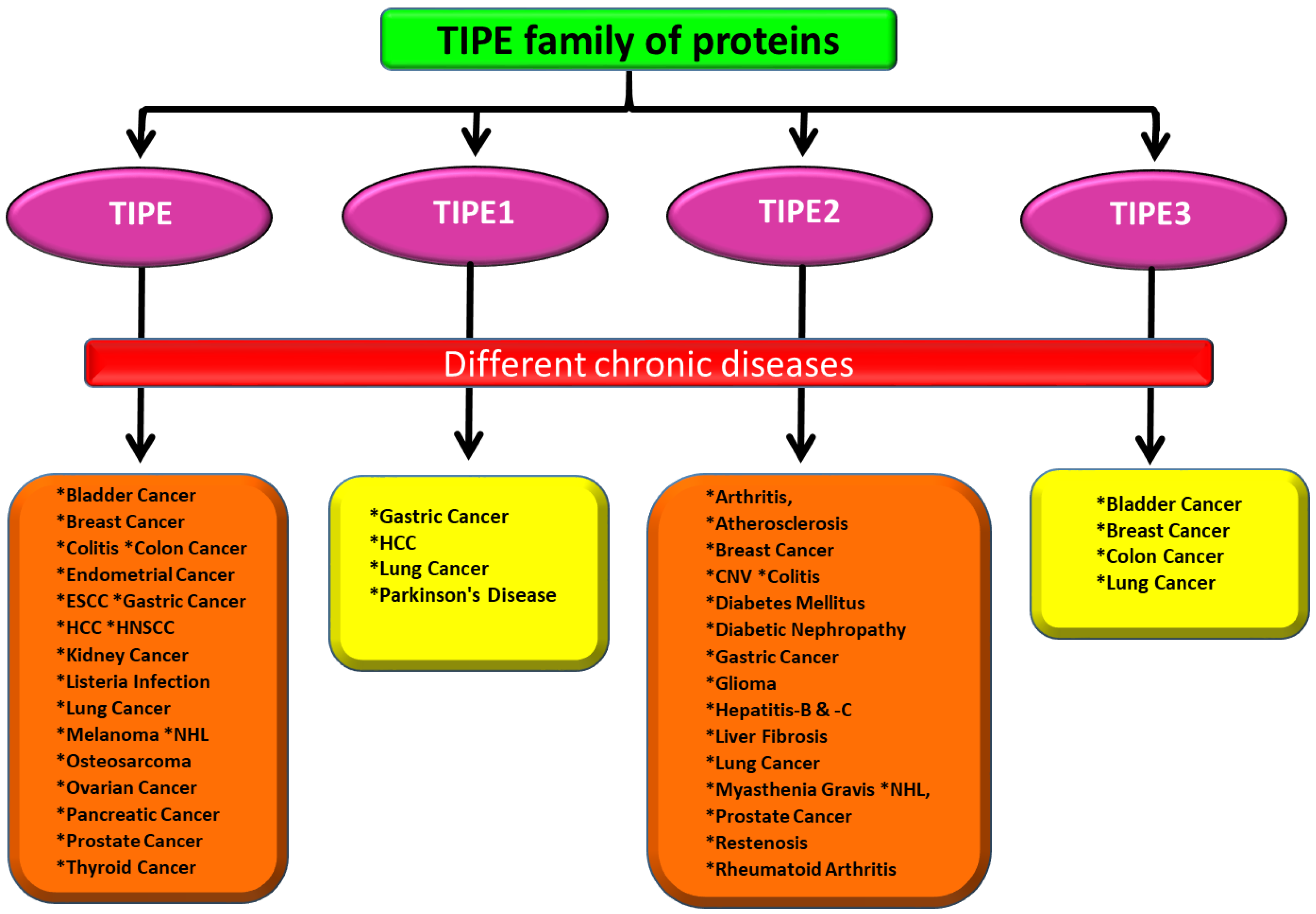
TIPE Family of Proteins and Its Implications in Different Chronic Diseases




 ,
, 
Abstract
:1. Introduction
2. Role of TIPE Family of Proteins in Different Chronic Diseases
2.1. TIPE Family of Proteins and Cancers
2.2. TIPE Family of Proteins and Inflammatory Diseases
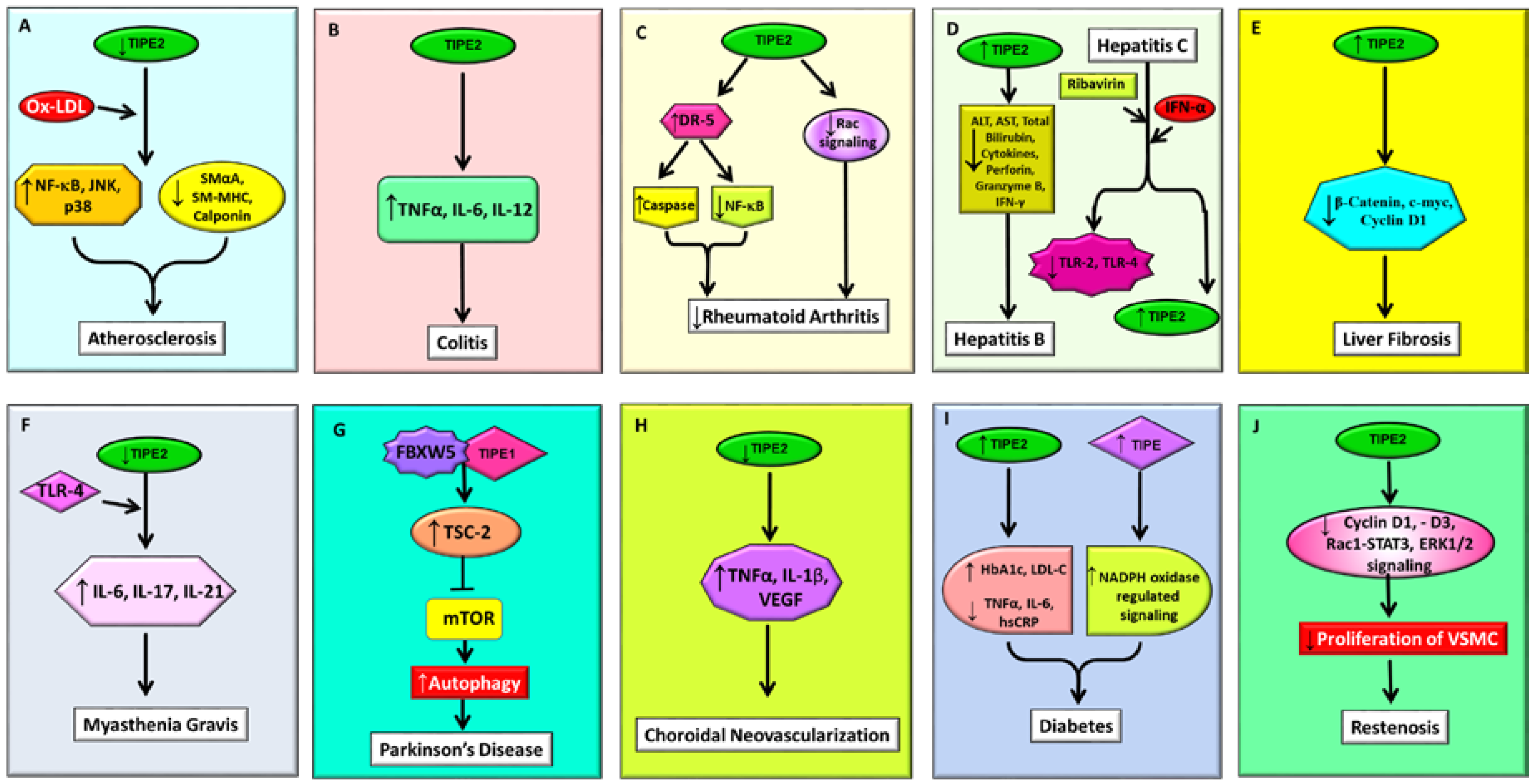
2.2.1. Atherosclerosis
2.2.2. Colitis
2.2.3. Rheumatoid Arthritis
2.3. TIPE Family of Proteins and Infectious Diseases
2.3.1. Hepatitis B
2.3.2. Hepatitis C
2.3.3. Listeria Infection
2.3.4. Liver Fibrosis
2.4. TIPE Family of Proteins in Neuromuscular and Neurodegenerative Diseases
2.4.1. Myasthenia Gravis
2.4.2. Parkinson’s Disease
2.5. The TIPE Family of Proteins and Other Chronic Diseases
2.5.1. Choroidal Neovascularization (CNV)
2.5.2. Diabetes
2.5.3. Restenosis
3. Conclusions
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| TIPE or TNFAIP8 | Tumor necrosis factor-α-induced protein 8 |
| TIPE1 | Tumor necrosis factor-α-induced protein 8-like 1 |
| TIPE2 | Tumor necrosis factor-α-induced protein 8-like 2 |
| TIPE3 | Tumor necrosis factor-α-induced protein 8-like 3 |
| TLR | Toll-like receptor |
| TNF-α | Tumor necrosis factor-α |
| FLIP | Fas-associated death domain-like interleukin-1β-converting enzyme-inhibitory protein |
| mTOR | Mammalian target of rapamycin |
| NF-κB | Nuclear factor κ-light-chain-enhancer of activated B cells |
| MAPK | Mitogen-activated protein kinase |
| MMP | Matrix metalloproteinase |
| VEGF | Vascular endothelial growth factor |
| DLBCL | Diffuse large B-cell lymphoma |
| EMT | Epithelial-to-mesenchymal transition |
| EC | Endometrial carcinoma |
| ESCC | Esophageal squamous cell carcinoma |
| ERK1/2 | Extracellular signal-regulated kinase 1/2 |
| HCC | Hepatocellular carcinoma |
| NHL | Non-Hodgkin’s lymphoma |
| PTCL | Peripheral T-cell lymphoma |
| NSCLC | Non-small cell lung cancer |
| OS | Osteosarcoma |
| CNV | Choroidal neovascularization |
| T2DM | Type 2 diabetes mellitus |
| MG | Myasthenia gravis |
| HBV | Hepatitis B virus |
| HCV | Hepatitis C virus |
| AIH | Autoimmune hepatitis |
| AST | Aspartate aminotransferase |
| ALT | Alanine aminotransferase |
| PBMCs | Peripheral blood mononuclear cells |
| DSS | Dextran sodium sulfate |
| IL-6 | Interleukin-6 |
| Ox-LDL | Oxidized low-density lipoprotein |
| VSMCs | Vascular smooth muscle cells |
| SM-MHC | Smooth muscle-myosin heavy chain |
| SmαA | Smooth muscle α-actin |
| FLSs | Fibroblast-like synoviocytes |
| AA | Adjuvant arthritis |
| RPE | Retinal pigment Epithelium |
References
- Goldsmith, J.R.; Chen, Y.H. Regulation of inflammation and tumorigenesis by the TIPE family of phospholipid transfer proteins. Cell. Mol. Immunol. 2017, 14, 482–487. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shen, P.; Zhang, H.; Su, Z.; Wang, S.; Xu, H. In Silico Analysis of Tumor Necrosis Factor α-Induced Protein 8-Like-1 (TIPE1) Protein. PLoS ONE 2015, 10, e0134114. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.S.; Park, J.; Kim, M.S.; Ha, J.Y.; Jang, Y.W.; Shin, D.H.; Son, J.H. The Tnfaip8-PE complex is a novel upstream effector in the anti-autophagic action of insulin. Sci. Rep. 2017, 7, 6248. [Google Scholar] [CrossRef] [PubMed]
- Sullivan, C.; Lage, C.R.; Yoder, J.A.; Postlethwait, J.H.; Kim, C.H. Evolutionary divergence of the vertebrate TNFAIP8 gene family: Applying the spotted gar orthology bridge to understand ohnolog loss in teleosts. PLoS ONE 2017, 12, e0179517. [Google Scholar] [CrossRef] [PubMed]
- Padmavathi, G.; Banik, K.; Monisha, J.; Bordoloi, D.; Shabnam, B.; Arfuso, F.; Sethi, G.; Fan, L.; Kunnumakkara, A.B. Novel tumor necrosis factor-α induced protein eight (TNFAIP8/TIPE) family: Functions and downstream targets involved in cancer progression. Cancer Lett. 2018, 432, 260–271. [Google Scholar] [CrossRef] [PubMed]
- Sun, Z.; Liu, X.; Song, J.H.; Cheng, Y.; Liu, Y.; Jia, Y.; Meltzer, S.J.; Wang, Z. TNFAIP8 overexpression: A potential predictor of lymphatic metastatic recurrence in pN0 esophageal squamous cell carcinoma after Ivor Lewis esophagectomy. Tumour Biol. 2016, 37, 10923–10934. [Google Scholar] [CrossRef] [PubMed]
- Dong, Q.; Fu, L.; Zhao, Y.; Xie, C.; Li, Q.; Wang, E. TNFAIP8 interacts with LATS1 and promotes aggressiveness through regulation of Hippo pathway in hepatocellular carcinoma. Oncotarget 2017, 8, 15689–15703. [Google Scholar] [CrossRef] [PubMed]
