Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients


 and
and 
Abstract
:
1. Introduction
2. Results
2.1. Seaweeds’ Nutritional Characterization
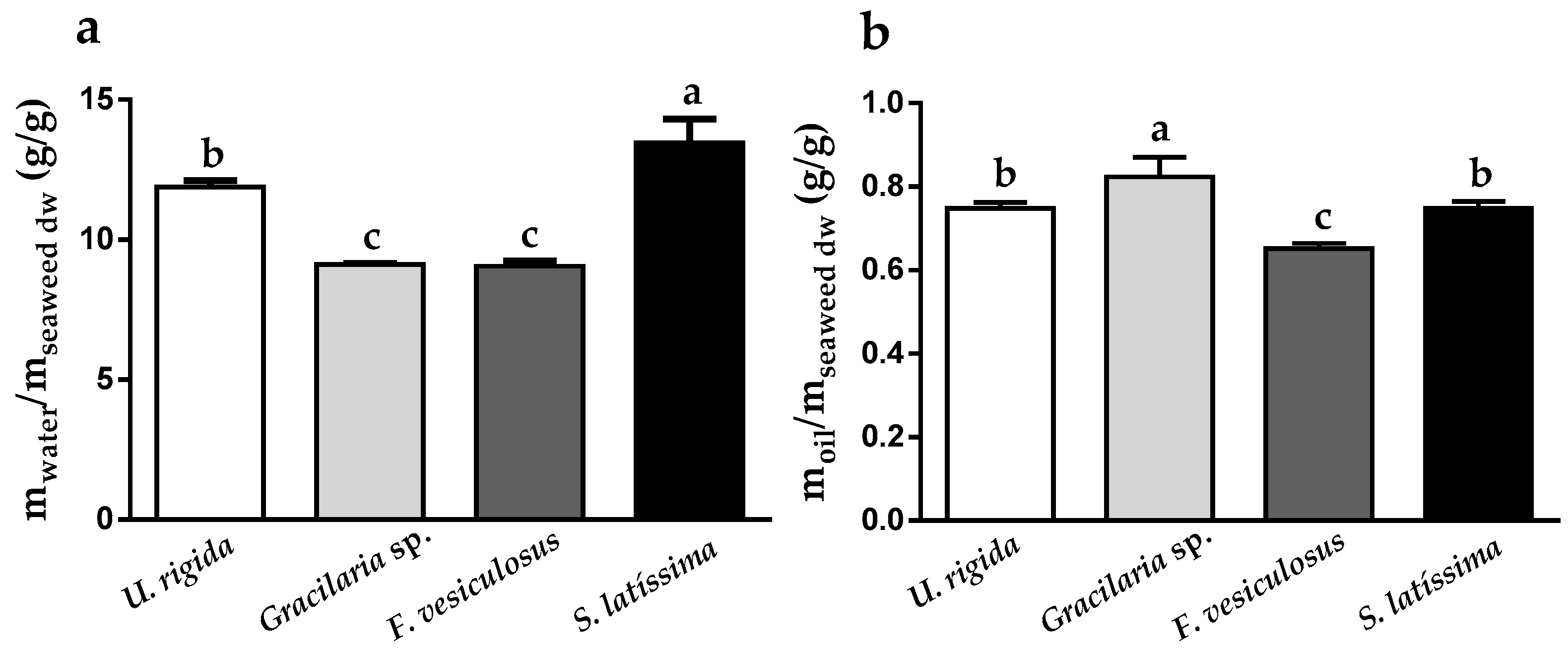
2.2. Seaweeds’ Water and Oil Holding Capacities
2.3. Phenolic Content and Antioxidant Activity of Seaweed Extracts
2.4. Enzyme Activity Inhibition by Seaweed Extracts
2.5. Seaweed Extracts’ Nutritional Characterization
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Seaweeds
4.3. Chemical Characterization
4.3.1. Proximate Composition
4.3.2. Mineral Composition
4.3.3. Lipidic Profile
4.4. Seaweeds’ Water and Oil Holding Capacity
4.5. Extract Preparation
4.6. Quantification of Phenolic Content of the Extracts
4.7. Extracts’ Antioxidant Activity
4.8. Ability of Seaweed Extracts to Inhibit Key Metabolic Syndrome Relevant Enzymes
4.8.1. α-Glucosidase Activity
4.8.2. Pancreatic Lipase Activity
4.8.3. Angiotensin Conversing Enzyme I (ACE) activity
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- World Health Organization: Cardiovascular Disease. Available online: http://www.who.int/cardiovascular_diseases/about_cvd/en/ (accessed on 12 May 2018).
