Germination and the Early Stages of Seedling Development in Brachypodium distachyon

 ,
, 
Abstract
:1. Introduction
2. Results
2.1. Germination Characteristics
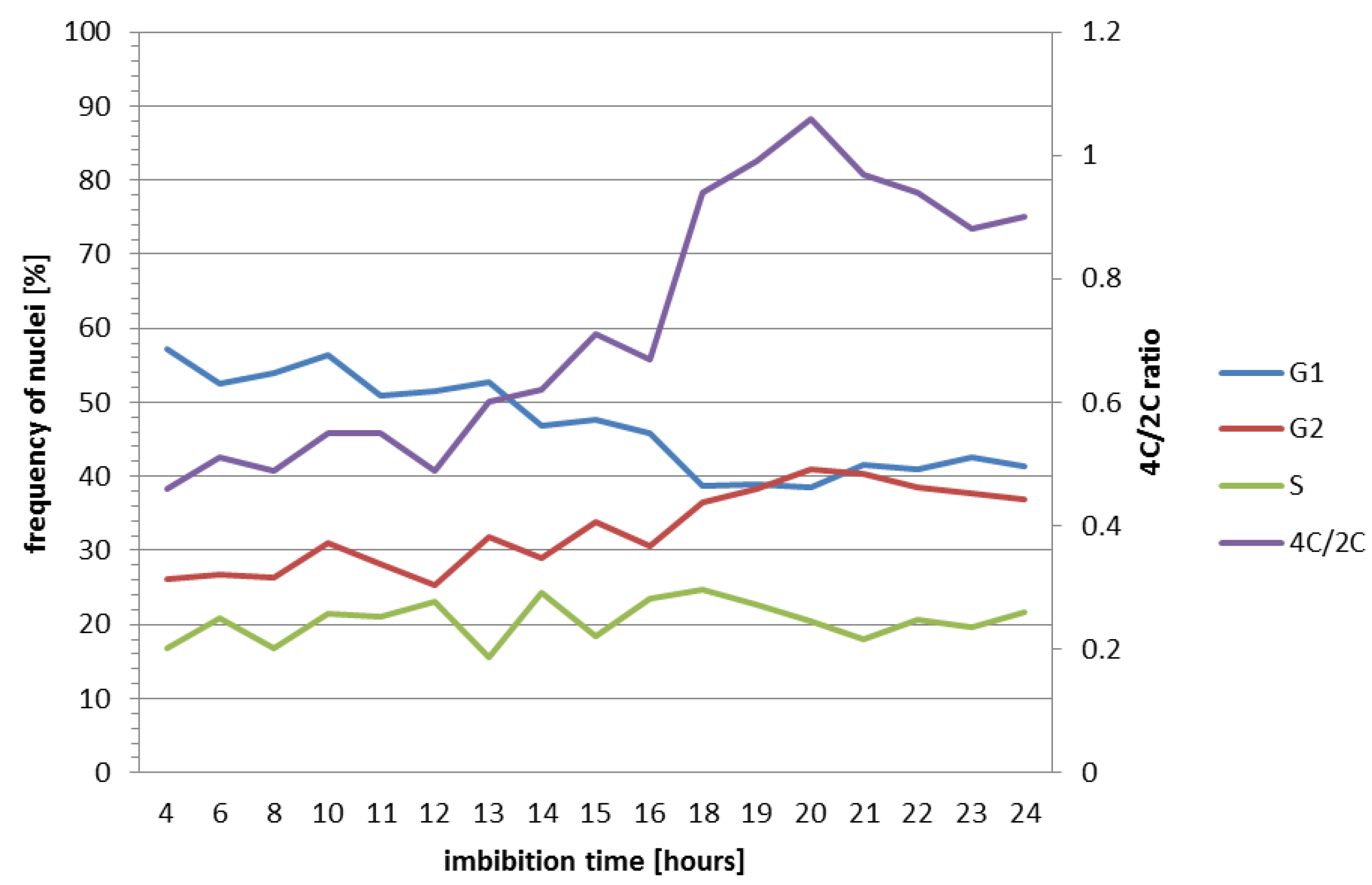
2.2. Nuclear DNA Content and Cell Cycle Analysis in Brachypodium Embryos
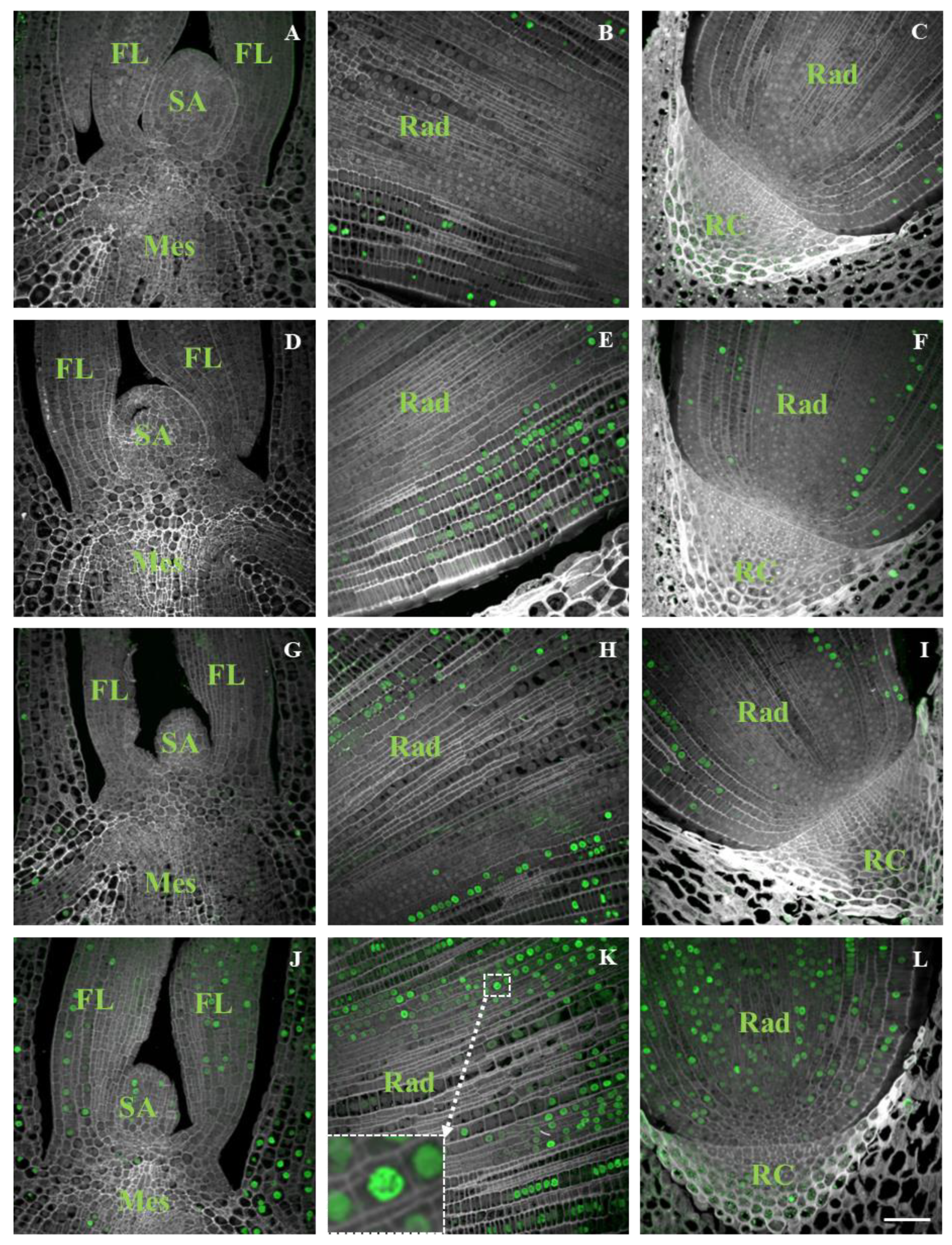
2.3. Distribution of DNA Synthesis
2.4. Analysis of Cell Cycle Gene Transcript Profiles Using Reverse-Transcription PCR
3. Discussion
4. Materials and Methods
4.1. Plant Material and Germination Assay
4.2. Flow Cytometry
4.3. EdU Detection on Embryo Cross Sections
4.4. Gene Expression Analysis
5. Conclusions
- Brachypodium embryos exhibit polysomaty, and nuclei with 2C, 4C, 8C, and 16C have been detected among embryo tissues. Nuclei with higher than 4C DNA content were found in the scutellum, coleorhiza, and coleoptile cells.