- Xiao, M.; Xu, Q.; Lou, C.; Qin, Y.; Ning, X.; Liu, T.; Zhao, X.; Jia, S.; Huang, Y. Overexpression of TNFAIP8 is associated with tumor aggressiveness and poor prognosis in patients with invasive ductal breast carcinoma. Hum. Pathol. 2017, 62, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Day, T.F.; Mewani, R.R.; Starr, J.; Li, X.; Chakravarty, D.; Ressom, H.; Zou, X.; Eidelman, O.; Pollard, H.B.; Srivastava, M.; et al. Transcriptome and Proteome Analyses of TNFAIP8 Knockdown Cancer Cells Reveal New Insights into Molecular Determinants of Cell Survival and Tumor Progression. Methods Mol. Biol. 2017, 1513, 83–100. [Google Scholar] [PubMed]
- Yu, B.; Xu, L.; Cai, M.; Zhang, D.; Li, S. Effect of tumor necrosis factor-α-induced protein 8 on the immune response of CD4+ T lymphocytes in mice following acute insult. Mol. Med. Rep. 2018, 17, 6655–6660. [Google Scholar] [PubMed]
- Kumar, D.; Whiteside, T.L.; Kasid, U. Identification of a novel tumor necrosis factor-α-inducible gene, SCC-S2, containing the consensus sequence of a death effector domain of fas-associated death domain-like interleukin-1β-converting enzyme-inhibitory protein. J. Biol. Chem. 2000, 275, 2973–2978. [Google Scholar] [CrossRef] [PubMed]
- Xin, Y.; Wan, D.H.; Wang, X.; Gao, X.J.; Xu, X.J.; Ju, X.L.; Li, A.M. Effect of tumor necrosis factor-induced protein 8 on T-cell-mediated immunity in mice after thermal injury. J. Biol. Regulat. Homeostat. Agents 2016, 30, 777–784. [Google Scholar]
- Luan, Y.Y.; Yao, Y.M.; Sheng, Z.Y. The tumor necrosis factor-α-induced protein 8 family in immune homeostasis and inflammatory cancer diseases. J. Biol. Regulat. Homeostat. Agents 2013, 27, 611–619. [Google Scholar]
- Lowe, J.M.; Nguyen, T.A.; Grimm, S.A.; Gabor, K.A.; Peddada, S.D.; Li, L.; Anderson, C.W.; Resnick, M.A.; Menendez, D.; Fessler, M.B. The novel p53 target TNFAIP8 variant 2 is increased in cancer and offsets p53-dependent tumor suppression. Cell Death Differ. 2017, 24, 181–191. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Ma, Y.; Cheng, J.; Li, X.; Zheng, H.; Jiang, L.; Zhou, R. TIPE1 function as a prognosis predictor and negative regulator of lung cancer. Oncotarget 2017, 8, 78496–78506. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Liang, X.; Gao, L.; Ma, H.; Liu, X.; Pan, Y.; Yan, W.; Shan, H.; Wang, Z.; Chen, Y.H.; et al. TIPE1 induces apoptosis by negatively regulating Rac1 activation in hepatocellular carcinoma cells. Oncogene 2015, 34, 2566–2574. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Liu, R.; Luan, Y.Y.; Yao, Y.M. Tumor Necrosis Factor-α Induced Protein 8: Pathophysiology, Clinical Significance, and Regulatory Mechanism. Int. J. Biol. Sci. 2018, 14, 398–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, Y.Y.; Yao, Y.M.; Zhang, L.; Dong, N.; Zhang, Q.H.; Yu, Y.; Sheng, Z.Y. Expression of tumor necrosis factor-α induced protein 8 like-2 contributes to the immunosuppressive property of CD4(+)CD25(+) regulatory T cells in mice. Mol. Immunol. 2011, 49, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Jia, W.; Niu, J.; Zhang, L. Understanding the roles of negative immune regulator TIPE2 in different diseases and tumourigenesis. Histol. Histopathol. 2018, 33, 919–928. [Google Scholar] [PubMed]
- Ruan, Q.; Wang, P.; Wang, T.; Qi, J.; Wei, M.; Wang, S.; Fan, T.; Johnson, D.; Wan, X.; Shi, W.; et al. MicroRNA-21 regulates T-cell apoptosis by directly targeting the tumor suppressor gene Tipe2. Cell Death Dis. 2014, 5, e1095. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Hao, C.; Lou, Y.; Xi, W.; Wang, X.; Wang, Y.; Qu, Z.; Guo, C.; Chen, Y.; Zhang, Y.; et al. Tissue-specific expression of TIPE2 provides insights into its function. Mol. Immunol. 2010, 47, 2435–2442. [Google Scholar] [CrossRef] [PubMed]
- Freundt, E.C.; Bidere, N.; Lenardo, M.J. A different TIPE of immune homeostasis. Cell 2008, 133, 401–402. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Gong, S.; Carmody, R.J.; Hilliard, A.; Li, L.; Sun, J.; Kong, L.; Xu, L.; Hilliard, B.; Hu, S.; et al. TIPE2, a negative regulator of innate and adaptive immunity that maintains immune homeostasis. Cell 2008, 133, 415–426. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Shi, Y.; Wang, Y.; Zhu, F.; Wang, Q.; Ma, C.; Chen, Y.H.; Zhang, L. The unique expression profile of human TIPE2 suggests new functions beyond its role in immune regulation. Mol. Immunol. 2011, 48, 1209–1215. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Zhang, G.; Geng, M.; Zhang, W.; Cui, J.; Liu, S. TIPE2 negatively regulates inflammation by switching arginine metabolism from nitric oxide synthase to arginase. PLoS ONE 2014, 9, e96508. [Google Scholar] [CrossRef] [PubMed]