- Aldworth, J.; Bache, N.; Hegelund, M.H.; Hirst, M.; Linnenkamp, U.; Magliano, D.; Oomatia, F.; Patterson, C.; Peer, N.; Pritulskiy, A.; et al. Chapter 3 in IDF Diabetes Atlas, 7th ed.; International Diabetes Federation: Brussels, Belgium, 2015; pp. 50–51. ISBN 978-2-930229-81-2. [Google Scholar]
- World Heart Federation. Available online: http://www.world-heart-federation.org/cardiovascular-health/cardiovascular-disease-risk-factors/ (accessed on 12 May 2018).
- Chen, C.; Pearson, A.M.; Gray, J.I. Effects of synthetic antioxidants (BHA, BHT and PG) on the mutagenicity of IQ-like compounds. Food Chem. 1992, 43, 177–183. [Google Scholar] [CrossRef]
- Brown, E.M.; Allsopp, P.J.; Magee, P.J.; Gill, C.I.; Nitecki, S.; Strain, C.R.; Mcsorley, E.M. Seaweed and human health. Nutr. Rev. 2014, 72, 205–216. [Google Scholar] [CrossRef] [PubMed]
- Le Bras, Q.; Leseur, M.; Lucas, S.; Gouin, S. Etude du marché français des algues alimentaires. Panorama de la distribution. Programme IDEALG Phase 2 in Agrocampus Ouest, 36th ed.; Les Publ. du Pôle Halieut: Rennes cedex, France, 2015; p. 42. [Google Scholar]
- Lordan, S.; Ross, R.P.; Stanton, C. Marine bioactives as functional food ingredients: Potential to reduce the incidence of chronic diseases. Mar. Drugs 2011, 9, 1056–1100. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.-Y.; Huang, X.; Cheong, K.-L. Recent Advances in Marine Algae Polysaccharides: Isolation, Structure, and Activities. Mar. Drugs 2017, 15, 388. [Google Scholar] [CrossRef] [PubMed]
- Jiménez-Escrig, A.; Sánchez-Muniz, F.J. Dietary fibre from edible seaweeds: Chemical structure, physicochemical properties and effects on cholesterol metabolism. Nutr. Res. 2000, 20, 585–598. [Google Scholar] [CrossRef] [Green Version]
- Rupérez, P. Mineral content of edible marine seaweeds. Food Chem. 2002, 79, 23–26. [Google Scholar] [CrossRef] [Green Version]
- Hanjabam, M.D.; Zynudheen, A.A.; Ninan, G.; Panda, S. Seaweed as an Ingredient for Nutritional Improvement of Fish Jerky. J. Food Process. Preserv. 2017, 41, 1–8. [Google Scholar] [CrossRef]
- Wells, M.L.; Potin, P.; Craigie, J.S.; Raven, J.A.; Merchant, S.S.; Helliwell, K.E.; Smith, A.G.; Camire, M.E.; Brawley, S.H. Algae as nutritional and functional food sources: Revisiting our understanding. J. Appl. Phycol. 2017, 29, 949–982. [Google Scholar] [CrossRef] [PubMed]
- Kirke, D.A.; Smyth, T.J.; Rai, D.K.; Kenny, O.; Stengel, D.B. The chemical and antioxidant stability of isolated low molecular weight phlorotannins. Food Chem. 2017, 221, 1104–1112. [Google Scholar] [CrossRef] [PubMed]
- Tenorio-Rodriguez, P.A.; Murillo-Álvarez, J.I.; Campa-Cordova, Á.I.; Angulo, C. Antioxidant screening and phenolic content of ethanol extracts of selected Baja California Peninsula macroalgae. J. Food Sci. Technol. 2017, 54, 422–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.-S.; Kum, J.; Jeon, K.; Park, J.; Choi, H.; Hwang, K.-E.; Jeong, T.-J.; Kim, Y.-B.; Kim, C.-J. Effects of Edible Seaweed on Physicochemical and Sensory Characteristics of Reduced-salt Frankfurters. Korean J. Food Sci. Anim. Resour. 2015, 35, 748–756. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kellogg, J.; Grace, M.H.; Lila, M.A. Phlorotannins from alaskan seaweed inhibit carbolytic enzyme activity. Mar. Drugs 2014, 12, 5277–5294. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Angiotensin I-converting enzyme (ACE) inhibitory activity of Fucus spiralis macroalgae and influence of the extracts storage temperature—A short report. J. Pharm. Biomed. Anal. 2016, 131, 503–507. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Neto, A.I.; Marcone, M.; Baptista, J. Nutritional and Functional Bioactivity Value of Selected Azorean Macroalgae: Ulva compressa, Ulva rigida, Gelidium microdon, and Pterocladiella capillacea. J. Food Sci. 2017, 82, 1757–1764. [Google Scholar] [CrossRef] [PubMed]
- Paiva, L.; Lima, E.; Neto, A.I.; Baptista, J. Isolation and characterization of angiotensin I-converting enzyme (ACE) inhibitory peptides from Ulva rigida C. Agardh protein hydrolysate. J. Funct. Foods 2016, 26, 65–76. [Google Scholar] [CrossRef]
- Lorenzo, J.M.; Agregán, R.; Munekata, P.E.S.; Franco, D.; Carballo, J.; Şahin, S.; Lacomba, R.; Barba, F.J. Proximate composition and nutritional value of three macroalgae: Ascophyllum nodosum, Fucus vesiculosus and Bifurcaria bifurcata. Mar. Drugs 2017, 15, 360. [Google Scholar] [CrossRef] [PubMed]
- Wang, T.; Jónsdóttir, R.; Kristinsson, H.G.; Thorkelsson, G.; Jacobsen, C.; Hamaguchi, P.Y.; Ólafsdóttir, G. Inhibition of haemoglobin-mediated lipid oxidation in washed cod muscle and cod protein isolates by Fucus vesiculosus extract and fractions. Food Chem. 2010, 123, 321–330. [Google Scholar] [CrossRef]
- Kim, K.-T.; Rioux, L.-E.; Turgeon, S.L. Alpha-amylase and alpha-glucosidase inhibition is differentially modulated by fucoidan obtained from Fucus vesiculosus and Ascophyllum nodosum. Phytochemistry 2014, 98, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Vázquez-Freire, M.J.; Lamela, M.; Calleja, J.M. Hypolipidaemic Activity of a Polysaccharide Extract from Fucus vesiculosus L. Phyther. Res. 1996, 10, 647–650. [Google Scholar] [CrossRef]
- Sharma, S.; Neves, L.; Funderud, J.; Mydland, L.T.; Øverland, M.; Horn, S.J. Seasonal and depth variations in the chemical composition of cultivated Saccharina latissima. Algal Res. 2018, 32, 107–112. [Google Scholar] [CrossRef]
- Stévant, P.; Indergård, E.; Ólafsdóttir, A.; Marfaing, H.; Larssen, W.E.; Fleurence, J.; Roleda, M.Y.; Rustad, T.; Slizyte, R.; Nordtvedt, T.S. Effects of drying on the nutrient content and physico-chemical and sensory characteristics of the edible kelp Saccharina latissima. J. Appl. Phycol. 2018, 30, 2587–2599. [Google Scholar] [CrossRef]
- Wen, X.; Peng, C.; Zhou, H.; Lin, Z.; Lin, G.; Chen, S.; Li, P. Nutritional Composition and Assessment of Gracilaria lemaneiformis Bory. J. Integr. Plant Biol. 2006, 48, 1047–1053. [Google Scholar] [CrossRef]
- Chan, P.T.; Matanjun, P. Chemical composition and physicochemical properties of tropical red seaweed, Gracilaria changii. Food Chem. 2017, 221, 302–310. [Google Scholar] [CrossRef] [PubMed]
- Chan, P.T.; Matanjun, P.; Yasir, S.M.; Tan, T.S. Antioxidant and hypolipidaemic properties of red seaweed, Gracilaria changii. J. Appl. Phycol. 2014, 26, 987–997. [Google Scholar] [CrossRef]
- Chan, P.T.; Matanjun, P.; Yasir, S.M.; Tan, T.S. Histopathological studies on liver, kidney and heart of normal and dietary induced hyperlipidaemic rats fed with tropical red seaweed Gracilaria changii. J. Funct. Foods 2015, 17, 202–213. [Google Scholar] [CrossRef]