- The cell cycle was initiated before radicle protrusion through coleorhiza and radicle elongation. Brachypodium embryo cells initiated DNA replication after only a few hours of imbibition and the first EdU-labelled nuclei were visible after 11 h of imbibition in the radicle tissues.
Author Contributions
Funding
Conflicts of Interest
References
- Rajjou, L.; Duval, M.; Gallardo, K.; Catusse, J.; Bally, J.; Job, C.; Job, D. Seed germination and vigor. Annu. Rev. Plant. Biol. 2012, 63, 507–533. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H. Seed Biology Updates—Highlights and New Discoveries in Seed Dormancy and Germination Research. Front. Plant Sci. 2017, 8, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination—Still a mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Bewley, J.D. Seed germination and dormancy. Plant Cell 1997, 9, 1055–1066. [Google Scholar] [CrossRef] [PubMed]
- Ma, Z.; Bykova, N.A.; Igamberdiev, A.U. Cell signaling mechanisms and metabolic regulation of germination and dormancyin barley seeds. Crop. J. 2017, 5, 459–477. [Google Scholar] [CrossRef]
- Weitbrecht, K.; Muller, K.; Leubner-Metzger, G. First off the mark: Early seed germination. J. Exp. Bot. 2011, 62, 3289–3309. [Google Scholar] [CrossRef] [PubMed]
- Barroco, R.M.; Van Poucke, K.; Bergervoet, J.H.; De Veylder, L.; Groot, S.P.; Inze, D.; Engler, G. The role of the cell cycle machinery in resumption of postembryonic development. Plant Physiol. 2005, 137, 127–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gendreau, E.; Cayla, T.; Corbineau, F. S phase of the cell cycle: A key phase for the regulation of thermodormancy in barley grain. J. Exp. Bot. 2012, 63, 5535–5543. [Google Scholar] [CrossRef] [PubMed]
- De Castro, R.D.; Van Lammeren, A.A.; Groot, S.P.; Bino, R.J.; Hilhorst, H.W. Cell division and subsequent radicle protrusion in tomato seeds are inhibited by osmotic stress but DNA synthesis and formation of microtubular cytoskeleton are not. Plant Physiol. 2000, 122, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Inze, D.; De Veylder, L. Cell cycle regulation in plant development. Annu. Rev. Genet. 2006, 40, 77–105. [Google Scholar] [CrossRef] [PubMed]
- Menges, M.; de Jager, S.M.; Gruissem, W.; Murray, J.A. Global analysis of the core cell cycle regulators of Arabidopsis identifies novel genes, reveals multiple and highly specific profiles of expression and provides a coherent model for plant cell cycle control. Plant J. 2005, 41, 546–566. [Google Scholar] [CrossRef] [PubMed]
- Nieuwland, J.; Stamm, P.; Wen, B.; Randall, R.S.; Murray, J.A.; Bassel, G.W. Re-induction of the cell cycle in the Arabidopsis post-embryonic root meristem is ABA-insensitive, GA-dependent and repressed by KRP6. Sci. Rep. 2016, 6, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Chen, F.; Li, X.; Cao, H.; Ding, M.; Zhang, C.; Zuo, J.; Xu, C.; Xu, J.; Deng, X.; et al. Arabidopsis seed germination speed is controlled by SNL histone deacetylase-binding factor-mediated regulation of AUX1. Nat. Commun. 2016, 7, 13412. [Google Scholar] [CrossRef] [PubMed]
