Differential Accumulation of Anthocyanins in Dendrobium officinale Stems with Red and Green Peels

 , and
, and
Abstract
:1. Introduction
2. Results and Discussion
2.1. Red D. officinale Stems Exhibit High Antioxidant Activity
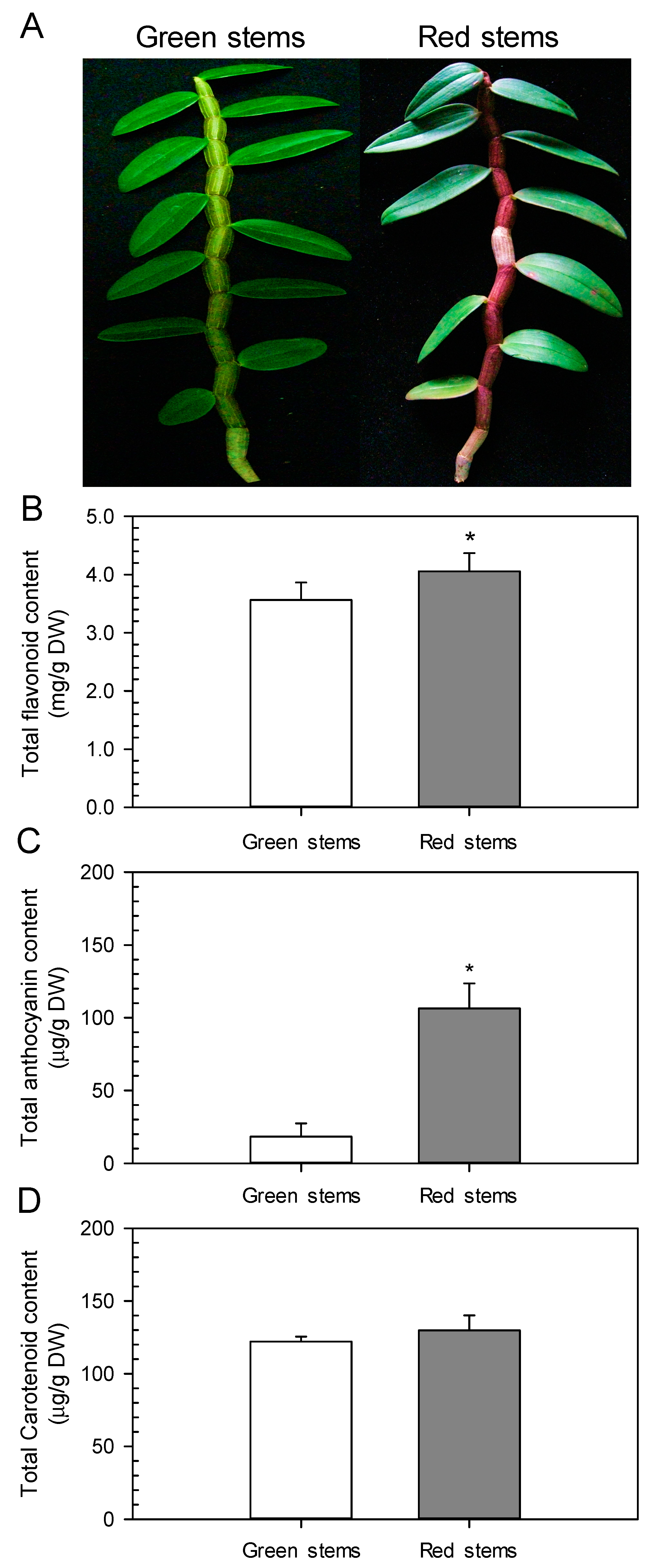
2.2. Red D. officinale Stems Possess High Anthocyanin Content
2.3. The Red Color of Stem Peels Is Mainly Localized in Epidermal Cells
2.4. Red-Peel Color Was Determined by Delphinidin and Cyanidin Derivatives of Anthocyanins
2.5. DoANS and DoUFGT Are Vital Regulatory Genes during Accumulation of Anthocyanidins
3. Materials and Methods
3.1. Chemicals and Regents
3.2. Plant Materials
3.3. Assessment of Total Flavonoids, Anthocyanins and Carotenoids Content in D. officinale Stems
3.4. Compositional Analysis of Flavonoids and Anthocyanins by UPLC-QTOF-MS
3.5. Evaluation of Antioxidant Activity In Vitro
3.6. Light Microscopy of D. officinale Stem
3.7. Gene Expression Analysis by Quantitative Real-Time PCR
3.8. Cloning of Genes and Phylogenetic Analysis
3.9. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| AA | ascorbic acid |
| ANS | anthocyanin synthase |
| 4CL | 4-coumarate CoA ligase |
| C4H | cinnamate 4-hydroxylase |
| CHI | chalcone isomerase |
| CHS | chalcone synthase |
| DFR | dihydroflavonol 4-reductase |
| DoANS | gene encoding anthocyanin synthase from Dendrobium officinale |
| DoUFGT | gene encoding UDP-glucose flavonoid 3-O-glucosyltransferase from Dendrobium officinale |
| DPPH | 2,2-diphenyl-1-picryhydrazyl |
| DW | dry weight |
| F3H | flavanone 3-hydroxylase |
| F3′H | flavonoid 3′-hydroxylase |
| F3′5′H | flavonoid 3′5′-hydroxylase |
| FLS | flavonol synthase |
| GC-MS | gas chromatography-mass spectrometry |
| ORF | open reading frame |
| PAL | phenylalanine ammonia lyase |
| qRT-PCR | quantitative real time polymerase chain reaction |
| SD | standard deviation |
| UFGT | UDP-glucose flavonoid 3-O-glucosyltransferase |
| UPLC-QTOF-MS | Ultra-performance liquid chromatography/quadrupole time-of-flight mass spectrometry |
References
- Ng, T.B.; Liu, J.; Wong, J.H.; Ye, X.; Wing Sze, S.C.; Tong, Y.; Zhang, K.Y. Review of research on Dendrobium, a prized folk medicine. Appl. Microbiol. Biotechnol. 2012, 93, 1795–1803. [Google Scholar] [CrossRef] [PubMed]
- Wei, W.; Feng, L.; Bao, W.R.; Ma, D.L.; Leung, C.H.; Nie, S.P.; Han, Q.B. Structure characterization and immunomodulating effects of polysaccharides isolated from Dendrobium officinale. J. Agric. Food Chem. 2016, 64, 881–889. [Google Scholar] [CrossRef] [PubMed]