- Houghton, D.; Wilcox, M.D.; Chater, P.I.; Brownlee, I.A.; Seal, C.J.; Pearson, J.P. Biological activity of alginate and its effect on pancreatic lipase inhibition as a potential treatment for obesity. Food Hydrocoll. 2015, 49, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Wilcox, M.D.; Brownlee, I.A.; Richardson, J.C.; Dettmar, P.W.; Pearson, J.P. The modulation of pancreatic lipase activity by alginates. Food Chem. 2014, 146, 479–484. [Google Scholar] [CrossRef] [PubMed]
- Cardoso, S.M.; Pereira, O.R.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, A.M.S. Seaweeds as Preventive Agents for Cardiovascular Diseases: From Nutrients to Functional Foods. Mar. Drugs 2015, 13, 6838–6865. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- El Maghraby, D.M.; Fakhry, E.M. Lipid content and fatty acid composition of Mediterranean macro-algae as dynamic factors for biodiesel production. Oceanologia 2015, 57, 86–92. [Google Scholar] [CrossRef]
- Fuentes-Alventosa, J.M.; Rodríguez-Gutiérrez, G.; Jaramillo-Carmona, S.; Espejo-Calvo, J.A.; Rodríguez-Arcos, R.; Fernández-Bolaños, J.; Guillén-Bejarano, R.; Jiménez-Araujo, A. Effect of extraction method on chemical composition and functional characteristics of high dietary fibre powders obtained from asparagus by-products. Food Chem. 2009, 113, 665–671. [Google Scholar] [CrossRef]
- Puls, W.; Keup, U.; Krause, H.P.; Thomas, G.; Hoffmeister, F. Glucosidase inhibition. Naturwissenschaften 1977, 64, 536–537. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.H.; Jeon, Y.J. Anti-diabetic effects of brown algae derived phlorotannins, marine polyphenols through diverse mechanisms. Fitoterapia 2013, 86, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Sánchez de Pedro, R.; Karsten, U.; Niell, F.X.; Carmona, R. Intraspecific phenotypic variation in two estuarine rhodophytes across their intertidal zonation. Mar. Biol. 2016, 163, 1–14. [Google Scholar] [CrossRef]
- Frikha, F.; Kammoun, M.; Hammami, N.; Mchirgui, R.A.; Belbahri, L.; Gargouri, Y.; Ben-Rebah, F. Chemical composition and some biological activities of marine algae collected in Tunisia. Ciencias Mar. 2011, 37, 113–124. [Google Scholar] [CrossRef]
- McDermid, K.J.; Stuercke, B. Nutritional composition of edible Hawaiian seaweeds. J. Appl. Phycol. 2003, 15, 513–524. [Google Scholar] [CrossRef]
- Tibbets, S.M.; Milley, J.E.; Lall, S.P. Nutritional quality of some wild and cultivated seaweeds: Nutrient composition, total phenolic content and in vitro digestibility. J. Appl. Phycol. 2016, 28, 3575–3585. [Google Scholar] [CrossRef]
- Yaich, H.; Garna, H.; Bchir, B.; Besbes, S.; Paquot, M.; Richel, A.; Blecker, C.; Attia, H. Chemical composition and functional properties of dietary fibre extracted by Englyst and Prosky methods from the alga Ulva lactuca collected in Tunisia. Algal Res. 2015, 9, 65–73. [Google Scholar] [CrossRef]
- Fernández-Martín, F.; López-López, I.; Cofrades, S.; Colmenero, F.J. Influence of adding Sea Spaghetti seaweed and replacing the animal fat with olive oil or a konjac gel on pork meat batter gelation. Potential protein/alginate association. Meat Sci. 2009, 83, 209–217. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- López-López, I.; Cofrades, S.; Jiménez-Colmenero, F. Low-fat frankfurters enriched with n-3 PUFA and edible seaweed: Effects of olive oil and chilled storage on physicochemical, sensory and microbial characteristics. Meat Sci. 2009, 83, 148–154. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.W.; Choi, J.H.; Choi, Y.S.; Han, D.J.; Kim, H.Y.; Lee, M.A.; Kim, S.Y.; Kim, C.J. Effects of sea tangle (Lamina japonica) Powder on quality characteristics of breakfast sausages. Korean J. Food Sci. Anim. Resour. 2010, 30, 55–61. [Google Scholar] [CrossRef]