- Shao, Y.; Chen, H.; Lv, M.; Li, C.; Zhang, W.; Li, Y.; Zhao, X.; Bao, Y. A novel TNFAIP8 gene mediates l-arginine metabolism in Apostichopus japonicus. Fish Shellfish Immunol. 2017, 69, 26–34. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.H.; Yan, H.Q.; Wang, F.; Wang, Y.Y.; Jiang, Y.N.; Wang, Y.N.; Gao, F.G. TIPE2 inhibits TNF-α-induced hepatocellular carcinoma cell metastasis via Erk1/2 downregulation and NF-κB activation. Int. J. Oncol. 2015, 46, 254–264. [Google Scholar] [CrossRef] [PubMed]
- Fayngerts, S.A.; Wang, Z.; Zamani, A.; Sun, H.; Boggs, A.E.; Porturas, T.P.; Xie, W.; Lin, M.; Cathopoulis, T.; Goldsmith, J.R.; et al. Direction of leukocyte polarization and migration by the phosphoinositide-transfer protein TIPE2. Nat. Immunol. 2017, 18, 1353–1360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, T.; Wang, W.; Gong, S.; Sun, H.; Zhang, H.; Yang, A.G.; Chen, Y.H.; Li, X. Genome-wide analysis reveals TNFAIP8L2 as an immune checkpoint regulator of inflammation and metabolism. Mol. Immunol. 2018, 99, 154–162. [Google Scholar] [CrossRef] [PubMed]
- Wang, G.; Guo, C.; Zhao, H.; Pan, Z.; Zhu, F.; Zhang, L.; Wang, Q. TIPE3 differentially modulates proliferation and migration of human non-small-cell lung cancer cells via distinct subcellular location. BMC Cancer 2018, 18, 260. [Google Scholar] [CrossRef] [PubMed]
- Fayngerts, S.A.; Wu, J.; Oxley, C.L.; Liu, X.; Vourekas, A.; Cathopoulis, T.; Wang, Z.; Cui, J.; Liu, S.; Sun, H.; et al. TIPE3 is the transfer protein of lipid second messengers that promote cancer. Cancer Cell 2014, 26, 465–478. [Google Scholar] [CrossRef] [PubMed]
- Lian, K.; Ma, C.; Hao, C.; Li, Y.; Zhang, N.; Chen, Y.H.; Liu, S. TIPE3 protein promotes breast cancer metastasis through activating AKT and NF-κB signaling pathways. Oncotarget 2017, 8, 48889–48904. [Google Scholar] [CrossRef] [PubMed]
- Lee, D.; Kim, M.S.; Park, J.; Jhon, G.J.; Son, J.H.; Shin, D.H. A preliminary X-ray study of murine Tnfaip8/Oxi-α. Int. J. Mol. Sci. 2014, 15, 4523–4530. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Wang, J.; Fan, C.; Li, H.; Sun, H.; Gong, S.; Chen, Y.H.; Shi, Y. Crystal structure of TIPE2 provides insights into immune homeostasis. Nat. Struct. Mol. Biol. 2009, 16, 89–90. [Google Scholar] [CrossRef] [PubMed]
- Antony, P.; Baby, B.; Vijayan, R. Molecular insights into the binding of phosphoinositides to the TH domain region of TIPE proteins. J. Mol. Model. 2016, 22, 272. [Google Scholar] [CrossRef] [PubMed]
- Wang, K.; Ren, Y.; Liu, Y.; Zhang, J.; He, J.J. Tumor Necrosis Factor (TNF)-α-Induced Protein 8-like-2 (TIPE2) Inhibits Proliferation and Tumorigenesis in Breast Cancer Cells. Oncol. Res. 2017, 25, 55–63. [Google Scholar] [CrossRef] [PubMed]
- Garcia, J.A.; Ferreira, H.L.; Vieira, F.V.; Gameiro, R.; Andrade, A.L.; Eugenio, F.R.; Flores, E.F.; Cardoso, T.C. Tumour necrosis factor-α-induced protein 8 (TNFAIP8) expression associated with cell survival and death in cancer cell lines infected with canine distemper virus. Vet. Comp. Oncol. 2017, 15, 336–344. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Chakravarty, D.; Sakabe, I.; Mewani, R.R.; Boudreau, H.E.; Kumar, D.; Ahmad, I.; Kasid, U.N. Role of SCC-S2 in experimental metastasis and modulation of VEGFR-2, MMP-1, and MMP-9 expression. Mol. Ther. 2006, 13, 947–955. [Google Scholar] [CrossRef] [PubMed]
- Kumar, D.; Gokhale, P.; Broustas, C.; Chakravarty, D.; Ahmad, I.; Kasid, U. Expression of SCC-S2, an antiapoptotic molecule, correlates with enhanced proliferation and tumorigenicity of MDA-MB 435 cells. Oncogene 2004, 23, 612–616. [Google Scholar] [CrossRef] [PubMed]
- Shi, T.Y.; Cheng, X.; Yu, K.D.; Sun, M.H.; Shao, Z.M.; Wang, M.Y.; Zhu, M.L.; He, J.; Li, Q.X.; Chen, X.J.; et al. Functional variants in TNFAIP8 associated with cervical cancer susceptibility and clinical outcomes. Carcinogenesis 2013, 34, 770–778. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, Z.; Zhao, T.; Wang, Z.; Xu, Y.; Song, Y.; Wu, J.; Xu, H. SCC-S2 is overexpressed in colon cancers and regulates cell proliferation. Tumour Biol. 2012, 33, 2099–2106. [Google Scholar] [CrossRef] [PubMed]
- Monteith, J.A.; Mellert, H.; Sammons, M.A.; Kuswanto, L.A.; Sykes, S.M.; Resnick-Silverman, L.; Manfredi, J.J.; Berger, S.L.; McMahon, S.B. A rare DNA contact mutation in cancer confers p53 gain-of-function and tumor cell survival via TNFAIP8 induction. Mol. Oncol. 2016, 10, 1207–1220. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, T.; Gao, H.; Yang, M.; Zhao, T.; Liu, Y.; Lou, G. Correlation of TNFAIP8 overexpression with the proliferation, metastasis, and disease-free survival in endometrial cancer. Tumour Biol. 2014, 35, 5805–5814. [Google Scholar] [CrossRef] [PubMed]
