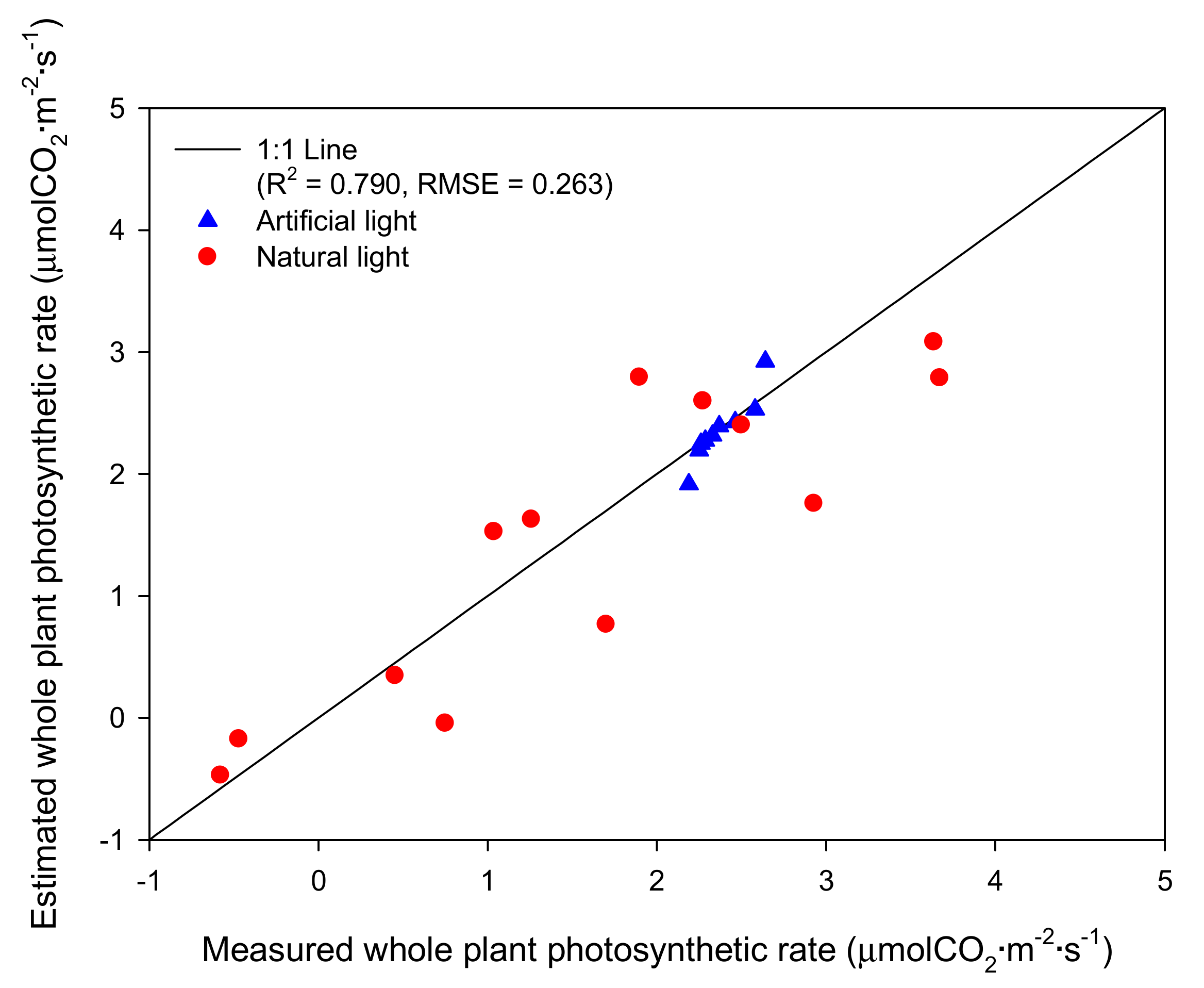
2.3. Validation of the Intercepted Light Intensity and Whole Plant Photosynthetic Rate
3. Discussion
4. Materials and Methods
4.1. Measurement of Photosynthetic Rates of Irwin Mango
4.2. Construction of the 3D Plant Model
4.3. Simulation of the Intercepted Light Intensity
4.4. Calculation of the Whole Plant Photosynthetic Rate
4.5. Validation of the Intercepted Light Intensity and Whole Plant Photosynthetic Rate
5. Conclusions
Acknowledgments
Authors Contribution
Conflicts of Interest
References
- Monteith, J.L. Light distribution and photosynthesis in field crops. Ann. Bot. 1965, 29, 17–37. [Google Scholar] [CrossRef]
- Hu, E.; Tong, L.; Hu, D.; Liu, H. Mixed effects of CO2 concentration on photosynthesis of lettuce in a closed artificial ecosystem. Ecol. Eng. 2011, 37, 2082–2086. [Google Scholar] [CrossRef]
- Jung, D.H.; Kim, D.; Yoon, H.I.; Moon, T.W.; Park, K.S.; Son, J.E. Modelling the canopy photosynthetic rate of romaine lettuce (Lactuca sativa L.) grown in a plant factory at varying CO2 concentrations and growth stages. Hortic. Environ. Biotechnol. 2016, 57, 487–492. [Google Scholar] [CrossRef]
- Son, J.E.; Park, J.S.; Park, H.Y. Analysis of carbon dioxide changes in urban-type plant factory system. Hortic. Environ. Biotechnol. 1999, 40, 205–208. [Google Scholar]
- Chen, J.M.; Liu, J.; Cihlar, J.; Goulden, M.L. Daily canopy photosynthesis model through temporal and spatial scaling for remote sensing applications. Ecol. Model. 1999, 124, 99–119. [Google Scholar] [CrossRef]
- Vos, J.; Evers, J.B.; Buck-Sorlin, G.H.; Andrieu, B.; Chelle, M.; de Visser, P.H.B. Functional-structural plant modelling: A new versatile tool in crop science. J. Exp. Bot. 2010, 61, 2101–2115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Buck-Sorlin, G.; de Visser, P.H.B.; Henke, M.; Sarlikioti, V.; van der Heijden, G.W.A.M.; Marcelis, L.F.M.; Vos, J. Towards a functional–structural plant model of cut-rose: Simulation of light environment, light absorption, photosynthesis and interference with the plant structure. Ann. Bot. 2011, 108, 1121–1134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leith, J.H.; Pasian, C.C. A model of net photosynthesis of rose leaves as a function of photosynthetically active radiation, leaf temperature, and leaf age. J. Am. Soc. Hortic. Sci. 1990, 115, 486–491. [Google Scholar]
- Stirling, C.M.; Aguilera, C.; Baker, N.R.; Long, S.P. Changes in the photosynthetic light response curve during leaf development of field grown maize with implications for modelling canopy photosynthesis. Photosynth. Res. 1994, 42, 217–225. [Google Scholar] [CrossRef] [PubMed]
- Yonemoto, Y. Tropical fruit. In Horticulture in Japan; The Japanese Society for Horticultural Science, Ed.; Shoukadoh Publication: Kyoto, Japan, 2006; pp. 100–106. [Google Scholar]