- Oracz, K.; Karpinski, S. Phytohormones Signaling Pathways and ROS Involvement in Seed Germination. Front. Plant Sci. 2016, 7, 864. [Google Scholar] [CrossRef] [PubMed]
- Draper, J.; Mur, L.A.; Jenkins, G.; Ghosh-Biswas, G.C.; Bablak, P.; Hasterok, R.; Routledge, A.P. Brachypodium distachyon. A new model system for functional genomics in grasses. Plant Physiol. 2001, 127, 1539–1555. [Google Scholar] [CrossRef] [PubMed]
- Girin, T.; David, L.C.; Chardin, C.; Sibout, R.; Krapp, A.; Ferrario-Mery, S.; Daniel-Vedele, F. Brachypodium: A promising hub between model species and cereals. J. Exp. Bot. 2014, 65, 5683–5696. [Google Scholar] [CrossRef] [PubMed]
- Catalan, P.; Chalhoub, B.; Chochois, V.; Garvin, D.F.; Hasterok, R.; Manzaneda, A.J.; Mur, L.A.J.; Pecchioni, N.; Rasmussen, S.K.; Vogel, J.P.; et al. Update of the genomics and basic biology of Brachypodium. International Brachypodium Initiative (IBI). Trends Plant Sci. 2014, 19, 414–418. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Salazar, A.; Zabotina, O.A.; Hong, M. Structure and dynamics of Brachypodium primary cell wall polysaccharides from two-dimensional (13)C solid-state nuclear magnetic resonance spectroscopy. Biochemistry 2014, 53, 2840–2854. [Google Scholar] [CrossRef] [PubMed]
- Betekhtin, A.; Milewska-Hendel, A.; Lusinska, J.; Chajec, L.; Kurczynska, E.; Hasterok, R. Organ and tissue-specific localisation of selected cell wall epitopes in the zygotic embryo of Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 725. [Google Scholar] [CrossRef] [PubMed]
- Christensen, U.; Alonso-Simon, A.; Scheller, H.V.; Willats, W.G.; Harholt, J. Characterization of the primary cell walls of seedlings of Brachypodium distachyon—A potential model plant for temperate grasses. Phytochemistry 2010, 71, 62–69. [Google Scholar] [CrossRef] [PubMed]
- Routledge, A.P.; Shelley, G.; Smith, J.V.; Talbot, N.J.; Draper, J.; Mur, L.A. Magnaporthe grisea interactions with the model grass Brachypodium distachyon closely resemble those with rice (Oryza sativa). Mol. Plant Pathol. 2004, 5, 253–265. [Google Scholar] [CrossRef] [PubMed]
- Fitzgerald, T.L.; Powell, J.J.; Schneebeli, K.; Hsia, M.M.; Gardiner, D.M.; Bragg, J.N.; McIntyre, C.L.; Manners, J.M.; Ayliffe, M.; Watt, M.; et al. Brachypodium as an emerging model for cereal-pathogen interactions. Ann. Bot. 2015, 115, 717–731. [Google Scholar] [CrossRef] [PubMed]
- Gendreau, E.; Romaniello, S.; Barad, S.; Leymarie, J.; Benech-Arnold, R.; Corbineau, F. Regulation of cell cycle activity in the embryo of barley seeds during germination as related to grain hydration. J. Exp. Bot. 2008, 59, 203–212. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baiza, A.M.; Vazquez-Ramos, J.M.; Sanchez de Jimenez, E. DNA synthesis and cell division in embryonic maize tissues during germination. J Plant Physiol. 1989, 135, 416–421. [Google Scholar] [CrossRef]