- Luo, Q.L.; Tang, Z.H.; Zhang, X.F.; Zhong, Y.H.; Yao, S.Z.; Wang, L.S.; Lin, C.W.; Luo, X. Chemical properties and antioxidant activity of a water-soluble polysaccharide from Dendrobium officinale. Int. J. Biol. Macromol. 2016, 89, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Yue, H.; Liu, Y.; Qu, H.; Ding, K. Structure analysis of a novel heteroxylan from the stem of Dendrobium officinale and anti-angiogenesis activities of its sulfated derivative. Int. J. Biol. Macromol. 2017, 103, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Xie, S.Z.; Liu, B.; Zhang, D.D.; Zha, X.Q.; Pan, L.H.; Luo, J.P. Intestinal immunomodulating activity and structural characterization of a new polysaccharide from stems of Dendrobium officinale. Food Funct. 2016, 7, 2789–2799. [Google Scholar] [CrossRef] [PubMed]
- Liang, J.; Chen, S.; Chen, J.; Lin, J.; Xiong, Q.; Yang, Y.; Yuan, J.; Zhou, L.; He, L.; Hou, S.; Li, S.; et al. Therapeutic roles of polysaccharides from Dendrobium officinale on colitis and its underlying mechanisms. Carbohydr. Polym. 2018, 185, 159–168. [Google Scholar] [CrossRef] [PubMed]
- Teixeira da Silva, J.A.; Ng, T.B. The medicinal and pharmaceutical importance of Dendrobium species. Appl. Microbiol. Biotechnol. 2017, 101, 2227–2239. [Google Scholar] [CrossRef] [PubMed]
- Xing, X.; Cui, S.W.; Nie, S.; Phillips, G.O.; Douglas Goff, H.; Wang, Q. A review of isolation process, structural characteristics, and bioactivities of water-soluble polysaccharides from Dendrobium plants. Bioact. Carbohydr. Diet. Fiber 2013, 1, 131–147. [Google Scholar] [CrossRef]
- Jin, Q.; Jiao, C.; Sun, S.; Song, C.; Cai, Y.; Lin, Y.; Fan, H.; Zhu, Y. Metabolic analysis of medicinal Dendrobium officinale and Dendrobium huoshanense during different growth years. PLoS ONE 2016, 11, e0146607. [Google Scholar] [CrossRef] [PubMed]
- Chinese Pharmacopeia Commission. Pharmacopoeia of the People’s Republic of China, 10th ed.; China Medical Science Press: Beijing, China, 2015; pp. 281–283. [Google Scholar]
- Jaakola, L. New insights into the regulation of anthocyanin biosynthesis in fruits. Trends Plant Sci. 2013, 18, 477–483. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tanaka, Y.; Sasaki, N.; Ohmiya, A. Biosynthesis of plant pigments: Anthocyanins, betalains and carotenoids. Plant J. 2008, 54, 733–749. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Butelli, E.; Martin, C. Engineering anthocyanin biosynthesis in plants. Curr. Opin. Plant Biol. 2014, 19, 81–90. [Google Scholar] [CrossRef] [PubMed]
- Liew, C.F.; Loh, C.S.; Goh, C.J.; Lim, S.H. The isolation, molecular characterization and expression of dihydroflavonol 4-reductase cDNA in the orchid, Bromheadia finlaysoniana. Plant Sci. 1998, 135, 161–169. [Google Scholar] [CrossRef]
- Kriangphan, N.; Vuttipongchaikij, S.; Kittiwongwattana, C.; Suttangkakul, A.; Pinmanee, P.; Sakulsathaporn, A.; Suwimon, R.; Suputtitada, S.; Chanvivattana, Y.; Apisitwanich, S. Effects of sequence and expression of eight anthocyanin biosynthesis genes on floral coloration in four Dendrobium hybrids. Horticult. J. 2015, 84, 83–92. [Google Scholar] [CrossRef]
