Whole-Body 12C Irradiation Transiently Decreases Mouse Hippocampal Dentate Gyrus Proliferation and Immature Neuron Number, but Does Not Change New Neuron Survival Rate

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. 12C Irradiation Had No Overt Physiological Influence on IRR Mice Relative to Sham Mice
2.2. 24-h Post-Irradiation, IRR Mice Had 57% Fewer Ki67+ Cells and 59% Fewer Ki67+ Clusters in the DG GCL Relative to Sham Mice
2.3. 24-h Post-Irradiation, IRR Mice Had 50% Fewer BrdU+ Cells and 59% Fewer BrdU+ Clusters in the DG GCL Relative to Sham Mice
2.4. 24-h Post-Irradiation, IRR Mice Had 33% Fewer DCX+ Cells in the DG GCL Relative to Sham Mice
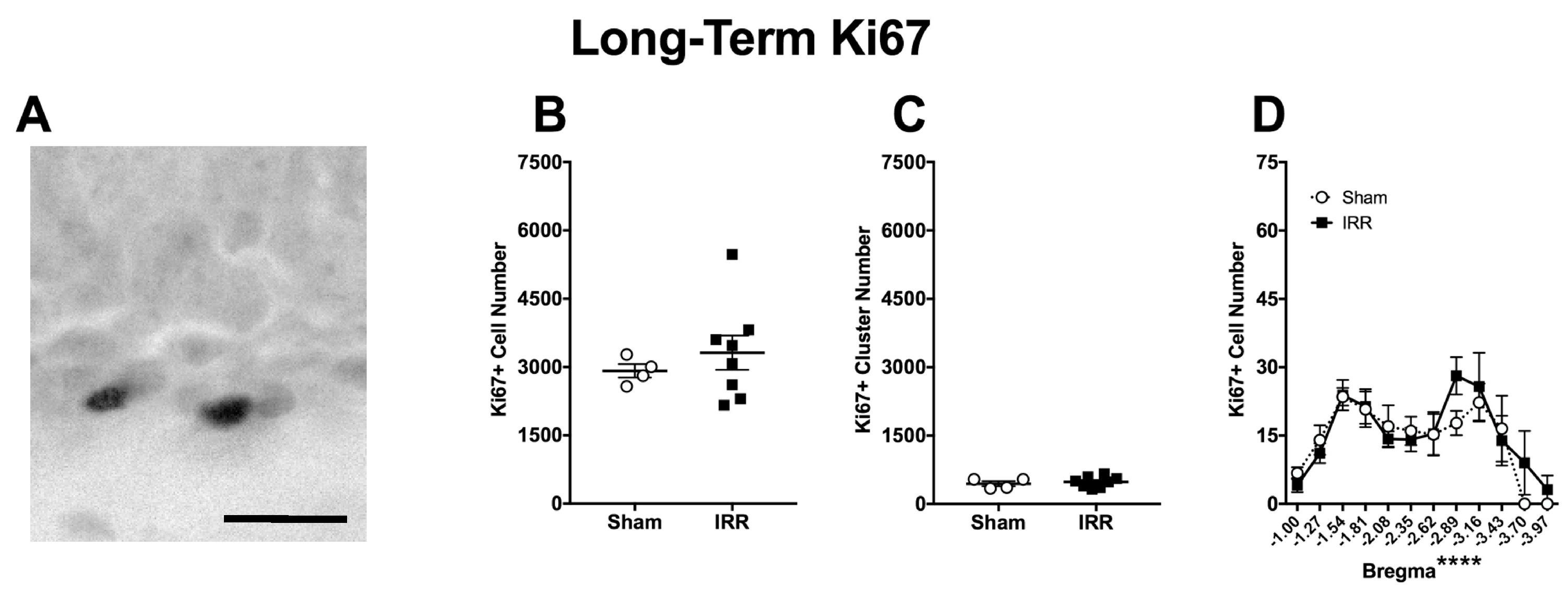
2.5. Three-mon Post-Irradiation, IRR Mice Had a Similar Amount of Ki67+ Cells and Clusters When Compared to Sham
2.6. Three-mon Post-Irradiation, IRR Mice Had 64% Fewer BrdU+ Cells, but a Similar Number of Clusters When Compared to Sham Mice
2.7. Three-mon Post-Irradiation, IRR Mice Had Similar Number of DCX+ Cells Compared to Sham Mice
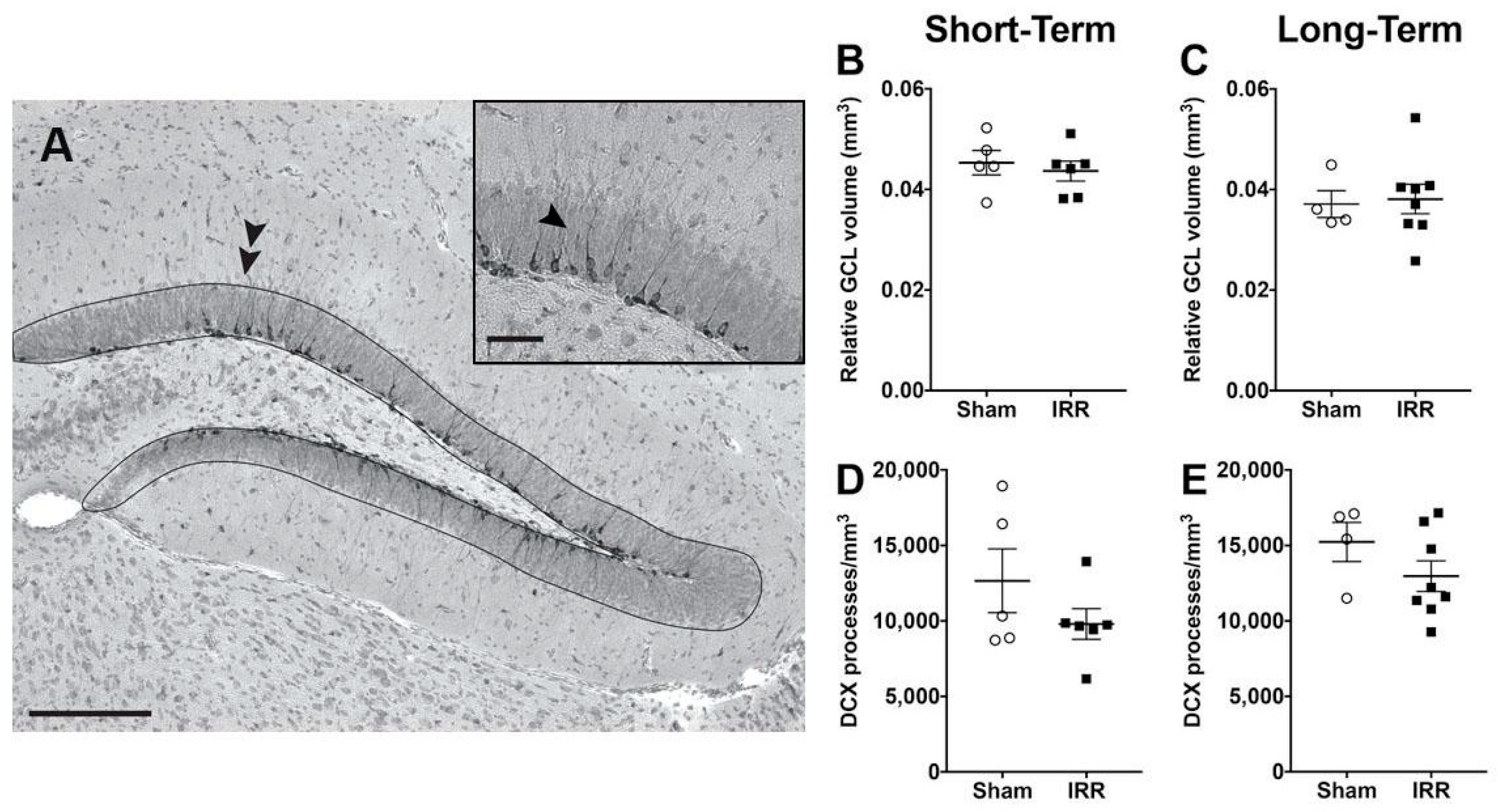
2.8. 24-h and 3-mon Post-Irradiation, IRR Mice Had Similar GCL Volume and Similar Density of Immature Neuron Dendritic Processes Compared to Sham Mice
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Irradiation Procedures
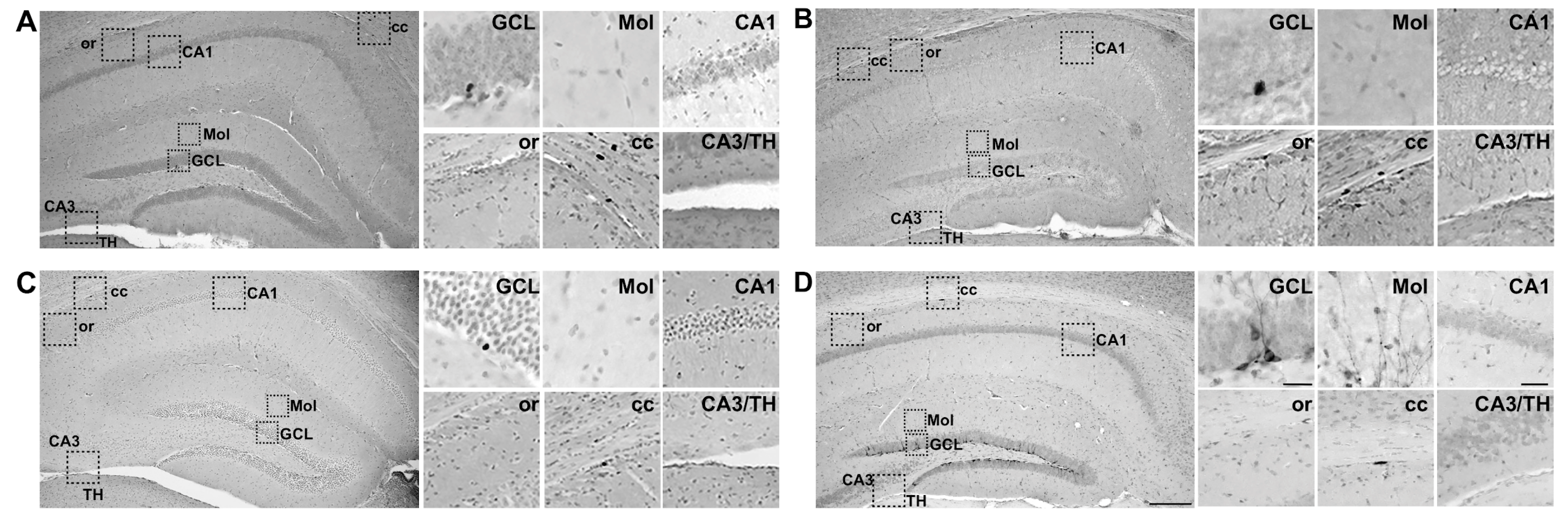
4.3. Tissue Preparation and Immunohistochemistry (IHC)
4.4. Stereological Cell Counts
4.5. Stereological Estimation of GCL Volume and Density of Immature Neuron Dendritic Processes
4.6. Statistical Analyses and Image Presentation
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BNL | Brookhaven National Laboratories |
| BrdU | bromodeoxyuridine |
| CA1 | cornus ammonis 1 |
| CA3 | cornus ammonis 3 |
| cc | corpus callosum |
| DAB | diaminobenzidine |
| DCX | doublecortin |
| DG | dentate gyrus |
| GCL | granule cell layer |
| h | hour |
| HZE | high-energy and high-charge |
| IHC | immunohistochemistry |
| IRR | irradiated |
| Mol | molecular layer |
| mon | month |
| NSC | neural stem cell |
| NSRL | NASA Space Radiation Laboratory |
| or | stratum oriens |
| SEM | standard error of the mean |
| SGZ | subgranular zone |
| TH | thalamus |
| UTSW | University of Texas Southwestern Medical Center |
References
- Altman, J.; Das, G.D. Autoradiographic and histological evidence of postnatal hippocampal neurogenesis in rats. J. Comp. Neurol. 1965, 124, 319–335. [Google Scholar] [CrossRef] [PubMed]
- Altman, J. Autoradiographic and histological studies of postnatal neurogenesis. IV. Cell proliferation and migration in the anterior forebrain, with special reference to persisting neurogenesis in the olfactory bulb. J. Comp. Neurol. 1969, 137, 433–457. [Google Scholar] [CrossRef] [PubMed]
- Bond, A.M.; Ming, G.-L.; Song, H. Adult Mammalian Neural Stem Cells and Neurogenesis: Five Decades Later. Cell Stem Cell 2015, 17, 385–395. [Google Scholar] [CrossRef] [PubMed]
- Tanti, A.; Belzung, C. Neurogenesis along the septo-temporal axis of the hippocampus: Are depression and the action of antidepressants region-specific? Neuroscience 2013, 252, 234–252. [Google Scholar] [CrossRef] [PubMed]
- Toda, T.; Parylak, S.L.; Linker, S.B.; Gage, F.H. The role of adult hippocampal neurogenesis in brain health and disease. Mol. Psychiatry 2018. [Google Scholar] [CrossRef] [PubMed]
- Petrik, D.; Lagace, D.C.; Eisch, A.J. The neurogenesis hypothesis of affective and anxiety disorders: Are we mistaking the scaffolding for the building? Neuropharmacology 2012, 62, 21–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tannenholz, L.; Jimenez, J.C.; Kheirbek, M.A. Local and regional heterogeneity underlying hippocampal modulation of cognition and mood. Front. Behav. Neurosci. 2014, 8, 147. [Google Scholar] [CrossRef] [PubMed]
- Kang, E.; Wen, Z.; Song, H.; Christian, K.M.; Ming, G.-L. Adult Neurogenesis and Psychiatric Disorders. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Oomen, C.A.; Bekinschtein, P.; Kent, B.A.; Saksida, L.M.; Bussey, T.J. Adult hippocampal neurogenesis and its role in cognition. Wiley Interdiscip. Rev. Cogn. Sci. 2014, 5, 573–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hladik, D.; Tapio, S. Effects of ionizing radiation on the mammalian brain. Mutat. Res. 2016, 770, 219–230. [Google Scholar] [CrossRef] [PubMed]
- Raber, J. Unintended effects of cranial irradiation on cognitive function. Toxicol. Pathol. 2010, 38, 198–202. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.-F.; Gu, S.; Shan, C.; Marchado, S.; Arias-Carrión, O. Oxidative Stress and Adult Neurogenesis. Stem Cell Rev. 2015, 11, 706–709. [Google Scholar] [CrossRef] [PubMed]
- Huang, T.-T.; Leu, D.; Zou, Y. Oxidative stress and redox regulation on hippocampal-dependent cognitive functions. Arch. Biochem. Biophys. 2015, 576, 2–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubera, M.; Obuchowicz, E.; Goehler, L.; Brzeszcz, J.; Maes, M. In animal models, psychosocial stress-induced (neuro)inflammation, apoptosis and reduced neurogenesis are associated to the onset of depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 2011, 35, 744–759. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Miller, M.G.; Scott, T.; Shukitt-Hale, B. Nutritional Factors Affecting Adult Neurogenesis and Cognitive Function. Adv. Nutr. 2017, 8, 804–811. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Balentova, S.; Adamkov, M. Molecular, Cellular and Functional Effects of Radiation-Induced Brain Injury: A Review. Int. J. Mol. Sci. 2015, 16, 27796–27815. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Greene-Schloesser, D.; Robbins, M.E.; Peiffer, A.M.; Shaw, E.G.; Wheeler, K.T.; Chan, M.D. Radiation-induced brain injury: A. review. Front. Oncol. 2012, 2, 73. [Google Scholar] [CrossRef] [PubMed]