- Georgieva, E.I.; Lopez-Rodas, G.; Hittmair, A.; Feichtinger, H.; Brosch, G.; Loidl, P. Maize embryo germination. I. Cell cycle analysis. Planta 1994, 192, 118–124. [Google Scholar] [CrossRef]
- Resentini, F.; Felipo-Benavent, A.; Colombo, L.; Blazquez, M.A.; Alabadi, D.; Masiero, S. TCP14 and TCP15 mediate the promotion of seed germination by gibberellins in Arabidopsis thaliana. Mol. Plant 2015, 8, 482–485. [Google Scholar] [CrossRef] [PubMed]
- Guillon, F.; Larre, C.; Petipas, F.; Berger, A.; Moussawi, J.; Rogniaux, H.; Santoni, A.; Saulnier, L.; Jamme, F.; Miquel, M.; et al. A comprehensive overview of grain development in Brachypodium distachyon variety Bd21. J. Exp. Bot. 2011, 63, 739–755. [Google Scholar] [CrossRef] [PubMed]
- Opanowicz, M.; Hands, P.; Betts, D.; Parker, M.L.; Toole, G.A.; Mills, E.N.; Doonan, J.H.; Drea, S. Endosperm development in Brachypodium distachyon. J. Exp. Bot. 2011, 62, 735–748. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Jacobsen, J.V.; Talbot, M.J.; White, R.G.; Swain, S.M.; Garvin, D.F.; Gubler, F. Grain dormancy and light quality effects on germination in the model grass Brachypodium distachyon. New Phytol. 2011, 193, 376–386. [Google Scholar] [CrossRef] [PubMed]
- Wolny, E.; Braszewska-Zalewska, A.; Hasterok, R. Spatial distribution of epigenetic modifications in Brachypodium distachyon embryos during seed maturation and germination. PLoS ONE 2014, 9, e101246. [Google Scholar] [CrossRef] [PubMed]
- Wolny, E.; Braszewska-Zalewska, A.; Kroczek, D.; Hasterok, R. Histone H3 and H4 acetylation patterns are more dynamic than those of DNA methylation in Brachypodium distachyon embryos during seed maturation and germination. Protoplasma 2017, 254, 2045–2052. [Google Scholar] [CrossRef] [PubMed]
- Kosina, R.; Tomaszewska, P. Variability of breeding system, caryopsis microstructure and germination in annual and perennial species of the genus Brachypodium P. Beauv. Genet. Resour. Crop. Evol. 2016, 63, 1003–1021. [Google Scholar] [CrossRef]
- Bino, R.J.; Lanteri, S.; Verhoeven, H.A.; KraaK, H.L. Flow cytometric determination of nuclear replication stages in seed tissues. Ann. Bot. 1993, 72, 181–187. [Google Scholar] [CrossRef]
- Innocenti, A.M.; Bitonti, M.B. Changes in histones/DNA ratio in scutellum nuclei during ageing of Triticum durum caryopses. Caryol. Int. J. Cytol. Cytosyst. Cytogenet. 1981, 34, 179–186. [Google Scholar]
- Ahmed, Z.; Yang, H.; Fu, Y.B. The associative changes in scutellum nuclear content and morphology with viability loss of naturally aged and accelerated aging wheat (Triticum aestivum) seeds. Front. Plant Sci. 2016, 7, 1474. [Google Scholar] [CrossRef] [PubMed]