- Han, Y.Y.; Ming, F.; Wang, W.; Wang, J.W.; Ye, M.M.; Shen, D.L. Molecular evolution and functional specialization of chalcone synthase superfamily from Phalaenopsis orchid. Genetica 2006, 128, 429–438. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.W.; Ming, F.; Han, Y.Y.; Shen, D.L. Flavonoid-3′,5′-hydroxylase from Phalaenopsis: A novel member of cytochrome P450s, its cDNA cloning, endogenous expression and molecular modeling. Biotechnol. Lett. 2006, 28, 327–334. [Google Scholar] [CrossRef] [PubMed]
- Grotewold, E. The genetics and biochemistry of floral pigments. Annu. Rev. Plant Biol. 2006, 57, 761–780. [Google Scholar] [CrossRef] [PubMed]
- Iwashina, T. Contribution to flower colors of flavonoids including anthocyanins: A review. Nat. Prod. Commun. 2015, 10, 529–544. [Google Scholar] [PubMed]
- Braidot, E.; Zancani, M.; Petrussa, E.; Peresson, C.; Bertolini, A.; Patui, S.; Macri, F.; Vianello, A. Transport and accumulation of flavonoids in grapevine (Vitis vinifera L.). Plant Signal Behav. 2008, 3, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Vegara, S.; Mena, P.; Marti, N.; Saura, D.; Valero, M. Approaches to understanding the contribution of anthocyanins to the antioxidant capacity of pasteurized pomegranate juices. Food Chem. 2013, 141, 1630–1636. [Google Scholar] [CrossRef] [PubMed]
- Bai, S.L.; Sun, Y.W.; Qian, M.J.; Yang, F.X.; Ni, J.B.; Tao, R.Y.; Li, L.; Shu, Q.; Zhang, D.; Teng, Y.W. Transcriptome analysis of bagging-treated red Chinese sand pear peels reveals light-responsive pathway functions in anthocyanin accumulation. Sci. Rep. 2017, 7, 63. [Google Scholar] [CrossRef] [PubMed]
- Lei, Z.X.; Zhou, C.H.; Ji, X.Y.; Wei, G.; Huang, Y.C.; Yu, W.X.; Luo, Y.Y.; Qiu, Y. Transcriptome analysis reveals genes involved in flavonoid biosynthesis and accumulation in Dendrobium catenatum from different locations. Sci. Rep. 2018, 8, 6373. [Google Scholar] [CrossRef] [PubMed]
- Takashima, M.; Nara, K.; Niki, E.; Yoshida, Y.; Hagihara, Y.; Stowe, M.; Horie, M. Evaluation of biological activities of a groundnut (Apios americana Medik) extract containing a novel isoflavone. Food Chem. 2013, 138, 298–305. [Google Scholar] [CrossRef] [PubMed]
- Pápay, Z.E.; Kállai-Szabó, N.; Balogh, E.; Ludányi, K.; Klebovich, I.; Antal, I. Controlled release oral delivery of apigenin containing pellets with antioxidant activity. Curr. Drug Deliv. 2017, 14, 145–154. [Google Scholar] [CrossRef] [PubMed]
- Bartley, G.E.; Scolnik, P.A. Plant carotenoids: Pigments for photoprotection, visual attraction, and human health. Plant Cell 1995, 7, 1027–1038. [Google Scholar] [CrossRef] [PubMed]