- Son, Y.; Yang, M.; Wang, H.; Moon, C. Hippocampal dysfunctions caused by cranial irradiation: A review of the experimental evidence. Brain Behav. Immun. 2015, 45, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Monje, M.L.; Palmer, T. Radiation injury and neurogenesis. Curr. Opin. Neurol. 2003, 16, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Monje, M. Cranial radiation therapy and damage to hippocampal neurogenesis. Dev. Disabil. Res. Rev. 2008, 14, 238–242. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, F.A.; Alp, M.; Sulzman, F.M.; Wang, M. Space radiation risks to the central nervous system. Life Sci. Space Res. 2014, 2, 54–69. [Google Scholar] [CrossRef] [Green Version]
- Rabin, B.M.; Shukitt-Hale, B.; Carrihill-Knoll, K.L. Effects of Age on the Disruption of Cognitive Performance by Exposure to Space Radiation. J. Behav. Brain Sci. 2014, 04, 297–307. [Google Scholar] [CrossRef]
- Rabin, B.M.; Buhler, L.L.; Joseph, J.A.; Shukitt-Hale, B.; Jenkins, D.G. Effects of exposure to 56Fe particles or protons on fixed-ratio operant responding in rats. J. Radiat. Res. 2002, 43, S225–S228. [Google Scholar] [CrossRef]
- Joseph, J.A.; Hunt, W.A.; Rabin, B.M.; Dalton, T.K. Possible “accelerated striatal aging” induced by 56Fe heavy-particle irradiation: Implications for manned space flights. Radiat. Res. 1992, 130, 88–93. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Casadesus, G.; McEwen, J.J.; Rabin, B.M.; Joseph, J.A. Spatial learning and memory deficits induced by exposure to iron-56-particle radiation. Radiat. Res. 2000, 154, 28–33. [Google Scholar] [CrossRef]
- Shukitt-Hale, B.; Casadesus, G.; Carey, A.N.; Rabin, B.M.; Joseph, J.A. Exposure to 56Fe irradiation accelerates normal brain aging and produces deficits in spatial learning and memory. Adv. Space Res. 2007, 39, 1087–1092. [Google Scholar] [CrossRef]
- Raber, J.; Rosi, S.; Chakraborti, A.; Fishman, K.; Dayger, C.; Davis, M.J.; Villasana, L.; Fike, J.R. Effects of 56Fe-Particle Cranial Radiation on Hippocampus-Dependent Cognition Depend on the Salience of the Environmental Stimuli. Radiat. Res. 2011, 176, 521–526. [Google Scholar] [CrossRef] [PubMed]
- Joseph, J.A.; Hunt, W.A.; Rabin, B.M.; Dalton, T.K.; Harris, A.H. Deficits in the sensitivity of striatal muscarinic receptors induced by 56Fe heavy-particle irradiation: Further “age-radiation” parallels. Radiat. Res. 1993, 135, 257–261. [Google Scholar] [CrossRef] [PubMed]
- Nelson, G.A. Fundamental space radiobiology. Gravitat. Space Res. 2003, 16, 29–36. [Google Scholar]
- Durante, M. Space radiation protection: Destination Mars. Life Sci. Space Res. 2014, 1, 2–9. [Google Scholar] [CrossRef] [PubMed]
- Gemignani, J.; Gheysens, T.; Summerer, L. Beyond astronaut’s capabilities: The current state of the art. Conf. Proc. IEEE Eng. Med. Biol. Soc. 2015, 2015, 3615–3618. [Google Scholar] [PubMed]
- Hagen, U. Radiation biology in space: A critical review. Adv. Space Res. 1989, 9, 3–8. [Google Scholar] [CrossRef]
- Bucker, H.; Facius, R. The role of HZE particles in space flight: Results from spaceflight and ground-based experiments. Acta Astronaut. 1981, 8, 1099–1107. [Google Scholar] [CrossRef]
- Baek, S.-J.; Ishii, H.; Tamari, K.; Hayashi, K.; Nishida, N.; Konno, M.; Kawamoto, K.; Koseki, J.; Fukusumi, T.; Hasegawa, S.; et al. Cancer stem cells: The potential of carbon ion beam radiation and new radiosensitizers (Review). Oncol. Rep. 2015, 34, 2233–2237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schlaff, C.D.; Krauze, A.; Belard, A.; O’Connell, J.J.; Camphausen, K.A. Bringing the heavy: Carbon ion therapy in the radiobiological and clinical context. Radiat. Oncol. 2014, 9. [Google Scholar] [CrossRef] [PubMed]
- Jiang, G.-L. Particle therapy for cancers: A new weapon in radiation therapy. Front. Med. 2012, 6, 165–172. [Google Scholar] [CrossRef] [PubMed]
- Walenta, S.; Mueller-Klieser, W. Differential Superiority of Heavy Charged-Particle Irradiation to X-rays: Studies on Biological Effectiveness and Side Effect Mechanisms in Multicellular Tumor and Normal Tissue Models. Front. Oncol. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Jones, B. The case for particle therapy. Br. J. Radiol. 2006, 79, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Rola, R.; Sarkissian, V.; Obenaus, A.; Nelson, G.A.; Otsuka, S.; Limoli, C.L.; Fike, J.R. High-LET Radiation Induces Inflammation and Persistent Changes in Markers of Hippocampal Neurogenesis. Radiat. Res. 2005, 164, 556–560. [Google Scholar] [CrossRef] [PubMed]
- DeCarolis, N.A.; Rivera, P.D.; Ahn, F.; Amaral, W.Z.; LeBlanc, J.A.; Malhotra, S.; Shih, H.Y.; Petrik, D.; Melvin, N.; Chen, B.P.; et al. Fe Particle Exposure Results in a Long-Lasting Increase in a Cellular Index of Genomic Instability and Transiently Suppresses Adult Hippocampal Neurogenesis. Life Sci. Space Res. 2014, 2, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Whoolery, C.W.; Walker, A.K.; Richardson, D.R.; Lucero, M.J.; Reynolds, R.P.; Beddow, D.H.; Clark, K.L.; Shih, H.-Y.; LeBlanc, J.A.; Cole, M.G.; et al. Whole-Body Exposure to 28Si-Radiation Dose-Dependently Disrupts Dentate Gyrus Neurogenesis and Proliferation in the Short Term and New Neuron Survival and Contextual Fear Conditioning in the Long Term. Radiat. Res. 2017, 188, 532–551. [Google Scholar] [CrossRef] [PubMed]