- López-López, I.; Cofrades, S.; Yakan, A.; Solas, M.T.; Jiménez-Colmenero, F. Frozen storage characteristics of low-salt and low-fat beef patties as affected by Wakame addition and replacing pork backfat with olive oil-in-water emulsion. Food Res. Int. 2010, 43, 1244–1254. [Google Scholar] [CrossRef] [Green Version]
- Moroney, N.C.; O’Grady, M.N.; Lordan, S.; Stanton, C.; Kerry, J.P. Seaweed polysaccharides (laminarin and fucoidan) as functional ingredients in pork meat: An evaluation of anti-oxidative potential, thermal stability and bioaccessibility. Mar. Drugs 2015, 13, 2447–2464. [Google Scholar] [CrossRef] [PubMed]
- Jónsdóttir, R.; Geirsdóttir, M.; Hamaguchi, P.Y.; Jamnik, P.; Kristinsson, H.G.; Undeland, I. The ability of in vitro antioxidant assays to predict the efficiency of a cod protein hydrolysate and brown seaweed extract to prevent oxidation in marine food model systems. J. Sci. Food Agric. 2016, 96, 2125–2135. [Google Scholar] [CrossRef] [PubMed]
- Ortiz, J.; Vivanco, J.P.; Aubourg, S.P. Lipid and sensory quality of canned Atlantic salmon (Salmo salar): Effect of the use of different seaweed extracts as covering liquids. Eur. J. Lipid Sci. Technol. 2014, 116, 596–605. [Google Scholar] [CrossRef]
- Gabbia, D.; Dall’Acqua, S.; Di Gangi, I.M.; Bogialli, S.; Caputi, V.; Albertoni, L.; Marsilio, I.; Paccagnella, N.; Carrara, M.; Giron, M.C.; et al. The Phytocomplex from Fucus vesiculosus and ascophyllum nodosum controls postprandial plasma glucose levels: An in vitro and in Vivo study in a mouse model of NASH. Mar. Drugs 2017, 15, 41. [Google Scholar] [CrossRef] [PubMed]
- Roy, M.C.; Anguenot, R.; Fillion, C.; Beaulieu, M.; Bérubé, J.; Richard, D. Effect of a commercially-available algal phlorotannins extract on digestive enzymes and carbohydrate absorption in vivo. Food Res. Int. 2011, 44, 3026–3029. [Google Scholar] [CrossRef]
- Ali, L.; Khan, A.; Al-Broumi, M.; Al-Harrasi, R.; Al-Kharusi, L.; Hussain, J.; Al-Harrasi, A. New Enzyme-Inhibitory Triterpenoid from Marine Macro Brown Alga Padina boergesenii Allender & Kraft. Mar. Drugs 2017, 15, 19. [Google Scholar] [CrossRef]
- Ali, L.; Khan, A.; Al-Kharusi, L.; Hussain, J.; Al-Harrasi, A. New α-Glucosidase Inhibitory Triterpenic Acid from Marine Macro Green Alga Codium dwarkense Boergs. Mar. Drugs 2015, 13, 4344–4356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, S.-H.; Kang, S.-M.; Ko, S.-C.; Moon, S.-H.; Jeon, B.-T.; Lee, D.H.; Jeon, Y.-J. Octaphlorethol A: A potent α-glucosidase inhibitor isolated from Ishige foliacea shows an anti-hyperglycemic effect in mice with streptozotocin-induced diabetes. Food Funct. 2014, 5, 2602–2608. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.Y.; Nam, K.A.; Kurihara, H.; Kim, S.M. Potent α-glucosidase inhibitors purified from the red alga Grateloupia elliptica. Phytochemistry 2008, 69, 2820–2825. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yu, Z.; Shuai, L.; Guo, Y.; Duan, D.; Fu, P. The effect of kelp on serum lipids of hyperlipidemia in rats. J. Food Biochem. 2013, 37, 129–135. [Google Scholar] [CrossRef]