- Hadisaputri, Y.E.; Miyazaki, T.; Suzuki, S.; Yokobori, T.; Kobayashi, T.; Tanaka, N.; Inose, T.; Sohda, M.; Kuwano, H. TNFAIP8 overexpression: Clinical relevance to esophageal squamous cell carcinoma. Ann. Surg. Oncol. 2012, 19 (Suppl. 3), S589–S596. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Tao, M.; Wu, J.; Meng, Y.; Xu, C.; Tian, Y.; Zhou, X.; Xiang, J.; Zhang, H.; Xie, Y. Adenovirus-directed expression of TIPE2 suppresses gastric cancer growth via induction of apoptosis and inhibition of AKT and ERK1/2 signaling. Cancer Gene Ther. 2016, 23, 98–106. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Zhang, H.; Xu, C.; Xu, H.; Zhou, X.; Xie, Y.; Tao, M. TIPE2 functions as a metastasis suppressor via negatively regulating β-catenin through activating GSK3β in gastric cancer. Int. J. Oncol. 2016, 48, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Qiu, X.; Hong, S.; Meng, L.; Hong, X.; Qiu, J.; Yang, J.; Zhuang, G.; Liu, Z. Clinical significance of TIPE expression in gastric carcinoma. OncoTargets Ther. 2016, 9, 4473–4481. [Google Scholar]
- Chen, L.; Yang, X.; Yang, X.; Fan, K.; Xiao, P.; Zhang, J.; Wang, X. Association between the expression levels of tumor necrosis factor-α-induced protein 8 and the prognosis of patients with gastric adenocarcinoma. Exp. Ther. Med. 2016, 12, 238–244. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, W.; Chen, Y.; Xie, H.; Guo, Y.; Ren, D.; Li, Y.; Jing, X.; Li, D.; Wang, X.; Zhao, M.; et al. TIPE1 suppresses invasion and migration through down-regulating Wnt/β-catenin pathway in gastric cancer. J. Cell. Mol. Med. 2018, 22, 1103–1117. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Jing, C.; Chen, Y.; Wang, J.; Zhou, M.; Liu, X.; Sun, D.; Mu, L.; Li, L.; Guo, X. Expression of tumor necrosis factor α-induced protein 8 is upregulated in human gastric cancer and regulates cell proliferation, invasion and migration. Mol. Med. Rep. 2015, 12, 2636–2642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Zhao, Q.; Wang, X.; Liu, T.; Yao, G.; Lou, C.; Zhang, Y. TNFAIP8 overexpression is associated with lymph node metastasis and poor prognosis in intestinal-type gastric adenocarcinoma. Histopathology 2014, 65, 517–526. [Google Scholar] [CrossRef] [PubMed]
- Yin, H.; Huang, X.; Tao, M.; Hu, Q.; Qiu, J.; Chen, W.; Wu, J.; Xie, Y. Adenovirus-mediated TIPE2 overexpression inhibits gastric cancer metastasis via reversal of epithelial-mesenchymal transition. Cancer Gene Ther. 2017, 24, 180–188. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.J.; Liu, H.L.; Zhou, H.C.; Wang, G.C. TIPE2 Inhibits Hypoxia-Induced Wnt/β-Catenin Pathway Activation and EMT in Glioma Cells. Oncol. Res. 2016, 24, 255–261. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Tang, Z.; Zhao, Y.; Li, Q.; Wang, E. TNFAIP8 regulates Hippo pathway through interacting with LATS1 to promote cell proliferation and invasion in lung cancer. Mol. Carcinogenesis 2018, 57, 159–166. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Song, Y.; Men, X. Variance of TNFAIP8 expression between tumor tissues and tumor-infiltrating CD4+ and CD8+ T cells in non-small cell lung cancer. Tumour Biol. 2014, 35, 2319–2325. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Guo, C.; Liu, X.; Zhou, C.; Zhu, F.; Wang, X.; Wang, Q.; Shi, Y.; Wang, J.; Zhao, W.; et al. TIPE2 suppresses angiogenesis and non-small cell lung cancer (NSCLC) invasiveness via inhibiting Rac1 activation and VEGF expression. Oncotarget 2016, 7, 62224–62239. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dong, Q.Z.; Zhao, Y.; Liu, Y.; Wang, Y.; Zhang, P.X.; Jiang, G.Y.; Dong, X.J.; Cui, Q.Z.; Wang, E.H. Overexpression of SCC-S2 correlates with lymph node metastasis and poor prognosis in patients with non-small-cell lung cancer. Cancer Sci. 2010, 101, 1562–1569. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Y.; Liu, Y.; Liu, T.; Meng, Q.; Lu, H.; Liu, W.; Hu, J.; Li, C.; Cao, M.; Yan, S.; et al. TNFAIP8 promotes the proliferation and cisplatin chemoresistance of non-small cell lung cancer through MDM2/p53 pathway. Cell Commun. Signal. 2018, 16, 43. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.; Zhang, N.; Geng, M.; Ren, Q.; Li, Y.; Wang, Y.; Chen, Y.H.; Liu, S. Clinical Significance of TIPE2 Protein Upregulation in Non-Hodgkin’s Lymphoma. J. Histochem. Cytochem. 2016, 64, 556–564. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, Y.; Wang, M.Y.; He, J.; Wang, J.C.; Yang, Y.J.; Jin, L.; Chen, Z.Y.; Ma, X.J.; Sun, M.H.; Xia, K.Q.; et al. Tumor necrosis factor-α induced protein 8 polymorphism and risk of non-Hodgkin’s lymphoma in a Chinese population: A case-control study. PLoS ONE 2012, 7, e37846. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Z.; Li, Z.; Shen, Y.; Chen, T. MicroRNA-138 directly targets TNFAIP8 and acts as a tumor suppressor in osteosarcoma. Exp. Ther. Med. 2017, 14, 3665–3673. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, B.; Ren, C. Tumor-suppressive miR-99a inhibits cell proliferation via targeting of TNFAIP8 in osteosarcoma cells. Am. J. Transl. Res. 2016, 8, 1082–1090. [Google Scholar] [PubMed]