- O’Leary, O.F.; Cryan, J.F. A ventral view on antidepressant action: Roles for adult hippocampal neurogenesis along the dorsoventral axis. Trends Pharmacol. Sci. 2014, 35, 675–687. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.V.; Sahay, A.; Duman, R.S.; Hen, R. Functional differentiation of adult-born neurons along the septotemporal axis of the dentate gyrus. Cold Spring Harb. Perspect. Biol. 2015, 7, a018978. [Google Scholar] [CrossRef] [PubMed]
- Snyder, J.S.; Ferrante, S.C.; Cameron, H.A. Late maturation of adult-born neurons in the temporal dentate gyrus. PLoS ONE 2012, 7, e48757. [Google Scholar] [CrossRef] [PubMed]
- Moser, M.B.; Moser, E.I. Functional differentiation in the hippocampus. Hippocampus 1998, 8, 608–619. [Google Scholar] [CrossRef]
- Fanselow, M.S.; Dong, H.-W. Are the dorsal and ventral hippocampus functionally distinct structures? Neuron 2010, 65, 7–19. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Carrihill-Knoll, K.L.; Shukitt-Hale, B. Operant responding following exposure to HZE particles and its relationship to particle energy and linear energy transfer. Adv. Space Res. 2011, 48, 370–377. [Google Scholar] [CrossRef]
- Raber, J.; Allen, A.R.; Sharma, S.; Allen, B.; Rosi, S.; Olsen, R.H.J.; Davis, M.J.; Eiwaz, M.; Fike, J.R.; Nelson, G.A. Effects of Proton and Combined Proton and 56Fe Radiation on the Hippocampus. Radiat. Res. 2015, 185, 20–30. [Google Scholar] [CrossRef] [PubMed]
- Bellone, J.A.; Rudobeck, E.; Hartman, R.E.; Szücs, A.; Vlkolinský, R. A Single Low Dose of Proton Radiation Induces Long-Term Behavioral and Electrophysiological Changes in Mice. Radiat. Res. 2015, 184, 193–202. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Allen, B.D.; Tran, K.K.; Chmielewski, N.N.; Craver, B.M.; Martirosian, V.; Morganti, J.M.; Rosi, S.; Vlkolinsky, R.; Acharya, M.M.; et al. Targeted overexpression of mitochondrial catalase prevents radiation-induced cognitive dysfunction. Antioxid. Redox Signal. 2015, 22, 78–91. [Google Scholar] [CrossRef] [PubMed]
- Marty, V.N.; Vlkolinsky, R.; Minassian, N.; Cohen, T.; Nelson, G.A.; Spigelman, I. Radiation-induced alterations in synaptic neurotransmission of dentate granule cells depend on the dose and species of charged particles. Radiat. Res. 2014, 182, 653–665. [Google Scholar] [PubMed]
- Sweet, T.B.; Panda, N.; Hein, A.M.; Das, S.L.; Hurley, S.D.; Olschowka, J.A.; Williams, J.P.; O’Banion, M.K. Central nervous system effects of whole-body proton irradiation. Radiat. Res. 2014, 182, 18–34. [Google Scholar] [CrossRef] [PubMed]
- Kiffer, F.; Howe, A.K.; Carr, H.; Wang, J.; Alexander, T.; Anderson, J.E.; Groves, T.; Seawright, J.W.; Sridharan, V.; Carter, G.; et al. Late effects of 1H irradiation on hippocampal physiology. Life Sci. Space Res. 2018, 17, 51–62. [Google Scholar] [CrossRef] [PubMed]
- Kiffer, F.; Carr, H.; Groves, T.; Anderson, J.E.; Alexander, T.; Wang, J.; Seawright, J.W.; Sridharan, V.; Carter, G.; Boerma, M.; et al. Effects of 1H + 16O Charged Particle Irradiation on Short-Term Memory and Hippocampal Physiology in a Murine Model. Radiat. Res. 2018, 189, 53–63. [Google Scholar] [PubMed]
- Impey, S.; Jopson, T.; Pelz, C.; Tafessu, A.; Fareh, F.; Zuloaga, D.; Marzulla, T.; Riparip, L.-K.; Stewart, B.; Rosi, S.; et al. Bi-directional and shared epigenomic signatures following proton and 56Fe irradiation. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Patel, R.; Arakawa, H.; Radivoyevitch, T.; Gerson, S.L.; Welford, S.M. Long-Term Deficits in Behavior Performances Caused by Low- and High-Linear Energy Transfer Radiation. Radiat. Res. 2017, 188, 672–680. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.; Pelz, C.; Tafessu, A.; Marzulla, T.; Turker, M.S.; Raber, J. Proton irradiation induces persistent and tissue-specific DNA methylation changes in the left ventricle and hippocampus. BMC Genom. 2016, 17. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Marzulla, T.; Stewart, B.; Kronenberg, A.; Turker, M.S. 28Silicon Irradiation Impairs Contextual Fear Memory in B6D2F1 Mice. Radiat. Res. 2015, 183, 708–712. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Heroux, N.A.; Shukitt-Hale, B.; Carrihill-Knoll, K.L.; Beck, Z.; Baxter, C. Lack of reliability in the disruption of cognitive performance following exposure to protons. Radiat. Environ. Biophys. 2015, 54, 285–295. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Carrihill-Knoll, K.L.; Miller, M.G.; Shukitt-Hale, B. Age as a factor in the responsiveness of the organism to the disruption of cognitive performance by exposure to HZE particles differing in linear energy transfer. Life Sci. Space Res. 2018, 16, 84–92. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Maroso, M.; Syage, A.; Allen, B.D.; Angulo, M.C.; Soltesz, I.; Limoli, C.L. Persistent nature of alterations in cognition and neuronal circuit excitability after exposure to simulated cosmic radiation in mice. Exp. Neurol. 2018, 305, 44–55. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Carrihill-Knoll, K.L.; Shukitt-Hale, B. Comparison of the Effectiveness of Exposure to Low-LET Helium Particles (4He) and Gamma Rays (137Cs) on the Disruption of Cognitive Performance. Radiat. Res. 2015, 184, 266–272. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Torres, E.R.S.; Akinyeke, T.; Lee, J.; Weber Boutros, S.J.; Turker, M.S.; Kronenberg, A. Detrimental Effects of Helium Ion Irradiation on Cognitive Performance and Cortical Levels of MAP-2 in B6D2F1 Mice. Int. J. Mol. Sci. 2018, 19, 1247. [Google Scholar] [CrossRef] [PubMed]