- Yue, Q.; Xu, L.; Xiang, G.; Yu, X.; Yao, Y. Characterization of gene expression profile, phenolic composition, and antioxidant capacity in red-fleshed grape berries and their wines. J. Agric. Food Chem. 2018, 66, 7190–7199. [Google Scholar] [CrossRef] [PubMed]
- Fu, X.M.; Cheng, S.H.; Liao, Y.Y.; Huang, B.Z.; Du, B.; Zeng, W.; Jiang, Y.M.; Duan, X.W.; Yang, Z.Y. Comparative analysis of pigments in red and yellow banana fruit. Food Chem. 2018, 239, 1009–1018. [Google Scholar] [CrossRef] [PubMed]
- Sui, X.; Gao, X.; Ao, M.; Wang, Q.M.; Yang, D.; Wang, M.; Fu, Y.; Wang, L. cDNA cloning and characterization of UDP-glucose: Anthocyanidin 3-O-glucosyltransferase in Freesia hybrida. Plant Cell Rep. 2011, 30, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Hieber, A.D.; Mudalige-Jayawickrama, R.G.; Kuehnle, A.R. Color genes in the orchid Oncidium Gower Ramsey: Identification, expression, and potential genetic instability in an interspecific cross. Planta 2006, 223, 521–531. [Google Scholar] [CrossRef] [PubMed]
- Li, C.H.; Qiu, F.; Ding, L.; Huang, M.Z.; Huang, S.R.; Yang, G.S.; Yin, J.M. Anthocyanin biosynthesis regulation of DhMYB2 and DhbHLH1 in Dendrobium hybrids petals. Plant Physiol. Biochem. 2017, 112, 335–345. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; He, C.; Wu, K.; Teixeira da Silva, J.A.; Zeng, S.; Zhang, X.; Yu, Z.; Xia, H.; Duan, J. Transcriptome analysis of Dendrobium officinale and its application to the identification of genes associated with polysaccharide synthesis. Front. Plant Sci. 2016, 7, 5. [Google Scholar] [CrossRef] [PubMed]
- Wilmouth, R.C.; Turnbull, J.J.; Welford, R.W.; Clifton, I.J.; Prescott, A.G.; Schofield, C.J. Structure and mechanism of anthocyanidin synthase from Arabidopsis thaliana. Structure 2002, 10, 93–103. [Google Scholar] [CrossRef]
- Chen, M.Q.; Xu, M.Y.; Xiao, Y.; Cui, D.D.; Qin, Y.Q.; Wu, J.Q.; Wang, W.Y.; Wang, G.P. Fine mapping identifies SmFAS encoding an anthocyanidin synthase as a putative candidate gene for flower purple color in Solanum melongena L. Int. J. Mol. Sci. 2018, 19, 789. [Google Scholar] [CrossRef] [PubMed]
- Wrolstad, R.E.; Putnam, T.P.; Varseveld, G.W. Color quality of frozen strawberries: Effect of anthocyanin, pH, total acidity and ascorbic acid variability. J. Food Sci. 1970, 35, 448–452. [Google Scholar] [CrossRef]
- Lichtenthaler, H.K.; Wellburn, A.R. Determinations of total carotenoids and chlorophylls A and B of leaf extracts in different solvents. Biochem. Soc. Trans. 1983, 11, 591–592. [Google Scholar] [CrossRef]
- Luo, A.X.; He, X.J.; Zhou, S.; Fan, Y.J.; Luo, A.S.; Chun, Z. Purification, composition analysis and antioxidant activity of the polysaccharides from Dendrobium nobile Lindl. Carbohydr. Polym. 2010, 79, 1014–1019. [Google Scholar] [CrossRef]