- Liu, T.; Gao, H.; Chen, X.; Lou, G.; Gu, L.; Yang, M.; Xia, B.; Yin, H. TNFAIP8 as a predictor of metastasis and a novel prognostic biomarker in patients with epithelial ovarian cancer. Br. J. Cancer 2013, 109, 1685–1692. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Gao, H.; Liu, G.; Gu, L.; Yang, C.; Zhang, F.; Liu, T. Tumor necrosis factor α-induced protein 8 expression as a predictor of prognosis and resistance in patients with advanced ovarian cancer treated with neoadjuvant chemotherapy. Hum. Pathol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Xia, B.; Lu, Y.; Xu, Y.; Lou, G. TNFAIP8 overexpression is associated with platinum resistance in epithelial ovarian cancers with optimal cytoreduction. Hum. Pathol. 2014, 45, 1251–1257. [Google Scholar] [CrossRef] [PubMed]
- Liu, K.; Qin, C.K.; Wang, Z.Y.; Liu, S.X.; Cui, X.P.; Zhang, D.Y. Expression of tumor necrosis factor-α-induced protein 8 in pancreas tissues and its correlation with epithelial growth factor receptor levels. Asian Pac. J. Cancer Prev. 2012, 13, 847–850. [Google Scholar] [CrossRef] [PubMed]
- Lu, Q.; Liu, Z.; Li, Z.; Chen, J.; Liao, Z.; Wu, W.R.; Li, Y.W. TIPE2 Overexpression Suppresses the Proliferation, Migration, and Invasion in Prostate Cancer Cells by Inhibiting PI3K/Akt Signaling Pathway. Oncol. Res. 2016, 24, 305–313. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.; Kallakury, B.V.; Ross, J.S.; Mewani, R.R.; Sheehan, C.E.; Sakabe, I.; Luta, G.; Kumar, D.; Yadavalli, S.; Starr, J.; et al. The significance of TNFAIP8 in prostate cancer response to radiation and docetaxel and disease recurrence. Int. J. Cancer 2013, 133, 31–42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niture, S.; Ramalinga, M.; Kedir, H.; Patacsil, D.; Niture, S.S.; Li, J.; Mani, H.; Suy, S.; Collins, S.; Kumar, D. TNFAIP8 promotes prostate cancer cell survival by inducing autophagy. Oncotarget 2018, 9, 26884–26899. [Google Scholar] [CrossRef] [PubMed]
- Duan, D.; Zhu, Y.Q.; Guan, L.L.; Wang, J. Upregulation of SCC-S2 in immune cells and tumor tissues of papillary thyroid carcinoma. Tumour Biol. 2014, 35, 4331–4337. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Liu, S.; Zhang, C.; Zhang, G.; Li, J.; Ni, M.; An, G.; Dong, M.; Liu, X.; Zhu, F.; et al. Enhanced atherosclerosis in TIPE2-deficient mice is associated with increased macrophage responses to oxidized low-density lipoprotein. J. Immunol. 2013, 191, 4849–4857. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhang, W.; Lou, Y.; Xi, W.; Cui, J.; Geng, M.; Zhu, F.; Chen, Y.H.; Liu, S. TIPE2 deficiency accelerates neointima formation by downregulating smooth muscle cell differentiation. Cell Cycle 2013, 12, 501–510. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lou, Y.; Sun, H.; Morrissey, S.; Porturas, T.; Liu, S.; Hua, X.; Chen, Y.H. Critical roles of TIPE2 protein in murine experimental colitis. J. Immunol. 2014, 193, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Lou, Y.; Porturas, T.; Morrissey, S.; Luo, G.; Qi, J.; Ruan, Q.; Shi, S.; Chen, Y.H. Exacerbated experimental colitis in TNFAIP8-deficient mice. J. Immunol. 2015, 194, 5736–5742. [Google Scholar] [CrossRef] [PubMed]
- Shi, C.; Zhang, S.; Hong, S.; Pang, J.; Yesibulati, Y.; Yin, P.; Zhuang, G. The pro-apoptotic effects of TIPE2 on AA rat fibroblast-like synoviocytes via regulation of the DR5-caspase-NF-κB pathway in vitro. OncoTargets Ther. 2016, 9, 993–1000. [Google Scholar]
- Shi, C.; Wang, Y.; Zhuang, G.; Qi, Z.; Li, Y.; Yin, P. Tumor necrosis factor-α-induced protein8 like 2 regulates lipopolysaccharideinduced rat rheumatoid arthritis immune responses and is associated with Rac activation and interferon regulatory factor 3 phosphorylation. Mol. Med. Rep. 2017, 16, 6346–6353. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Meng, Z.; Guan, J.; Zhang, Z.; Wang, Y. Expression and roles of TIPE2 in autoimmune hepatitis. Exp. Ther. Med. 2017, 13, 942–946. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, W.; Zhang, J.; Zhao, L.; Shao, J.; Cui, J.; Guo, C.; Zhu, F.; Chen, Y.H.; Liu, S. TIPE2 protein negatively regulates HBV-specific CD8(+) T lymphocyte functions in humans. Mol. Immunol. 2015, 64, 204–209. [Google Scholar] [CrossRef] [PubMed]