- Cacao, E.; Cucinotta, F.A. Modeling Heavy-Ion Impairment of Hippocampal Neurogenesis after Acute and Fractionated Irradiation. Radiat. Res. 2016, 186, 624–637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Belov, O.V.; Belokopytova, K.V.; Bazyan, A.S.; Kudrin, V.S.; Narkevich, V.B.; Ivanov, A.A.; Severiukhin, Y.S.; Timoshenko, G.N.; Krasavin, E.A. Exposure to 12C particles alters the normal dynamics of brain monoamine metabolism and behaviour in rats. Phys. Med. 2016, 32, 1088–1094. [Google Scholar] [CrossRef] [PubMed]
- Carr, H.; Alexander, T.C.; Groves, T.; Kiffer, F.; Wang, J.; Price, E.; Boerma, M.; Allen, A.R. Early effects of 16O radiation on neuronal morphology and cognition in a murine model. Life Sci. Space Res. 2018, 17, 63–73. [Google Scholar]
- Mange, A.; Cao, Y.; Zhang, S.; Hienz, R.D.; Davis, C.M. Whole-Body Oxygen 16O Ion-Exposure-Induced Impairments in Social Odor Recognition Memory in Rats Are Dose and Time Dependent. Radiat. Res. 2018, 189, 292–299. [Google Scholar] [PubMed]
- Parihar, V.K.; Allen, B.D.; Caressi, C.; Kwok, S.; Chu, E.; Tran, K.K.; Chmielewski, N.N.; Giedzinski, E.; Acharya, M.M.; et al. Cosmic radiation exposure and persistent cognitive dysfunction. Sci. Rep. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- Parihar, V.K.; Allen, B.; Tran, K.K.; Macaraeg, T.G.; Chu, E.M.; Kwok, S.F.; Chmielewski, N.N.; Craver, B.M.; Baulch, J.E.; Acharya, M.M.; et al. What happens to your brain on the way to Mars. Sci Adv 2015, 1. [Google Scholar] [CrossRef]
- Raber, J.; Marzulla, T.; Kronenberg, A.; Turker, M.S. 16Oxygen irradiation enhances cued fear memory in B6D2F1 mice. Life Sci. Space Res. 2015, 7, 61–65. [Google Scholar]
- Rabin, B.M.; Poulose, S.M.; Carrihill-Knoll, K.L.; Ramirez, F.; Bielinski, D.F.; Heroux, N.; Shukitt-Hale, B. Acute Effects of Exposure to 56Fe and 16O Particles on Learning and Memory. Radiat. Res. 2015, 184, 143–150. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Shukitt-Hale, B.; Carrihill-Knoll, K.L.; Gomes, S.M. Comparison of the Effects of Partial- or Whole-Body Exposures to 16O Particles on Cognitive Performance in Rats. Radiat. Res. 2014, 181, 251–257. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Bielinski, D.F.; Carrihill-Knoll, K.; Rabin, B.M.; Shukitt-Hale, B. Exposure to 16O-particle radiation causes aging-like decrements in rats through increased oxidative stress, inflammation and loss of autophagy. Radiat. Res. 2011, 176, 761–769. [Google Scholar] [CrossRef] [PubMed]
- Krukowski, K.; Jones, T.; Campbell-Beachler, M.; Nelson, G.; Rosi, S. Peripheral T Cells as a Biomarker for Oxygen-Ion-Radiation-Induced Social Impairments. Radiat. Res. 2018, 190, 186–193. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Rudobeck, E.; Campbell-Beachler, M.; Allen, A.R.; Allen, B.; Rosi, S.; Nelson, G.A.; Ramachandran, S.; Turner, J.; Fike, J.R.; et al. 28Silicon Radiation-Induced Enhancement of Synaptic Plasticity in the Hippocampus of Naïve and Cognitively Tested Mice. Radiat. Res. 2014, 181, 362–368. [Google Scholar] [CrossRef] [PubMed]
- Rudobeck, E.; Nelson, G.A.; Sokolova, I.V.; Vlkolinský, R. 28Silicon Radiation Impairs Neuronal Output in CA1 Neurons of Mouse Ventral Hippocampus without Altering Dendritic Excitability. Radiat. Res. 2014, 181, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Jewell, J.S.; Duncan, V.D.; Hadley, M.M.; Macadat, E.; Musto, A.E.; Tessa, C.L. Impaired Attentional Set-Shifting Performance after Exposure to 5 cGy of 600 MeV/n 28Si Particles. Radiat. Res. 2018, 189, 273–282. [Google Scholar] [CrossRef] [PubMed]
- Acharya, M.M.; Baddour, A.A.D.; Kawashita, T.; Allen, B.D.; Syage, A.R.; Nguyen, T.H.; Yoon, N.; Giedzinski, E.; Yu, L.; Parihar, V.K.; et al. Epigenetic determinants of space radiation-induced cognitive dysfunction. Sci. Rep. 2017, 7. [Google Scholar] [CrossRef] [PubMed]
- Hadley, M.M.; Davis, L.K.; Jewell, J.S.; Miller, V.D.; Britten, R.A. Exposure to Mission-Relevant Doses of 1 GeV/n 48Ti Particles Impairs Attentional Set-Shifting Performance in Retired Breeder Rats. Radiat. Res. 2016, 185, 13–19. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Allen, A.R.; Weber, S.; Chakraborti, A.; Sharma, S.; Fike, J.R. Effect of behavioral testing on spine density of basal dendrites in the CA1 region of the hippocampus modulated by 56Fe irradiation. Behav. Brain Res. 2016, 302, 263–268. [Google Scholar] [CrossRef] [PubMed]
- Sweet, T.B.; Hurley, S.D.; Wu, M.D.; Olschowka, J.A.; Williams, J.P.; O’Banion, M.K. Neurogenic Effects of Low-Dose Whole-Body HZE (Fe) Ion and Gamma Irradiation. Radiat. Res. 2016, 186, 614–623. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Allen, A.R.; Raber, J.; Chakraborti, A.; Sharma, S.; Fike, J.R. 56Fe Irradiation Alters Spine Density and Dendritic Complexity in the Mouse Hippocampus. Radiat. Res. 2015, 184, 586–594. [Google Scholar] [CrossRef] [PubMed]
- Raber, J.; Allen, A.R.; Rosi, S.; Sharma, S.; Dayger, C.; Davis, M.J.; Fike, J.R. Effects of whole body 56Fe radiation on contextual freezing and Arc-positive cells in the dentate gyrus. Behav. Brain Res. 2013, 246, 162–167. [Google Scholar] [CrossRef] [PubMed]
- Rivera, P.D.; Shih, H.Y.; Leblanc, J.A.; Cole, M.G.; Amaral, W.Z.; Mukherjee, S.; Zhang, S.; Lucero, M.J.; Decarolis, N.A.; Chen, B.P.; et al. Acute and fractionated exposure to high-LET 56Fe HZE-particle radiation both result in similar long-term deficits in adult hippocampal neurogenesis. Radiat. Res. 2013, 180, 658–667. [Google Scholar] [CrossRef] [PubMed]