- Balasubramaniam, V.; Mustar, S.; Mustafa Khalid, N.; Abd Rashed, A.; Mohd Noh, M.F.; Wilcox, M.D.; Chater, P.I.; Brownlee, I.A.; Pearson, J.P. Inhibitory activities of three Malaysian edible seaweeds on lipase and α-amylase. J. Appl. Phycol. 2013, 25, 1405–1412. [Google Scholar] [CrossRef]
- AOAC: Official Method of Analysis; Association of Official Analytical Chemists: Washington, DC, USA, 1990.
- Lourenço, S.O.; Barbarino, E.; De-Paula, J.C.; Pereira, L.O.d.S.; Marquez, U.M.L. Amino acid composition, protein content and calculation of nitrogen-to-protein conversion factors for 19 tropical seaweeds. Phycol. Res. 2002, 50, 233–241. [Google Scholar] [CrossRef]
- DuBois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Isca, V.M.S.; Seca, A.M.L.; Pinto, D.C.G.A.; Silva, H.; Silva, A.M.S. Lipophilic profile of the edible halophyte Salicornia ramosissima. Food Chem. 2014, 165, 330–336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdul-Hamid, A.; Luan, Y.S. Functional properties of dietary fibre prepared from defatted rice bran. Food Chem. 2000, 68, 15–19. [Google Scholar] [CrossRef]
- Catarino, M.D.; Silva, A.M.S.; Cruz, M.T.; Cardoso, S.M. Antioxidant and anti-inflammatory activities of Geranium robertianum L. decoctions. Food Funct. 2017, 8, 3355–3365. [Google Scholar] [CrossRef] [PubMed]
- Benzie, I.F.; Strain, J.J. The ferric reducing ability of plasma (FRAP) as a measure of “antioxidant power”: The FRAP assay. Anal. Biochem. 1996, 239, 70–76. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, D.; Orwat, J.; Hurburt, T.; Woods, F.; Pitts, J.A.; Shetty, K. Evaluation of phenolic bioactive-linked functionality of blackberry cultivars targeting dietary management of early stages type-2 diabetes using in vitro models. Sci. Hortic. 2016, 212, 193–202. [Google Scholar] [CrossRef]
- Panteghini, M.; Bonora, R.; Pagani, F. Measurement of pancreatic lipase activity in serum by a kinetic colorimetric assay using a new chromogenic substrate. Ann. Clin. Biochem. 2001, 38, 365–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hou, W.-C.; Chen, H.-J.; Lin, Y.-H. Antioxidant peptides with Angiotensin converting enzyme inhibitory activities and applications for Angiotensin converting enzyme purification. J. Agric. Food Chem. 2003, 51, 1706–1709. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
{kind=link}
| Total Content (g/100 g Seaweed dw) | Sample | |||
|---|---|---|---|---|
| U. rigida | Gracilaria sp. | F. vesiculosus | S. latissima | |
| Total Carbohydrates | 58.1 ± 0.7 b | 46.9 ± 0.4 d | 56.4 ± 0.4 c | 68.9 ± 0.3 a |
| Fiber | ||||
| Total | 36.6 ± 1.5 b | 40.6 ± 3.8 a,b | 45.0 ± 0.1 a | 40.9 ± 0.6 a |
| Soluble | 17.7 ± 1.1 b | 22.8 ± 1.3 a | 7.6 ± 0.3 d | 12.8 ± 0.3 c |
| Insoluble | 18.9 ± 0.8 c | 17.9 ± 3.3 c | 37.4 ± 0.3 a | 28.2 ± 0.3 b |
| Ash | 31.7 ± 0.6 a | 28.9 ± 0.2 b | 25.5 ± 0.2 c | 20.4 ± 0.1 d |
| Minerals (mg/100 g dw) | ||||
| Na | 2424.0 ± 233.8 b | 1594.7 ± 296.6 c | 2266.1 ± 35.5 b | 3048.3 ± 129.4 a |
| K | 2466.6 ± 65.2 c | 9243.1 ± 205.8 a | 4083.1 ± 61.0 b | 3869.4 ± 138.7 b |
| Ca | 414.3 ± 33.8 c | 200.4 ± 24.3 d | 1382.0 ± 5.1 a | 919.4 ± 32.5 b |
| Mg | 3758.6 ± 430.3 a | 285.7 ± 59.5 b | 835.5 ± 9.7 a | 611.1 ± 25.0 b |
| Fe | 110.2 ± 22.6 b | 211.0 ± 52.3 a | 8.8 ± 1.1 c | 185.4 ± 8.2 a |