- Xi, W.; Hu, Y.; Liu, Y.; Zhang, J.; Wang, L.; Lou, Y.; Qu, Z.; Cui, J.; Zhang, G.; Liang, X.; et al. Roles of TIPE2 in hepatitis B virus-induced hepatic inflammation in humans and mice. Mol. Immunol. 2011, 48, 1203–1208. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.Y.; Fan, Y.C.; Zhao, J.; Gao, S.; Sun, F.K.; Han, J.; Yang, Y.; Wang, K. Elevated expression of tumour necrosis factor-α-induced protein 8 (TNFAIP8)-like 2 mRNA in peripheral blood mononuclear cells is associated with disease progression of acute-on-chronic hepatitis B liver failure. J. Viral Hepat. 2014, 21, 64–73. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.C.; Zhang, Y.Y.; Wang, N.; Sun, Y.Y.; Wang, K. Tumor necrosis factor-α-induced protein 8-like 2 (TIPE2) is associated with immune phases of patients with chronic hepatitis B. Oncotarget 2017, 8, 30781–30792. [Google Scholar] [CrossRef] [PubMed]
- Kong, L.; Liu, K.; Zhang, Y.Z.; Jin, M.; Wu, B.R.; Wang, W.Z.; Li, W.; Nan, Y.M.; Chen, Y.H. Downregulation of TIPE2 mRNA expression in peripheral blood mononuclear cells from patients with chronic hepatitis C. Hepatol. Int. 2013, 7, 844–849. [Google Scholar] [CrossRef] [PubMed]
- Porturas, T.P.; Sun, H.; Buchlis, G.; Lou, Y.; Liang, X.; Cathopoulis, T.; Fayngerts, S.; Johnson, D.S.; Wang, Z.; Chen, Y.H. Crucial roles of TNFAIP8 protein in regulating apoptosis and Listeria infection. J. Immunol. 2015, 194, 5743–5750.h. [Google Scholar] [CrossRef] [PubMed]
- Xu, D.D.; Li, X.F.; Li, Y.H.; Liu, Y.H.; Huang, C.; Meng, X.M.; Li, J. TIPE2 attenuates liver fibrosis by reversing the activated hepatic stellate cells. Biochem. Biophys. Res. Commun. 2018, 498, 199–206. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Shao, Z.; Zhang, X.; Jia, X.; Xia, Y.; Zhang, Y.; Xin, N.; Guo, M.; Chen, J.; Zheng, S.; et al. TIPE2 Play a Negative Role in TLR4-Mediated Autoimmune T Helper 17 Cell Responses in Patients with Myasthenia Gravis. J. Neuroimmune Pharmacol. 2015, 10, 635–644. [Google Scholar] [CrossRef] [PubMed]
- Ha, J.Y.; Kim, J.S.; Kang, Y.H.; Bok, E.; Kim, Y.S.; Son, J.H. Tnfaip8 l1/Oxi-β binds to FBXW5, increasing autophagy through activation of TSC2 in a Parkinson’s disease model. J. Neurochem. 2014, 129, 527–538. [Google Scholar] [CrossRef] [PubMed]
- Suo, L.G.; Cui, Y.Y.; Bai, Y.; Qin, X.J. Anti-inflammatory TIPE2 inhibits angiogenic VEGF in retinal pigment epithelium. Mol. Immunol. 2016, 73, 46–52. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Wang, X.; Zhao, Y.; Zhao, P.; Wang, L.; Zhai, Q.; Zhang, X.; Tian, W.; Xiang, X.; Li, T. Upregulation of Tumor Necrosis Factor-α-Induced Protein 8-Like 2 mRNA Is Negatively Correlated with Serum Concentrations of Tumor Necrosis Factor-α and Interleukin 6 in Type 2 Diabetes Mellitus. J. Diabetes Res. 2017, 2017, 4802319. [Google Scholar] [CrossRef] [PubMed]
- Zhang, S.; Zhang, Y.; Wei, X.; Zhen, J.; Wang, Z.; Li, M.; Miao, W.; Ding, H.; Du, P.; Zhang, W.; et al. Expression and regulation of a novel identified TNFAIP8 family is associated with diabetic nephropathy. Biochim. Biophys. Acta 2010, 1802, 1078–1086. [Google Scholar] [CrossRef] [PubMed]
- Zhang, G.; Zhao, L.; Wang, Y.; Shao, J.; Cui, J.; Lou, Y.; Geng, M.; Zhang, N.; Chen, Y.H.; Liu, S. TIPE2 protein prevents injury-induced restenosis in mice. Biochim. Biophys. Acta 2015, 1852, 1574–1584. [Google Scholar] [CrossRef] [PubMed]
- Bordoloi, D.; Roy, N.K.; Monisha, J.; Padmavathi, G.; Kunnumakkara, A.B. Multi-Targeted Agents in Cancer Cell Chemosensitization: What We Learnt from Curcumin Thus Far. Recent Patents Anti-Cancer Drug Discov. 2016, 11, 67–97. [Google Scholar] [CrossRef]
- Kunnumakkara, A.B.; Bordoloi, D.; Harsha, C.; Banik, K.; Gupta, S.C.; Aggarwal, B.B. Curcumin mediates anticancer effects by modulating multiple cell signaling pathways. Clin. Sci. 2017, 131, 1781–1799. [Google Scholar] [CrossRef] [PubMed]
- Aletaha, D.; Neogi, T.; Silman, A.J.; Funovits, J.; Felson, D.T.; Bingham, C.O., 3rd; Birnbaum, N.S.; Burmester, G.R.; Bykerk, V.P.; Cohen, M.D.; et al. Rheumatoid arthritis classification criteria: An American College of Rheumatology/European League against Rheumatism collaborative initiative. Arthritis Rheumat. 2010, 62, 2569–2581. [Google Scholar] [CrossRef] [PubMed]