- Cherry, J.D.; Liu, B.; Frost, J.L.; Lemere, C.A.; Williams, J.P.; Olschowka, J.A.; O’Banion, M.K. Galactic cosmic radiation leads to cognitive impairment and increased aβ plaque accumulation in a mouse model of Alzheimer’s disease. PLoS ONE 2012, 7, e53275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mao, X.W.; Favre, C.J.; Fike, J.R.; Kubinova, L.; Anderson, E.; Campbell-Beachler, M.; Jones, T.; Smith, A.; Rightnar, S.; Nelson, G.A. High-LET radiation-induced response of microvessels in the Hippocampus. Radiat. Res. 2010, 173, 486–493. [Google Scholar] [CrossRef] [PubMed]
- Encinas, J.M.; Vazquez, M.E.; Switzer, R.C.; Chamberland, D.W.; Nick, H.; Levine, H.G.; Scarpa, P.J.; Enikolopov, G.; Steindler, D.A. Quiescent adult neural stem cells are exceptionally sensitive to cosmic radiation. Exp. Neurol. 2008, 210, 274–279. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Manda, K.; Ueno, M.; Anzai, K. Memory impairment, oxidative damage and apoptosis induced by space radiation: Ameliorative potential of alpha-lipoic acid. Behav. Brain Res. 2008, 187, 387–395. [Google Scholar] [CrossRef] [PubMed]
- Manda, K.; Ueno, M.; Anzai, K. Space radiation-induced inhibition of neurogenesis in the hippocampal dentate gyrus and memory impairment in mice: Ameliorative potential of the melatonin metabolite, AFMK. J. Pineal Res. 2008, 45, 430–438. [Google Scholar] [CrossRef] [PubMed]
- Rola, R.; Fishman, K.; Baure, J.; Rosi, S.; Lamborn, K.R.; Obenaus, A.; Nelson, G.A.; Fike, J.R. Hippocampal Neurogenesis and Neuroinflammation after Cranial Irradiation with 56Fe Particles. Radiat. Res. 2008, 169, 626–632. [Google Scholar] [CrossRef] [PubMed]
- Pecaut, M.J.; Haerich, P.; Miller, C.N.Z.; Smith, A.L.; Zendejas, E.D.; Nelson, G.A. The effects of low-dose, high-LET radiation exposure on three models of behavior in C57BL/6 mice. Radiat. Res. 2004, 162, 148–156. [Google Scholar] [CrossRef] [PubMed]
- Rola, R.; Otsuka, S.; Obenaus, A.; Nelson, G.A.; Limoli, C.L.; VandenBerg, S.R.; Fike, J.R. Indicators of hippocampal neurogenesis are altered by 56Fe-particle irradiation in a dose-dependent manner. Radiat. Res. 2004, 162, 442–446. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Jewell, J.S.; Davis, L.K.; Miller, V.D.; Hadley, M.M.; Semmes, O.J.; Lonart, G.; Dutta, S.M. Changes in the Hippocampal Proteome Associated with Spatial Memory Impairment after Exposure to Low (20 cGy) Doses of 1 GeV/n 56Fe Radiation. Radiat. Res. 2017, 187, 287–297. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Miller, V.D.; Hadley, M.M.; Jewell, J.S.; Macadat, E. Performance in hippocampus-and PFC-dependent cognitive domains are not concomitantly impaired in rats exposed to 20 cGy of 1 GeV/n 56Fe particles. Life Sci. Space Res. 2016, 10, 17–22. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Jewell, J.S.; Miller, V.D.; Davis, L.K.; Hadley, M.M.; Wyrobek, A.J. Impaired Spatial Memory Performance in Adult Wistar Rats Exposed to Low (5–20 cGy) Doses of 1 GeV/n 56Fe Particles. Radiat. Res. 2016, 185, 332–337. [Google Scholar] [CrossRef] [PubMed]
- Impey, S.; Jopson, T.; Pelz, C.; Tafessu, A.; Fareh, F.; Zuloaga, D.; Marzulla, T.; Riparip, L.-K.; Stewart, B.; Rosi, S.; et al. Short- and long-term effects of 56Fe irradiation on cognition and hippocampal DNA methylation and gene expression. BMC Genomics 2016, 17, 825. [Google Scholar] [CrossRef] [PubMed]
- Wyrobek, A.J.; Britten, R.A. Individual variations in dose response for spatial memory learning among outbred wistar rats exposed from 5 to 20 cGy of 56Fe particles. Environ. Mol. Mutagen. 2016, 57, 331–340. [Google Scholar] [CrossRef] [PubMed]
- Yan, J.; Liu, Y.; Zhao, Q.; Li, J.; Mao, A.; Li, H.; Di, C.; Zhang, H. 56Fe irradiation-induced cognitive deficits through oxidative stress in mice. Toxicol. Res. 2016, 5, 1672–1679. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Davis, L.K.; Jewell, J.S.; Miller, V.D.; Hadley, M.M.; Sanford, L.D.; Machida, M.; Lonart, G. Exposure to mission relevant doses of 1 GeV/Nucleon 56Fe particles leads to impairment of attentional set-shifting performance in socially mature rats. Radiat. Res. 2014, 182, 292–298. [Google Scholar] [CrossRef] [PubMed]
- Haley, G.E.; Yeiser, L.; Olsen, R.H.J.; Davis, M.J.; Johnson, L.A.; Raber, J. Early effects of whole-body 56Fe irradiation on hippocampal function in C57BL/6J mice. Radiat. Res. 2013, 179, 590–596. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Lau, F.C.; Cheng, V.; Luskin, K.; Carey, A.N.; Carrihill-Knoll, K.; Rabin, B.M.; Joseph, J.A. Changes in gene expression in the rat hippocampus following exposure to 56Fe particles and protection by berry diets. Cent. Nerv. Syst. Agents Med. Chem. 2013, 13, 36–42. [Google Scholar] [CrossRef] [PubMed]
- Britten, R.A.; Davis, L.K.; Johnson, A.M.; Keeney, S.; Siegel, A.; Sanford, L.D.; Singletary, S.J.; Lonart, G. Low (20 cGy) Doses of 1 GeV/u 56Fe-Particle Radiation Lead to a Persistent Reduction in the Spatial Learning Ability of Rats. Radiat. Res. 2012, 177, 146–151. [Google Scholar] [CrossRef] [PubMed]
- Lonart, G.; Parris, B.; Johnson, A.M.; Miles, S.; Sanford, L.D.; Singletary, S.J.; Britten, R.A. Executive function in rats is impaired by low (20 cGy) doses of 1 GeV/u 56Fe particles. Radiat. Res. 2012, 178, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Joseph, J.A.; Shukitt-Hale, B.; Carrihill-Knoll, K.L. Interaction between age of irradiation and age of testing in the disruption of operant performance using a ground-based model for exposure to cosmic rays. Age 2012, 34, 121–131. [Google Scholar] [CrossRef] [PubMed]