| Mn | 6.84 ± 1.05 c | 15.66 ± 2.85 b | 54.66 ± 0.31 a | 0.56 ± 0.02 d |
| Cu | 3.27 ± 0.26 b | 3.46 ± 0.49 b | 3.05 ± 0.04 b | 3.86 ± 0.18 a |
| Zn | 3.27 ± 0.26 a | 3.45 ± 0.61 a | 3.06 ± 0.06 a | 3.86 ± 0.18 a |
| Ni | 1.07 ± 0.09 c | 1.93 ± 0.43 b | 2.49 ± 0.08 a | 0.26 ± 0.02 d |
| Protein | 9.3 ± 0.5 c | 23.6 ± 0.2 a | 15.1 ± 0.2 b | 10.2 ± 0.3 c |
| Lipid | 0.9 ± 0.1 b | 0.7 ± 0.1 b,c | 3.0 ± 0.3 a | 0.5 ± 0.1 c |
| Caloric content (kcal/100 g dw) | 167.3 | 112.7 | 162.6 | 198.3 |
| Compound | Compound Content (mg/kg dw) | |||
|---|---|---|---|---|
| U. rigida | Gracilaria sp. | F. vesiculosus | S. latissima | |
| Saturated Fatty Acids | ||||
| Myristic acid | 77.9 ± 34.7 b | 100.5 ± 49.5 b | 927.9 ± 114.6 a | 185.2 ± 12.5 b |
| Palmitic acid | 595.3 ± 67.8 b | 458.1 ± 146.0 b,c | 1298.1 ± 106.2 a | 361.3 ± 22.2 c |
| Stearic acid | 85.6 ± 17.0 c | 98.4 ± 1.9 b,c | 539.9 ± 37.9 a | 146.4 ± 4.3 b |
| Unsaturated Fatty Acids | ||||
| Palmitoleic acid | 86.4 ± 37.2 b | 39.8 ± 14.2 b | 154.8 ± 19.8 a | 60.4 ± 6.0 b |
| Oleic acid | 410.2 ± 60.7 b | 136.7 ± 7.7 c | 756.1 ± 117.7 a | 134.4 ± 16.4 c |
| Linoleic acid | 97.4 ± 24.1 c | 59.9 ± 14.5 c | 815.0 ± 45.2 a | 240.3 ± 11.7 b |
| α-Linolenic acid | 261.9 ± 31.3 b | 216.2 ± 48.3 b | 1760.7 ± 151.0 a | 385.0 ± 13.1 b |
| Eicosapentaenoic acid | v | 61.9 ± 7.3 c | 276.8 ± 15.7 a | 111.5 ± 5.3 b |
| Esters of Fatty Acids | ||||
| Butyl octadecatetraenoate | 160.8 ± 42.2 a | 40.3 ± 6.8 b | 182.4 ± 14.1 a | 61.5 ± 4.7 b |
| Long Chain Alcohols | ||||
| 2-Hexadecen-1-ol | 85.5 ± 4.4 b | 22.9 ± 18.2 c | 123.8 ± 20.3 a | 14 ± 2.3 c |
| Diterpenes | ||||
| Phytol | 30.8 ± 11.1 c | 279.7 ± 24.8 a | 115.6 ± 16.8 b | 16.6 ± 3.0 c |
| Neophytadiene | 172.3 ± 7.6 a | 39.5 ± 9.7 b | 182.5 ± 35.3 a | 27.7 ± 2.8 b |
| Sterols | ||||
| Cholesterol derivatives | 87.8 ± 10.0 b | 888.1 ± 185.6 a | 921.2 ± 136.9 a | 202.3 ± 24.3 b |
| ∑ Saturated fatty acids | 758.8 ± 119.5 b | 657.0 ± 197.4 b | 2765.9 ± 257.7 a | 692.9 ± 39.0 b |
| ∑ Unsaturated fatty acids | 855.9 ± 153.3 b,c | 514.5 ± 92.0 c | 3763.4 ± 349.4 a | 931.6 ± 52.5 b |
| ∑ Ω3 | 261.9 ± 31.3 c | 278.1 ± 55.6 b,c | 2037.5 ± 166.7 a | 496.5 ± 18.4 b |
| ∑ Ω6 | 97.4 ± 24.1 c | 59.9 ± 14.5 c | 815.0 ± 45.2 a | 240.3 ± 11.7 b |
| Ω6/Ω3 | 0.37 | 0.22 | 0.40 | 0.48 |
| ∑ Sterols | 87.8 ± 10.0 b | 888.1 ± 185.6 a | 921.2 ± 136.9 a | 202.3 ± 24.3 b |
| Sample | Extract | Yield (g/100 g Seaweed dw) | TPC (g GAE/100 g Extract) | DPPH (g AAE/100 g Extract) | FRAP (g BHAE/100 g Extract) |
|---|---|---|---|---|---|
| U. rigida | RTWE | 43.1 ± 0.7 b | 0.23 ± 0.03 b | nd | 0.08 ± 0.02 d |
| HWE | 46.2 ± 0.6 a | 0.16 ± 0.02 b | nd | 0.17 ± 0.03 c | |
| Eth80 | 18.9 ± 1.8 c | 0.53 ± 0.07 a | 0.09 ± 0.01 a | 0.67 ± 0.04 a | |
| Acet70 | 16.2 ± 0.4 c | 0.48 ± 0.06 a | 0.08 ± 0.01 a | 0.53 ± 0.03 b | |
| Gracilaria sp. | RTWE | 30.7 ± 1.4 a | 0.59 ± 0.03 a | nd | nd |
| Eth80 | 23.3 ± 0.8 b | 0.49 ± 0.03 b | 0.04 ± 0.01 a | 0.23 ± 0.02 a | |
| Acet70 | 23.2 ± 1.6 b | 0.54 ± 0.01 a,b | 0.04 ± 0.00 a | 0.21 ± 0.03 a | |
| F. vesiculosus | RTWE | 27.8 ± 2.3 a | 1.48 ± 0.11 c | 0.57 ± 0.06 c | 2.90 ± 1.02 b |
| HWE | 30.5 ± 0.8 a | 1.74 ± 0.09 c | 0.77 ± 0.12 c | 3.23 ± 1.00 b | |
| Eth80 | 15.3 ± 1.8 b | 5.66 ± 0.26 a | 2.60 ± 0.28 a | 8.30 ± 1.04 a | |
| Acet70 | 18.2 ± 0.9 b | 3.91 ± 0.09 b | 1.47 ± 0.12 b | 8.03 ± 3.91 a | |