- Dominguez, F.; Cejudo, F.J. Programmed cell death (PCD): An essential process of cereal seed development and germination. Front. Plant Sci. 2014, 5, 366. [Google Scholar] [CrossRef] [PubMed]
- Barrero, J.M.; Talbot, M.J.; White, R.G.; Jacobsen, J.V.; Gubler, F. Anatomical and transcriptomic studies of the coleorhiza reveal the importance of this tissue in regulating dormancy in barley. Plant Physiol. 2009, 150, 1006–1021. [Google Scholar] [CrossRef] [PubMed]
- Jing, H.-C.; Van Lammeren, A.A.; De Castro, R.D.; Bino, R.J.; Hilhorst, H.W.; Groot, S.P. B-tubulin accumulation and DNA synthesis are sequentially resumed in embryo organs of cucumber (Cucumis sativus L.) seeds during germination. Protoplasma 1999, 208, 230–239. [Google Scholar] [CrossRef]
- Gornik, K.; De Castro, R.D.; Liu, Y.; Bino, R.J.; Groot, S.P. Inhibition of cell division during cabbage (Bras sica oleracea L.) seed germination. Seed Sci. Res. 1997, 7, 333–340. [Google Scholar] [CrossRef]
- Kono, A.; Umeda-Hara, C.; Lee, J.; Ito, M.; Uchimiya, H.; Umeda, M. Arabidopsis D-type cyclin CYCD4;1 is a novel cyclin partner of B2-type cyclin-dependent kinase. Plant Physiol. 2003, 132, 1315–1321. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, H.; Ohno, R.; Umeda, M. The Arabidopsis cyclin-dependent kinase-activating kinase CDKF;1 is a major regulator of cell proliferation and cell expansion but is dispensable for CDKA activation. Plant J. 2009, 59, 475–487. [Google Scholar] [CrossRef] [PubMed]
- Takatsuka, H.; Umeda-Hara, C.; Umeda, M. Cyclin-dependent kinase-activating kinases CDKD;1 and CDKD;3 are essential for preserving mitotic activity in Arabidopsis thaliana. Plant J. 2015, 82, 1004–1017. [Google Scholar] [CrossRef] [PubMed]
- Masubelele, N.H.; Dewitte, W.; Menges, M.; Maughan, S.; Collins, C.; Huntley, R.; Nieuwland, J.; Scofield, S.; Murray, J.A. D-type cyclins activate division in the root apex to promote seed germination in Arabidopsis. Proc. Natl. Acad. Sci. USA 2005, 102, 15694–15699. [Google Scholar] [CrossRef] [PubMed]
- Schnittger, A.; De Veylder, L. The Dual Face of Cyclin B1. Trends Plant Sci. 2018, 23, 475–478. [Google Scholar] [CrossRef] [PubMed]
- Steedman, H.F. Polyester wax; a new ribboning embedding medium for histology. Nature 1957, 179, 1345. [Google Scholar] [CrossRef] [PubMed]
- Vitha, S.; Baluska, F.; Jasik, J.; Volkmann, D.; Barlow, P.W. Actin: A Dynamic Framework for Multiple Plant Cell Functions; Springer: Dordrecht, The Netherlands, 2000; pp. 619–636. [Google Scholar]
- Betekhtin, A.; Milewska-Hendel, A.; Chajec, L.; Rojek, M.; Nowak, K.; Kwasniewska, J.; Wolny, E.; Kurczynska, E.; Hasterok, R. 5-Azacitidine induces cell death in a tissue culture of Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 1806. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Imbibition Time (hours) | Nuclei (%) | 4C/2C | ||