- Villasana, L.E.; Benice, T.S.; Raber, J. Long-term effects of 56Fe irradiation on spatial memory of mice: Role of sex and apolipoprotein E. isoform. Int. J. Radiat. Oncol. Biol. Phys. 2011, 80, 567–573. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.; Smith, A.; Badaut, J.; Obenaus, A. Dynamic characteristics of 56Fe-particle radiation-induced alterations in the rat brain: Magnetic resonance imaging and histological assessments. Radiat. Res. 2010, 173, 729–737. [Google Scholar] [CrossRef] [PubMed]
- Machida, M.; Lonart, G.; Britten, R.A. Low (60 cGy) Doses of 56Fe HZE-Particle Radiation Lead to a Persistent Reduction in the Glutamatergic Readily Releasable Pool in Rat Hippocampal Synaptosomes. Radiat. Res. 2010, 174, 618–623. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Carrihill-Knoll, K.; Hinchman, M.; Shukitt-Hale, B.; Joseph, J.A.; Foster, B.C. Effects of heavy particle irradiation and diet on object recognition memory in rats. Adv. Space Res. 2009, 43, 1193–1199. [Google Scholar] [CrossRef]
- Carrihill-Knoll, K.L.; Rabin, B.M.; Shukitt-Hale, B.; Joseph, J.A.; Carey, A. Amphetamine-induced taste aversion learning in young and old F-344 rats following exposure to 56Fe particles. Age 2007, 29, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Cummings, P.; Obenaus, A.; Heffron, D.; Mandell, J. High-energy (HZE) radiation exposure causes delayed axonal degeneration and astrogliosis in the central nervous system of rats. Gravitat. Space Res. 2007, 20, 89. [Google Scholar]
- Rabin, B.M.; Joseph, J.A.; Shukitt-Hale, B.; Carey, A.N. Dietary modulation of the effects of exposure to 56Fe particles. Adv. Space Res. 2007, 40, 576–580. [Google Scholar] [CrossRef]
- Rabin, B.M.; Carrihill-Knoll, K.L.; Carey, A.N.; Shukitt-Hale, B.; Joseph, J.A.; Foster, B.C. Elevated plus-maze performance of Fischer-344 rats as a function of age and of exposure to 56Fe particles. Adv. Space Res. 2007, 39, 981–986. [Google Scholar] [CrossRef]
- Shukitt-Hale, B.; Carey, A.N.; Jenkins, D.; Rabin, B.M.; Joseph, J.A. Beneficial effects of fruit extracts on neuronal function and behavior in a rodent model of accelerated aging. Neurobiol. Aging 2007, 28, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Carey, A.N.; Shukitt-Hale, B.; Rabin, B.M.; Joseph, J.A. Interaction between age and exposure to 56Fe particles on behavior and neurochemistry. Adv. Space Res. 2007, 39, 987–993. [Google Scholar] [CrossRef]
- Casadesus, G.; Shukitt-Hale, B.; Stellwagen, H.M.; Smith, M.A.; Rabin, B.M.; Joseph, J.A. Hippocampal neurogenesis and PSA-NCAM expression following exposure to 56Fe particles mimics that seen during aging in rats. Exp. Gerontol. 2005, 40, 249–254. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Joseph, J.A.; Shukitt-Hale, B. A longitudinal study of operant responding in rats irradiated when 2 months old. Radiat. Res. 2005, 164, 552–555. [Google Scholar] [CrossRef] [PubMed]
- Casadesus, G.; Shukitt-Hale, B.; Cantuti-Castelvetri, I.; Rabin, B.M.; Joseph, J.A. The effects of heavy particle irradiation on exploration and response to environmental change. Adv. Space Res. 2004, 33, 1340–1346. [Google Scholar] [CrossRef] [PubMed]
- Shukitt-Hale, B.; Casadesus, G.; Cantuti-Castelvetri, I.; Rabin, B.M.; Joseph, J.A. Cognitive deficits induced by 56Fe radiation exposure. Adv. Space Res. 2003, 31, 119–126. [Google Scholar] [CrossRef]
- Denisova, N.A.; Shukitt-Hale, B.; Rabin, B.M.; Joseph, J.A. Brain signaling and behavioral responses induced by exposure to 56Fe-particle radiation. Radiat. Res. 2002, 158, 725–734. [Google Scholar] [CrossRef]
- Rabin, B.M.; Shukitt-Hale, B.; Szprengiel, A.; Joseph, J.A. Effects of heavy particle irradiation and diet on amphetamine- and lithium chloride-induced taste avoidance learning in rats. Brain Res. 2002, 953, 31–36. [Google Scholar] [CrossRef]
- Rabin, B.M.; Hunt, W.A.; Joseph, J.A. An assessment of the behavioral toxicity of high-energy iron particles compared to other qualities of radiation. Radiat. Res. 1989, 119, 113–122. [Google Scholar] [CrossRef] [PubMed]
- Paxinos, G.; Franklin, K.B.J. The Mouse Brain in Stereotaxic Coordinates; Gulf Professional Publishing: Oxford, UK, 2004; ISBN 9780125476409. [Google Scholar]
- Eisch, A.J.; Mandyam, C.D. Adult Neurogenesis and Central Nervous System Cell Cycle Analysis. In The Cell Cycle in the Central Nervous System; Janigro, D., Ed.; Humana Press: Totowa, NJ, USA, 2006; pp. 331–358. ISBN 9781597450218. [Google Scholar]
- Zhao, C.; Deng, W.; Gage, F.H. Mechanisms and functional implications of adult neurogenesis. Cell 2008, 132, 645–660. [Google Scholar] [CrossRef] [PubMed]
- Eisch, A.J. Adult neurogenesis: Implications for psychiatry. Prog. Brain Res. 2002, 138, 315–342. [Google Scholar] [PubMed]
- Mamber, C.; Kozareva, D.A.; Kamphuis, W.; Hol, E.M. Shades of gray: The delineation of marker expression within the adult rodent subventricular zone. Prog. Neurobiol. 2013, 111, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Lim, D.A.; Alvarez-Buylla, A. The Adult Ventricular–Subventricular Zone (V-SVZ) and Olfactory Bulb (OB) Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8. [Google Scholar] [CrossRef] [PubMed]