| S. latissima | RTWE | 40.2 ± 1.6 a,b,c | 0.47 ± 0.02 b | 0.20 ± 0.04 a | 0.68 ± 0.02 c |
| HWE | 44.3 ± 0.9 a | 0.78 ± 0.06 a | 0.25 ± 0.05 a | 1.06 ± 0.06 a | |
| Eth80 | 36.5 ± 2.2 b,c | 0.19 ± 0.02 c | 0.07 ± 0.02 b | 0.21 ± 0.02 d | |
| Acet70 | 37.5 ± 2.8 b | 0.52 ± 0.02 b | 0.25 ± 0.01 a | 0.87 ± 0.07 b |
| Sample | α-Glucosidase | Lipase | ACE | ||||
|---|---|---|---|---|---|---|---|
| IC50 | %inb | IC50 | %inb | IC50 | %inb | ||
| Seaweed extract | |||||||
| U. rigida | RTWE | - | ni | - | ni | - | 24.7 ± 5.0% (4.6) |
| HWE | - | ni | - | ni | - | 24.3 ± 7.1% (4.8) | |
| Eth80 | - | ni | - | ni | 3.67 ± 0.14 b | - | |
| Acet70 | - | ni | - | ni | 4.33 ± 0.30 a | - | |
| Gracilaria sp. | RTWE | - | ni | - | ni | - | 15.8 ± 4.3% (4.7) |
| Eth80 | - | ni | - | 26.1± 3.7% (3.0) | 3.21 ± 0.23 a | - | |
| Acet70 | - | ni | - | ni | 3.45 ± 0.26 a | - | |
| F. vesiculosus | RTWE | 6.73 ± 0.74 c | - | - | 25.0 ± 2.8% (3.0) | - | ni |
| HWE | 5.12 ± 0.81 b | - | - | 15.2 ± 2.5% (3.0) | - | ni | |
| Eth80 | 0.119 ± 0.036 b | - | - | ni | - | ni | |
| Acet70 | 0.032 ± 0.003 a | - | - | 19.5 ± 3.1% (3.0) | - | ni | |
| S. latissima | RTWE | 6.73 ± 0.74 c | - | - | ni | 4.48 ± 0.66 b | - |
| HWE | 5.12 ± 0.81 b | - | - | ni | - | 39.6 ± 2.3% (4.7) | |
| Eth80 | - | 47.8 ± 4.5% (7.3) | - | 30.8 ± 4.1% (3.0) | 4.56 ± 0.05 a | - | |
| Acet70 | 1.68 ± 0.22 a | - | - | ni | 3.98 ± 0.29 b | - | |
| Reference compound | |||||||
| Acarbose | 0.264 ± 0.041 | - | - | - | - | - | |
| Orlistat | - | - | 0.075 ± 0.004 | - | - | - | |
| Captopril | - | - | - | - | 7.8 × 10−7 ± 1.1 × 10−7 | - | |
| Seaweed | Extract | g Ash/100 g extdw | g GlcE/100 g extdw | g Sol Fiber/100 g extdw | g Protein/100 g extdw |
|---|---|---|---|---|---|
| U. rigida | RTWE | 48.5 ± 0.7 b | 19.6 ± 0.6 b | 33.8 ± 3.0 a | 5.2 ± 0.1 c |
| HWE | 49.2 ± 0.5 b | 19.4 ± 1.2 b | 37.3 ± 1.1 a | 3.9 ± 0.3 d | |
| Gracilaria sp. | RTWE | 55.9 ± 1.0 a | 8.9 ± 1.5 e | 14.9 ± 0.6 d | 19.0 ± 0.1 a |
| F. vesiculosus | RTWE | 28.8 ± 0.3 c | 7.5 ± 1.0 e | 17.2 ± 0.5 c | 7.0 ± 0.5 b |
| HWE | 29.4 ± 0.4 c | 13.8 ± 2.1 d | 24.5 ± 2.8 b | 6.6 ± 0.4 b | |
| S. latíssima | RTWE | 29.7 ± 0.4 c | 18.2 ± 0.7 b | 17.1 ± 0.9 c | 4.4 ± 0.1 d |
| HWE | 27.8 ± 0.1 c | 23.4 ± 3.1 a | 25.2 ± 0.2 b | 4.6 ± 0.2 d |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Neto, R.T.; Marçal, C.; Queirós, A.S.; Abreu, H.; Silva, A.M.S.; Cardoso, S.M. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. Int. J. Mol. Sci. 2018, 19, 2987. https://doi.org/10.3390/ijms19102987
Neto RT, Marçal C, Queirós AS, Abreu H, Silva AMS, Cardoso SM. Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. International Journal of Molecular Sciences. 2018; 19(10):2987. https://doi.org/10.3390/ijms19102987
Chicago/Turabian StyleNeto, Rodrigo T., Catarina Marçal, Ana S. Queirós, Helena Abreu, Artur M. S. Silva, and Susana M. Cardoso. 2018. "Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients" International Journal of Molecular Sciences 19, no. 10: 2987. https://doi.org/10.3390/ijms19102987
APA StyleNeto, R. T., Marçal, C., Queirós, A. S., Abreu, H., Silva, A. M. S., & Cardoso, S. M. (2018). Screening of Ulva rigida, Gracilaria sp., Fucus vesiculosus and Saccharina latissima as Functional Ingredients. International Journal of Molecular Sciences, 19(10), 2987. https://doi.org/10.3390/ijms19102987