|---|---|---|---|---|
| G1 (2C) | S | G2 (4C) | ||
| 4 | 57.1 | 16.7 | 26.11 | 0.46 |
| 6 | 52.57 | 20.76 | 26.67 | 0.51 |
| 8 | 53.9 | 16.8 | 26.29 | 0.49 |
| 10 | 56.46 | 21.51 | 31.03 | 0.55 |
| 11 | 50.92 | 21.01 | 28.07 | 0.55 |
| 12 | 51.48 | 23.18 | 25.34 | 0.49 |
| 13 | 52.72 | 15.54 | 31.74 | 0.60 |
| 14 | 46.77 | 24.24 | 28.98 | 0.62 |
| 15 | 47.74 | 18.39 | 33.87 | 0.71 |
| 16 | 45.91 | 23.42 | 30.67 | 0.67 |
| 17 | n/a | n/a | n/a | n/a |
| 18 | 38.72 | 24.77 | 36.51 | 0.94 |
| 19 | 38.92 | 22.72 | 38.36 | 0.99 |
| 20 | 38.57 | 20.4 | 41.03 | 1.06 |
| 21 | 41.64 | 17.99 | 40.37 | 0.97 |
| 22 | 40.85 | 20.73 | 38.42 | 0.94 |
| 23 | 42.66 | 19.6 | 37.74 | 0.88 |
| 24 | 41.28 | 21.73 | 36.99 | 0.90 |
| Gene Name | Gene Description | Primer Sequence (5′-3′) |
|---|---|---|
| AK437296 | ubiquitin | pF-TCAAAATGCAAGAACGCAAA |
| pR-TCCACACTCCACTTGGTGCT | ||
| Bradi1g54570.1 | protein kinase binding | pF-TTGTGAAGAGGTTCGCGGATGC |
| pR-CCTTCAAGCTCCTTCAGATCC | ||
| Bradi3g02270.1 | cyclin-dependent kinase (CDK), subfamily CDKA | pF-CGAGAAGGTGGAGAAGATCG |
| pR-CGATGGTCTCGTTGGTGTAG | ||
| Bradi4g25980.1 | cyclin-dependent kinase (CDK), subfamily CDKB1 | pF-AAGTGTACAAGGCGCAGGAC |
| pR-ATCCCTTCGTCGTCCATCTC | ||
| Bradi3g40200.1 | cyclin-dependent kinase (CDK), subfamily CDKB2 | pF-AGGGCCAGACCATCCTCTAC |
| pR-GGATCTTCTCGTGGTTCTGG | ||
| Bradi2g26510.1 | cyclin-dependent kinase (CDK), subfamily CDKD | pF-ACAATGGCCAGACATGGTTT |
| pR-CCATTGGAAACAATGAACGA | ||
| Bradi1g14820.1 | CYCLIN, subfamily CYCA3 | pF-ATCCTTGTTGACTGGCTCGT |
| pR-CGGTCGATGTAGGAGATGGT | ||
| Bradi2g52760.1 | CYCLIN, subfamily CYCB1 | pF-GTCCTGGGAAAGCAGAAGGT |
| pR-GGACGTTGACGACGTTGC | ||
| Bradi3g58300.1 | CYCLIN, subfamily CYCD3 | pF-AGCTGTGACTGCTTGCTCAT |
| pR-GATAAGGTCAGACGAGCGGG | ||
| Bradi4g32556.1 | CYCLIN-D4-1-RELATED | pF-CTTGTCTGTAGCGGCCAAGA |
| pR-CTGGATCGTCATGGCTTCGA | ||
| Bradi3g03112.3 | wee1-like protein kinase (WEE1) | pF-AGGATTTCTTCTGCACCCCG |
| pR-GGAGATTTGGGGCAAGGGAT |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wolny, E.; Betekhtin, A.; Rojek, M.; Braszewska-Zalewska, A.; Lusinska, J.; Hasterok, R. Germination and the Early Stages of Seedling Development in Brachypodium distachyon. Int. J. Mol. Sci. 2018, 19, 2916. https://doi.org/10.3390/ijms19102916
Wolny E, Betekhtin A, Rojek M, Braszewska-Zalewska A, Lusinska J, Hasterok R. Germination and the Early Stages of Seedling Development in Brachypodium distachyon. International Journal of Molecular Sciences. 2018; 19(10):2916. https://doi.org/10.3390/ijms19102916
Chicago/Turabian StyleWolny, Elzbieta, Alexander Betekhtin, Magdalena Rojek, Agnieszka Braszewska-Zalewska, Joanna Lusinska, and Robert Hasterok. 2018. "Germination and the Early Stages of Seedling Development in Brachypodium distachyon" International Journal of Molecular Sciences 19, no. 10: 2916. https://doi.org/10.3390/ijms19102916