- Whitman, M.C.; Greer, C.A. Adult neurogenesis and the olfactory system. Prog. Neurobiol. 2009, 89, 162–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mandyam, C.D.; Harburg, G.C.; Eisch, A.J. Determination of key aspects of precursor cell proliferation, cell cycle length and kinetics in the adult mouse subgranular zone. Neuroscience 2007, 146, 108–122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagace, D.C.; Donovan, M.H.; DeCarolis, N.A.; Farnbauch, L.A.; Malhotra, S.; Berton, O.; Nestler, E.J.; Krishnan, V.; Eisch, A.J. Adult hippocampal neurogenesis is functionally important for stress-induced social avoidance. Proc. Natl. Acad. Sci. USA 2010, 107, 4436–4441. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amaral, D.G.; Scharfman, H.E.; Lavenex, P. The dentate gyrus: Fundamental neuroanatomical organization (dentate gyrus for dummies). In Progress in Brain Research; Scharfman, H.E., Ed.; Elsevier: Amsterdam, The Netherlands, 2007; Volume 163, pp. 3–790. [Google Scholar]
- Kesner, R.P. A behavioral analysis of dentate gyrus function. Prog. Brain Res. 2007, 163, 567–576. [Google Scholar] [PubMed]
- Scharfman, H.E. The Dentate Gyrus: A Comprehensive Guide to Structure, Function, and Clinical Implications; Elsevier: Amsterdam, The Netherlands, 2011; ISBN 9780080551753. [Google Scholar]
- Jonas, P.; Lisman, J. Structure, function, and plasticity of hippocampal dentate gyrus microcircuits. Front. Neural Circuits 2014, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.P.; Couillard-Després, S.; Cooper-Kuhn, C.M.; Winkler, J.; Aigner, L.; Kuhn, H.G. Transient expression of doublecortin during adult neurogenesis. J. Comp. Neurol. 2003, 467, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Francis, F.; Koulakoff, A.; Boucher, D.; Chafey, P.; Schaar, B.; Vinet, M.C.; Friocourt, G.; McDonnell, N.; Reiner, O.; Kahn, A.; et al. Doublecortin is a developmentally regulated, microtubule-associated protein expressed in migrating and differentiating neurons. Neuron 1999, 23, 247–256. [Google Scholar] [CrossRef]
- Kempermann, G.; Gast, D.; Kronenberg, G.; Yamaguchi, M.; Gage, F.H. Early determination and long-term persistence of adult-generated new neurons in the hippocampus of mice. Development 2003, 130, 391–399. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, M.S.; Shetty, A.K. Efficacy of doublecortin as a marker to analyse the absolute number anddendritic growth of newly generated neurons in the adult dentate gyrus. Eur. J. Neurosci. 2004, 19, 234–246. [Google Scholar] [CrossRef] [PubMed]
- Couillard-Despres, S.; Winner, B.; Schaubeck, S.; Aigner, R.; Vroemen, M.; Weidner, N.; Bogdahn, U.; Winkler, J.; Kuhn, H.-G.; Aigner, L. Doublecortin expression levels in adult brain reflect neurogenesis. Eur. J. Neurosci. 2005, 21, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Knoth, R.; Singec, I.; Ditter, M.; Pantazis, G.; Capetian, P.; Meyer, R.P.; Horvat, V.; Volk, B.; Kempermann, G. Murine features of neurogenesis in the human hippocampus across the lifespan from 0 to 100 years. PLoS ONE 2010, 5, e8809. [Google Scholar] [CrossRef] [PubMed]
- Plümpe, T.; Ehninger, D.; Steiner, B.; Jessberger, S.; Klempin, F.; Brandt, M.; Römer, B.; Ramirez-Rodriguez, G.; Kronenberg, G.; Kempermann, G. Variability of doublecortin-associated dendrite maturation in adult hippocampal neurogenesis is independent of the regulation of precursor cell proliferation. BMC Neurosci. 2006, 7, 77. [Google Scholar] [CrossRef] [PubMed]
- Lazarov, O.; Mattson, M.P.; Peterson, D.A.; Pimplikar, S.W.; van Praag, H. When neurogenesis encounters aging and disease. Trends Neurosci. 2010, 33, 569–579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drapeau, E.; Nora Abrous, D. Stem cell review series: Role of neurogenesis in age-related memory disorders. Aging Cell 2008, 7, 569–589. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.G.; Dickinson-Anson, H.; Gage, F.H. Neurogenesis in the dentate gyrus of the adult rat: Age-related decrease of neuronal progenitor proliferation. J. Neurosci. 1996, 16, 2027–2033. [Google Scholar] [CrossRef] [PubMed]
- Kronenberg, G.; Bick-Sander, A.; Bunk, E.; Wolf, C.; Ehninger, D.; Kempermann, G. Physical exercise prevents age-related decline in precursor cell activity in the mouse dentate gyrus. Neurobiol. Aging 2006, 27, 1505–1513. [Google Scholar] [CrossRef] [PubMed]
- Seib, D.R.M.; Martin-Villalba, A. Neurogenesis in the Normal Ageing Hippocampus: A Mini-Review. Gerontology 2015, 61, 327–335. [Google Scholar] [CrossRef] [PubMed]
- Wenger, E.; Brozzoli, C.; Lindenberger, U.; Lövdén, M. Expansion and Renormalization of Human Brain Structure During Skill Acquisition. Trends Cogn. Sci. 2017, 21, 930–939. [Google Scholar] [CrossRef] [PubMed]
- Deniz, Ö.G.; Altun, G.; Kaplan, A.A.; Yurt, K.K.; von Bartheld, C.S.; Kaplan, S. A concise review of optical, physical and isotropic fractionator techniques in neuroscience studies, including recent developments. J. Neurosci. Methods 2018. [Google Scholar] [CrossRef] [PubMed]
- Guidi, S.; Ciani, E.; Severi, S.; Contestabile, A.; Bartesaghi, R. Postnatal neurogenesis in the dentate gyrus of the guinea pig. Hippocampus 2005, 15, 285–301. [Google Scholar] [CrossRef] [PubMed]
- Chumley, M.J.; Catchpole, T.; Silvany, R.E.; Kernie, S.G.; Henkemeyer, M. EphB receptors regulate stem/progenitor cell proliferation, migration, and polarity during hippocampal neurogenesis. J. Neurosci. 2007, 27, 13481–13490. [Google Scholar] [CrossRef] [PubMed]
- Naylor, A.S.; Bull, C.; Nilsson, M.K.L.; Zhu, C.; Björk-Eriksson, T.; Eriksson, P.S.; Blomgren, K.; Kuhn, H.G. Voluntary running rescues adult hippocampal neurogenesis after irradiation of the young mouse brain. Proc. Natl. Acad. Sci. USA 2008, 105, 14632–14637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pillai, A.G.; de Jong, D.; Kanatsou, S.; Krugers, H.; Knapman, A.; Heinzmann, J.-M.; Holsboer, F.; Landgraf, R.; Joëls, M.; Touma, C. Dendritic morphology of hippocampal and amygdalar neurons in adolescent mice is resilient to genetic differences in stress reactivity. PLoS ONE 2012, 7, e38971. [Google Scholar] [CrossRef] [PubMed]