- Kouchaki, E.; Daneshvar Kakhaki, R.; Tamtaji, O.R.; Dadgostar, E.; Behnam, M.; Zaribaf, A.; Nikoueinejad, H.; Akbari, H.; Asemi, Z. Correlation of serum levels and gene expression of tumor necrosis factor-α-induced protein-8 like-2 with Parkinson disease severity. Metab. Brain Dis. 2018. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
| Disease | Model | Protein Involved | Targets Associated/Outcome | References |
|---|---|---|---|---|
| Cancers | ||||
| Bladder cancer | T24 cells | ↑TIPE3 | ↑PI3K-Akt, ↑MEK-ERK | [31] |
| Breast cancer | Tissue samples | ↑TIPE | tumor progression | [8] |
| MDA-MB-231 and MCF-7 cells | ↑TIPE2 | ↓β-catenin, ↓cyclin D1, ↓c-Myc | [36] | |
| MDA-MB-231 cells transfected into the dorsal flank of female BALB/c nude mice (4–5 weeks old); two groups (vector group and TIPE2 group) | ↓tumor growth | |||
| MCF-7 cells | ↓TIPE | ↑p21 | [14] | |
| MDA-MB-231 and LM2-4175 cells | ↓TIPE | ↑SNX1, ↑NR4A1, ↑AP2A1, ↓IL5 ↓SRC, ↓MAPT, ↓NEK2, ↓TRAF4, ↓PDCL, ↓GTF2F2, ↓GRAP2, ↓ABL1, ↓AKAP2, ↓GAP43, ↓PIK3CA, ↓EGFR, | [9] | |
| HS578T and MCF-7 cells | TIPE | - | [37] | |
| MDA-MB-435 cells MDA-MB 435 cells transfected via tail vein into female BALB/c mice (6–8 weeks old); eight animals in vector group and 12 animals in TIPE group | ↓TIPE | ↓VEGFR-2, ↓MMP-1, ↓MMP-9 | [38] | |
| MCF-7 and MDA-MB-231 | TIPE3 | ↑MMP2, ↑uPA, ↑AKT, ↑NF-κB | [32] | |
| MDA-MB-231 cells | ↑TIPE | - | [39] | |
| Cervical cancer | Tissue samples | ↑TIPE | platinum resistance | [40] |
| Colon cancer | CACO2 and HCT116 cells | ↑TIPE | ↑cyclin D1, ↑phospho-Rb | [41] |
| HT-29 cells | ↓TIPE3 | ↓PI3K-Akt | [31] | |
| HCT116 cells | ↓TIPE | ↑p21 | [14] | |
| HCT116 cells | TIPE | - | [42] | |
| Endometrial cancer | Tumor specimens | ↑TIPE | ↑Ki-67, ↑MMP-9 | [43] |
| ESCC | TE-1, TE-8, and TE-15 cells | ↑TIPE | - | [44] |
| Eca109 cells | ↓TIPE | ↑apoptosis | [6] | |
| Gastric cancer | AGS and HGC-27 cells | ↑TIPE2 | ↓Akt, ↓ERK1/2 | [45] |
| AGS cells xenografted female BALB/c nude mice (4-week-old); Five mice in each group | ||||
| AGS, HGC-27, and SGC-7901 cells | ↑TIPE2 | ↓Akt, ↑GSK3β | [46] | |
| BGC823 cells | ↓TIPE | caspase-3, -8, -9 | [47] | |
| BGC823 cells injected athymic nude mice; four groups containing six animals in each group | ↓TIPE | ↓tumor growth | ||
| Tissue samples | ↑TIPE | ↑metastasis, ↓prognosis | [48] | |
| AGS, BGC-823, and SGC-7901 cells | TIPE1 | ↓Wnt/β-catenin, ↓MMP-2, ↓MMP-9 | [49] | |
| BGC-823 cells injected through tail vein into male nude mice (5–6 weeks old), four groups (including control group) | ||||
| MKN-28, SGC-7901, and MGC-803 cells | ↑TIPE | - | [50] | |
| Tissue samples | ↑TIPE | - | [51] | |
| AGS and HGC-27 cells | ↑TIPE2 | reversal of EMT | [52] | |
| Glioma | U87, U251, and U373 MG cells | ↑TIPE2 | ↓Wnt/β-catenin, ↓EMT | [53] |
| HCC | Bel7402, SK-Hep-1, HepG2, | ↑TIPE | ↓YAP phosphorylation | [7] |
| SMMC7721 and Huh7 cells | ||||
| Bel7402, SMMC7721, QSG770, HepG2, and HepG2.2.1 cells | TIPE1 | ↓Rac1 | [16] | |
| Subcutaneously transplanted H22 cells into male BALB/c mice (6–8 weeks old); two groups (including control group) containing at least five mice in each cohort | ↓tumor growth and weight | |||
| HepG2 cells | TIPE2 | ↓Erk1/2-NF-κB | [27] | |
| Lung cancer | H292 and A549 cells | ↑TIPE1 | ↓cyclin D1, cyclin B1, ↓caspase3, | [15] |
| ↓caspase 8, ↓MMP-2, ↓MMP-9 | ||||
| A549 cells engrafted into the flank of female BALB/c nude mice; two groups (control group and TIPE1 group) containing five mice in each group | ↑TIPE1 | ↓tumor growth | ||
| H460 and H1299 cells | ↑TIPE | ↓phosphorylated LATS1 | [54] | |
| A549 cells | ↓TIPE | ↑p21 | [14] | |
| H1299 cells | TIPE | - | [42] | |
| Tissue samples | ↑TIPE | - | [55] | |
| H1975 and A549 cells | ↑TIPE2 | ↓Rac1, ↓VEGF | [56] | |
| Tissue samples | ↑TIPE | - | [57] | |
| H1975 and A549 cells | ↑TIPE3 | ↑Akt, ↑ERK | [30] | |
| A549 cells transfected into flanks of male BALB/c nude mice (4–6 weeks old), two groups (mock and C-3 flag TIPE3) containing five mice in each group | ↑tumor growth | |||
| NCI-H727 cells | ↑TIPE3 | - | [31] | |
| Tissue samples | - | |||
| NCI-H460 and A549 cells A549 cells injected into flanks of female BALB/c nude mice (4–6 weeks old); two groups (Control and TIPE group) containing 10 mice in each group | ↓TIPE | ↓MDM2, ↓RAD51 ↓tumor volume | [58] | |
| Melanoma | MDA-MB-435 cells | ↓TIPE | ↑NR4A1, ↑AP2A1, ↓TOP2A, ↓EGFR ↓PDCL, ↓GTF2F2, ↓IL5, ↓GRAP2, ↓AKAP2, ↓GAP43, ↓ABL1 | [9] |
| NHL | Tissue samples | ↑TIPE2 | ↑prognosis | [59] |
| 514 NHL patients and 557 cancer-free controls | TIPE | TIPE polymorphism rs1045241C > T | [60] | |