- Farquhar, G.D.; von Caemmerer, S.; Berry, J.A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta 1980, 149, 78–90. [Google Scholar] [CrossRef] [PubMed]
- Zhu, X.G.; Song, Q.; Ort, D.R. Elements of a dynamic systems model of canopy photosynthesis. Curr. Opin. Plant Biol. 2012, 15, 237–244. [Google Scholar] [CrossRef] [PubMed]
- De Pury, D.G.G.; Farquhar, G.D. Simple scaling of photosynthesis from leaves to canopies without the errors of big-leaf models. Plant Cell Environ. 1997, 20, 537–557. [Google Scholar] [CrossRef]
- Johnson, I.R.; Thornley, J.H.M.; Frantz, J.M.; Bugbee, B. A model of canopy photosynthesis incorporating protein distribution through the canopy and its acclimation to light, temperature and CO2. Ann. Bot. 2010, 106, 735–749. [Google Scholar] [CrossRef] [PubMed]
- Monsi, M.; Saeki, T. The light factor in plant communities and its significance for dry matter production. J. Bot. 1953, 14, 22–52. [Google Scholar]
- Norman, J.M. Interfacing Leaf and Canopy Light Interception Models in Predicting Photosynthesis for Ecosystem Models; CRC Press: Boca Raton, FL, USA, 1980; Volume 2, pp. 49–67. [Google Scholar]
- Hikosaka, K. Optimal nitrogen distribution within a leaf canopy under direct and diffuse light. Plant Cell Environ. 2014, 37, 2077–2085. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.S.; Palmroth, S.; Thérezién, M.; Stenberg, P.; Oren, R. Analysis of the sensitivity of absorbed light and incident light profile to various canopy architecture and stand conditions. Tree Physiol. 2010, 31, 30–47. [Google Scholar] [CrossRef] [PubMed]
- Tang, L.; Hou, C.; Huang, H.; Chen, C.; Zou, J.; Lin, D. Light interception efficiency analysis based on three-dimensional peach canopy models. Ecol. Inform. 2015, 30, 60–67. [Google Scholar] [CrossRef]
- Yin, X.; Struik, P.C. C3 and C4 photosynthesis models: An overview from the perspective of crop modelling. NJAS Wagening. J. Life Sci. 2009, 57, 27–38. [Google Scholar] [CrossRef]
- Jung, D.H.; Shin, J.H.; Cho, Y.Y.; Son, J.E. Development of a two-variable spatial leaf photosynthetic model of Irwin mango grown in greenhouse. Prot. Hortic. Plant Fact. 2015, 24, 161–166. [Google Scholar] [CrossRef]
- Schaffer, B.; Whiley, A.W.; Searle, C.; Nissen, R.J. Leaf gas exchange, dry matter partitioning, and mineral element concentrations in mangos as influenced by elevated atmospheric carbon dioxide and root restriction. J. Am. Soc. Hortic. Sci. 1997, 122, 849–855. [Google Scholar]
- Chen, T.W.; Nguyen, T.M.; Kahlen, K.; Stützel, H. High temperature and vapor pressure deficit aggravate architectural effects but ameliorate non-architectural effects of salinity on dry mass production of tomato. Front. Plant Sci. 2015, 6, 887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De Visser, P.H.B.; Buck-Sorlin, G.H.; van der Heijden, G.W.A.M. Optimizing illumination in the greenhouse using a 3D model of tomato and a ray tracer. Front. Plant Sci. 2014, 5, 48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Der Zande, D.V.; Struckens, J.; Verstraeten, W.W.; Mereu, S.; Muys, B.; Coppin, P. 3D modelling of light interception in heterogeneous forest canopies using ground-based LiDAR data. Int. J. Appl. Earth Obs. Geo Inf. 2011, 13, 792–800. [Google Scholar] [CrossRef]