- Yu, Z.M.; He, C.M.; Teixeira da Silva, J.A.; Zhang, G.H.; Dong, W.; Luo, J.P.; Duan, J. Molecular cloning and functional analysis of DoUGE related to water-soluble polysaccharides from Dendrobium officinale with enhanced abiotic stress tolerance. Plant Cell Tissue Organ Culture 2017, 131, 579–599. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Chenna, R.; Sugawara, H.; Koike, T.; Lopez, R.; Gibson, T.J.; Higgins, D.G.; Thompson, J.D. Multiple sequence alignment with the Clustal series of programs. Nucleic Acids Res. 2003, 31, 3497–3500. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Identification and Tentative Identification | Molecule | Rt (min) | ESI (+) MS/MS2 (m/z) | Red/Green Peak Area Ratio |
|---|---|---|---|---|---|
| 1 | Delphinidin 3,5-O-diglucoside | C27H31O17 | 3.62 | 627.1422([M]+)/465.0927([M-glu]+); 303.0477([M-2glu]+) | + |
| 2 | Peonidin 3,5-O-diglucoside | C28H33O16 | 4.54 | 625.1638([M]+)/463.0942([M-glu]+); 301.0627([M-2glu]+) | 18.66 (p = 7.64 × 10−9) |
| 3 | Cyanidin 3-O-galactoside | C21H21O11 | 4.93 | 449.1007([M]+)/287.0515([M-gala]+) | + |
| 4 | Cyanidin 3-O-glucoside | C21H21O11 | 5.07 | 449.1010([M]+)/287.0521([M-glu]+) | + |
| 5 | Pelargonidin 3,5-O-diglucoside | C27H31O15 | 5.26 | 595.1585([M]+)/433.1092([M-glu]+); 271.0602([M-glu]+) | − |
| 6 | Pelargonidin 3-O-rutinoside | C27H31O14 | 5.46 | 579.1615([M]+)/271.0587([M-rutin]+) | 33.85 (p = 1.35 × 10−6) |
| 7 | Malvidin 3-O-glucoside | C23H25O12 | 7.11 | 493.13([M]+)/331.0772([M-glu]+) | 8.53 (p = 2.28 × 10−5) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, Z.; Liao, Y.; Teixeira da Silva, J.A.; Yang, Z.; Duan, J. Differential Accumulation of Anthocyanins in Dendrobium officinale Stems with Red and Green Peels. Int. J. Mol. Sci. 2018, 19, 2857. https://doi.org/10.3390/ijms19102857
Yu Z, Liao Y, Teixeira da Silva JA, Yang Z, Duan J. Differential Accumulation of Anthocyanins in Dendrobium officinale Stems with Red and Green Peels. International Journal of Molecular Sciences. 2018; 19(10):2857. https://doi.org/10.3390/ijms19102857
Chicago/Turabian StyleYu, Zhenming, Yinyin Liao, Jaime A. Teixeira da Silva, Ziyin Yang, and Jun Duan. 2018. "Differential Accumulation of Anthocyanins in Dendrobium officinale Stems with Red and Green Peels" International Journal of Molecular Sciences 19, no. 10: 2857. https://doi.org/10.3390/ijms19102857
APA StyleYu, Z., Liao, Y., Teixeira da Silva, J. A., Yang, Z., & Duan, J. (2018). Differential Accumulation of Anthocyanins in Dendrobium officinale Stems with Red and Green Peels. International Journal of Molecular Sciences, 19(10), 2857. https://doi.org/10.3390/ijms19102857