| Osteosarcoma | 143b, LM7, HOS, SaOS-2, | ↑TIPE | Modulation of miR-138 | [61] |
| U2OS and MG-63 cells | ||||
| KHOS, 143b, LM7, | ↑TIPE | Modulation of miR-99a | [62] | |
| U2OS and MG-63 cells | ||||
| U2OS cells | ↓TIPE | ↑p21 | [14] | |
| Ovarian cancer | Tissue samples | ↑TIPE | ↓survival | [63] |
| OVCAR-3 cells | ↓TIPE | G0/G1 cell cycle arrest, ↑beclin 1, ↑LC II | [64] | |
| Tissue samples | ↑TIPE | - | [65] | |
| Pancreatic cancer | Tissue samples | ↑TIPE | ↑EGFR | [66] |
| Prostate cancer | PC-3 cells | ↓TIPE | ↑IGFBP3, ↑NR4A1, ↑AP2A1, ↓IL5, | [9] |
| ↓MAPT, ↓TOP2A, ↓TRAF4, ↓EGFR, | ||||
| ↓PDCL, ↓GTF2F2, ↓GRAP2, ↓ABL1, | ||||
| ↓GAP43, ↓AKAP2, ↓GRIP1 | ||||
| PC-3 cells | ↑TIPE2 | ↓PI3K/Akt signaling | [67] | |
| PC-3 cells | ↑TIPE | ↑MMPs, ↑VEGFR-2 | [68] | |
| PC-3 cells | TIPE | ↑autophagy | [69] | |
| Renal cancer | RCC-RS cells | ↑TIPE | - | [39] |
| Thyroid cancer | Tissue samples | ↑TIPE | - | [70] |
| Inflammatory diseases | ||||
| Atherosclerosis | Ldlr−/− female mice; two groups (wild type and TIPE2−/−) containing eight mice in each group | ↓TIPE2 | ↑JNK, ↑NF-κB, ↑p38 | [71] |
| VSMCs | ↓TIPE2 | ↓contractile proteins, ↑synthetic capacity for growth factors and cytokines | [72] | |
| Colitis | DSS induced male C57BL/6 mice (8-12 weeks old); two groups (wild type and TIPE2−/−) | ↓TIPE2 | ↓TNF-α, ↓IL-6, ↓IL-12 | [73] |
| Colonic epithelial cells | ↓TIPE | ↑cell death | [74] | |
| DSS-induced mice (8–10 weeks old); two groups (wild type and TIPE2−/−) | ↓TIPE | ↓survival rate, ↑body weight loss, ↑leukocyte infiltration, ↑bacterial invasion, ↑inflammatory cytokine production in the colon | ||
| Rheumatoid arthritis | AA-FLSs | ↑TIPE2 | ↑DR5, ↑caspase, ↓NF-κB | [75] |
| Synovial fibroblasts | ↑TIPE2 | ↓Rac signaling | [76] | |
| Infectious diseases | ||||
| AIH | PBMCs | ↓TIPE2 | ↑ALT, ↑AST | [77] |
| Hepatitis B | PBMCs | ↓TIPE2 | ↑perforin, ↑granzyme B, ↑IFN-γ | [78] |
| PBMCs | ↓TIPE2 | ↑ALT, ↑AST, ↑total bilirubin | [79] | |
| ↑HBV load | ||||
| Male C57BL/6 mice (10–12 weeks old); two groups (wild type and TIPE2−/−) | ↓TIPE2 | ↑hepatic inflammation | ||
| Hepatitis B liver failure | PBMCs | ↑TIPE2 | ↓TNF-α, ↓IL-6 | [80] |
| Hepatitis B | PBMCs | ↑TIPE2 | ↓IL-6, ↓TNF-α, ↓IFN-γ | [81] |
| Hepatitis-C induced hepatic inflammation | PBMCs | ↓TIPE2 | ↑TLR signaling | [82] |
| Listeria infection | HEPA1-6 cells | ↓TIPE | ↑apoptosis, deregulated Rac1-GTP | [83] |
| Liver fibrosis | HSC-T6 cells | ↑TIPE2 | ↓β-Catenin, ↓cmyc, ↓cyclin D1 | [84] |
| Neuromuscular and neurodegenerative diseases | ||||
| Myasthenia Gravis | PBMCs | ↓TIPE2 | ↑IL-6, ↑IL-17, ↑IL-21 | [85] |
| Parkinson’s disease | Dopaminergic neurons | ↑TIPE1 | ↑autophagy, ↓mTOR phosphorylation | [86] |
| Other diseases | ||||
| CNV | RPE cells | ↓TIPE2 | ↑TNF-α, ↑IL-1β, ↑VEGF | [87] |
| Diabetes Mellitus | PBMCs | ↑TIPE2 | ↓TNF-α, ↓IL-6 | [88] |
| Diabetic nephropathy | Mesangial cells | ↑TIPE | modulation of NADPH oxidase-mediated signaling pathway | [89] |
| Male Sprague-Dawley diabetic rats | ↑TIPE | - | ||
| Restenosis | VSMCs | ↓TIPE2 | ↑cyclin D1, ↑cyclin D3 | [90] |
| Male C57BL/6J mice (8–12 weeks old); two groups (wild type and TIPE2−/−) | ↓TIPE2 | ↑severity of disease | ||
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bordoloi, D.; Banik, K.; Shabnam, B.; Padmavathi, G.; Monisha, J.; Arfuso, F.; Dharmarajan, A.; Mao, X.; Lim, L.H.K.; Wang, L.; et al. TIPE Family of Proteins and Its Implications in Different Chronic Diseases. Int. J. Mol. Sci. 2018, 19, 2974. https://doi.org/10.3390/ijms19102974
Bordoloi D, Banik K, Shabnam B, Padmavathi G, Monisha J, Arfuso F, Dharmarajan A, Mao X, Lim LHK, Wang L, et al. TIPE Family of Proteins and Its Implications in Different Chronic Diseases. International Journal of Molecular Sciences. 2018; 19(10):2974. https://doi.org/10.3390/ijms19102974
Chicago/Turabian StyleBordoloi, Devivasha, Kishore Banik, Bano Shabnam, Ganesan Padmavathi, Javadi Monisha, Frank Arfuso, Arunasalam Dharmarajan, Xinliang Mao, Lina H. K. Lim, Lingzhi Wang, and et al. 2018. "TIPE Family of Proteins and Its Implications in Different Chronic Diseases" International Journal of Molecular Sciences 19, no. 10: 2974. https://doi.org/10.3390/ijms19102974
APA StyleBordoloi, D., Banik, K., Shabnam, B., Padmavathi, G., Monisha, J., Arfuso, F., Dharmarajan, A., Mao, X., Lim, L. H. K., Wang, L., Fan, L., Hui, K. M., Kumar, A. P., Sethi, G., & Kunnumakkara, A. B. (2018). TIPE Family of Proteins and Its Implications in Different Chronic Diseases. International Journal of Molecular Sciences, 19(10), 2974. https://doi.org/10.3390/ijms19102974