- Kim, J.H.; Lee, J.W.; Ahn, T.I.; Shin, J.H.; Park, K.S.; Son, J.E. Sweet pepper (Capsicum annuum L.) canopy photosynthesis modelling using 3D plant architecture and light ray-tracing. Front. Plant Sci. 2016, 7, 1321. [Google Scholar] [CrossRef] [PubMed]
- Gonzalez-Real, M.M.; Baille, A. Changes in leaf photosynthetic parameters with leaf position and nitrogen content within a rose plant canopy (Rosa hybrid). Plant Cell Environ. 2000, 23, 351–363. [Google Scholar] [CrossRef]
- Sarlikioti, V.; de Visser, P.H.B.; Buck-Sorlin, G.H.; Marcelis, L.F.M. How plant architecture affects light absorption and photosynthesis in tomato: Towards an ideotype for plant architecture using a functional-structural plant model. Ann. Bot. 2011, 108, 1065–1073. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, M.; Xu, X.; Chen, X.; Xue, L.; Cao, B. Cucumber carbohydrate metabolism and translocation under chilling night temperature. J. Plant Physiol. 2007, 164, 621–628. [Google Scholar] [CrossRef] [PubMed]
- Bugbee, B. Steady-state canopy gas exchange: System design and operation. HortScience 1992, 27, 770–776. [Google Scholar] [PubMed]
- Takahashi, N.; Ling, P.P.; Frantz, J.M. Considerations for accurate whole plant photosynthesis measurement. Environ. Control Biol. 2008, 46, 91–101. [Google Scholar] [CrossRef]
- Prusinkiewicz, P. Modelling plant growth and development. Curr. Opin. Plant Biol. 2004, 7, 79–83. [Google Scholar] [CrossRef] [PubMed]
- Prior, L.D.; Eamus, D.; Duff, G.A. Seasonal and diurnal patterns of carbon assimilation, stomatal conductance and leaf water potential in Eucalyptus tetrodonta saplings in a wet-dry savanna in northern Australia. Aust. J. Bot. 1997, 45, 241–258. [Google Scholar] [CrossRef]
- Flanagan, L.B.; Ehleringer, J.R. Effects of mild water stress and diurnal changes in temperature and humidity on the stable oxygen and hydrogen isotopic composition of leaf water in Cornus stolonifera L. Plant Physiol. 1991, 97, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R. Photosynthetic acclimation and nitrogen partitioning within a Lucerne canopy. I. Canopy characteristics. Aust. J. Plant Physiol. 1993, 20, 55–67. [Google Scholar] [CrossRef]
- Osborne, C.P.; LaRoche, J.; Garcia, R.L.; Kimball, B.A.; Wall, G.W.; Pinter, P.J., Jr.; LaMorte, R.L.; Hendrey, G.R.; Long, S.P. Does leaf position within a canopy affect acclimation of photosynthesis to elevated CO2? Plant Physiol. 1998, 117, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Del Pozo, A.; Perez, P.; Gutiérrez, D.; Alonso, A.; Morcuende, R.; Martinez-Carrasco, R. Gas exchange acclimation to elevated CO2 in upper-sunlit and lower-shaded canopy leaves in relation to nitrogen acquisition and partitioning in wheat grown in field chambers. Environ. Exp. Bot. 2007, 59, 371–380. [Google Scholar] [CrossRef]
- Thornley, J.H.M. Acclimation of photosynthesis to light and canopy nitrogen distribution: An interpretation. Ann. Bot. 2004, 93, 473–475. [Google Scholar] [CrossRef] [PubMed]
- Marcelis, L.F.M.; Heuvelink, E.; Goudriaan, J. Modelling biomass production and yield of horticultural crops: A review. Sci. Hortic. 1998, 74, 83–111. [Google Scholar] [CrossRef]
- Jung, D.H.; Cho, Y.Y.; Lee, J.G.; Son, J.E. Estimation of leaf area, leaf fresh weight, and leaf dry weight of Irwin mango grown in greenhouse using leaf length, leaf width, petiole length, and SPAD value. Prot. Hortic. Plant Fact. 2016, 25, 146–152. [Google Scholar] [CrossRef]
- Whiley, A.W.; Schaffer, B. Stress physiology of mango. In The Mango: Botany, Production and Uses; Litz, R.E., Ed.; CAB International Press: London, UK, 1997; pp. 147–173. [Google Scholar]
- Chamchaiyaporn, T.; Jutamanee, K.; Kasemsap, P.; Vaithanomsat, P. Selection of the most appropriate coating particle film for improving photosynthesis in mango. Kasetsart J. (Nat. Sci.) 2013, 47, 323–332. [Google Scholar]
- Jones, J.W.; Dayan, E.; Allen, L.H.; van Keulen, H.; Challa, H. A dynamic tomato growth and yield model (TOMGRO). Trans. ASAE 1991, 34, 663–672. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Leaf Number | Angle (°) |
|---|---|
| 1–2 | 151.5 ± 4.75 |
| 2–3 | 133.2 ± 5.20 |
| 3–4 | 141.5 ± 11.80 |
| 4–5 | 157.6 ± 7.05 |
| 5–6 | 129.9 ± 0.25 |
| 6–7 | 122.1 ± 0.50 |
| 7–8 | 138.0 ± 6.75 |
| 8–9 | 138.8 ± 3.00 |
| 9–10 | 124.4 ± 10.25 |
| 10–11 | 150.9 ± 2.15 |
| 11–12 | 125.9 ± 18.40 |
| Average | 137.6 ± 13.38 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jung, D.H.; Lee, J.W.; Kang, W.H.; Hwang, I.H.; Son, J.E. Estimation of Whole Plant Photosynthetic Rate of Irwin Mango under Artificial and Natural Lights Using a Three-Dimensional Plant Model and Ray-Tracing. Int. J. Mol. Sci. 2018, 19, 152. https://doi.org/10.3390/ijms19010152
Jung DH, Lee JW, Kang WH, Hwang IH, Son JE. Estimation of Whole Plant Photosynthetic Rate of Irwin Mango under Artificial and Natural Lights Using a Three-Dimensional Plant Model and Ray-Tracing. International Journal of Molecular Sciences. 2018; 19(1):152. https://doi.org/10.3390/ijms19010152
Chicago/Turabian StyleJung, Dae Ho, Joon Woo Lee, Woo Hyun Kang, In Ha Hwang, and Jung Eek Son. 2018. "Estimation of Whole Plant Photosynthetic Rate of Irwin Mango under Artificial and Natural Lights Using a Three-Dimensional Plant Model and Ray-Tracing" International Journal of Molecular Sciences 19, no. 1: 152. https://doi.org/10.3390/ijms19010152