- Jinno, S.; Kosaka, T. Cellular architecture of the mouse hippocampus: A quantitative aspect of chemically defined GABAergic neurons with stereology. Neurosci. Res. 2006, 56, 229–245. [Google Scholar] [CrossRef] [PubMed]
- Kuhn, H.G.; Eisch, A.J.; Spalding, K.; Peterson, D.A. Detection and Phenotypic Characterization of Adult Neurogenesis. Cold Spring Harb. Perspect. Biol. 2016, 8, a025981. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Poppenk, J.; Evensmoen, H.R.; Moscovitch, M.; Nadel, L. Long-axis specialization of the human hippocampus. Trends Cogn. Sci. 2013, 17, 230–240. [Google Scholar] [CrossRef] [PubMed]
- Witter, M.P. Organization of the entorhinal—Hippocampal system: A review of current anatomical data. Hippocampus 1993, 3, 33–44. [Google Scholar] [PubMed]
- Amaral, D.G.; Witter, M.P. The three-dimensional organization of the hippocampal formation: A review of anatomical data. Neuroscience 1989, 31, 571–591. [Google Scholar] [CrossRef]
- Jarrard, L.E.; Luu, L.P.; Davidson, T.L. A study of hippocampal structure-function relations along the septo-temporal axis. Hippocampus 2012, 22, 680–692. [Google Scholar] [CrossRef] [PubMed]
- Grivas, I.; Michaloudi, H.; Batzios, C.; Chiotelli, M.; Papatheodoropoulos, C.; Kostopoulos, G.; Papadopoulos, G.C. Vascular network of the rat hippocampus is not homogeneous along the septotemporal axis. Brain Res. 2003, 971, 245–249. [Google Scholar] [CrossRef]
- Kheirbek, M.A.; Hen, R. Dorsal vs ventral hippocampal neurogenesis: Implications for cognition and mood. Neuropsychopharmacology 2011, 36, 373–374. [Google Scholar] [CrossRef] [PubMed]
- Jinno, S. Topographic differences in adult neurogenesis in the mouse hippocampus: A stereology-based study using endogenous markers. Hippocampus 2011, 21, 467–480. [Google Scholar] [CrossRef] [PubMed]
- Wu, M.V.; Hen, R. Functional dissociation of adult-born neurons along the dorsoventral axis of the dentate gyrus. Hippocampus 2014, 24, 751–761. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiget, F.; van Dijk, R.M.; Louet, E.R.; Slomianka, L.; Amrein, I. Effects of Strain and Species on the Septo-Temporal Distribution of Adult Neurogenesis in Rodents. Front. Neurosci. 2017, 11, 719. [Google Scholar] [CrossRef] [PubMed]
- Schreurs, A.; Sabanov, V.; Balschun, D. Distinct Properties of Long-Term Potentiation in the Dentate Gyrus along the Dorsoventral Axis: Influence of Age and Inhibition. Sci. Rep. 2017, 7, 5157. [Google Scholar] [CrossRef] [PubMed]
- Bekiari, C.; Grivas, I.; Giannakopoulou, A.; Michaloudi-Pavlou, H.; Kostopoulos, G.K.; Papadopoulos, G.C. Dentate gyrus variation along its septo-temporal axis: Structure and function in health and disease. In Dentate Gyrus: Structure, Role in Disease, and Potential Health Implications; Nova Science Publishers, Inc.: Hauppauge, NY, USA, 2015. [Google Scholar]
- Seki, T.; Namba, T.; Mochizuki, H.; Onodera, M. Clustering, migration, and neurite formation of neural precursor cells in the adult rat hippocampus. J. Comp. Neurol. 2007, 502, 275–290. [Google Scholar] [CrossRef] [PubMed]
- Rabin, B.M.; Shukitt-Hale, B. A voyage to Mars: Space radiation, aging, and nutrition. Nutr. Aging 2014, 2, 233–241. [Google Scholar]
- Sridharan, D.M.; Asaithamby, A.; Bailey, S.M.; Costes, S.V.; Doetsch, P.W.; Dynan, W.S.; Kronenberg, A.; Rithidech, K.N.; Saha, J.; Snijders, A.M.; et al. Understanding cancer development processes after HZE-particle exposure: Roles of ROS, DNA damage repair and inflammation. Radiat. Res. 2015, 183, 1–26. [Google Scholar] [CrossRef] [PubMed]
- Poulose, S.M.; Rabin, B.M.; Bielinski, D.F.; Kelly, M.E.; Miller, M.G.; Thanthaeng, N.; Shukitt-Hale, B. Neurochemical differences in learning and memory paradigms among rats supplemented with anthocyanin-rich blueberry diets and exposed to acute doses of 56Fe particles. Life Sci. Space Res. 2017, 12, 16–23. [Google Scholar] [CrossRef] [PubMed]
- Goodwin, T.J.; Christofidou-Solomidou, M. Oxidative Stress and Space Biology: An Organ-Based Approach. Int. J. Mol. Sci. 2018, 19, 959. [Google Scholar] [CrossRef] [PubMed]
- Reagan, L.P.; Magariños, A.M.; Yee, D.K.; Swzeda, L.I.; Van Bueren, A.; McCall, A.L.; McEwen, B.S. Oxidative stress and HNE conjugation of GLUT3 are increased in the hippocampus of diabetic rats subjected to stress. Brain Res. 2000, 862, 292–300. [Google Scholar] [CrossRef]
- Resende, R.; Moreira, P.I.; Proença, T.; Deshpande, A.; Busciglio, J.; Pereira, C.; Oliveira, C.R. Brain oxidative stress in a triple-transgenic mouse model of Alzheimer disease. Free Radic. Biol. Med. 2008, 44, 2051–2057. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shang, J.; Yan, H.; Jiao, Y.; Ohta, Y.; Liu, X.; Li, X.; Morihara, R.; Nakano, Y.; Fukui, Y.; Shi, X.; et al. Therapeutic Effects of Pretreatment with Tocovid on Oxidative Stress in Postischemic Mice Brain. J. Stroke Cerebrovasc. Dis. 2018, 27, 2096–2105. [Google Scholar] [CrossRef] [PubMed]
- Folbergrová, J.; Ješina, P.; Kubová, H.; Otáhal, J. Effect of Resveratrol on Oxidative Stress and Mitochondrial Dysfunction in Immature Brain during Epileptogenesis. Mol. Neurobiol. 2018, 55, 7512–7522. [Google Scholar] [CrossRef] [PubMed]
- Canever, L.; Freire, T.G.; Mastella, G.A.; Damázio, L.; Gomes, S.; Fachim, I.; Michels, C.; Carvalho, G.; Godói, A.K.; Peterle, B.R.; et al. Changes in behavioural parameters, oxidative stress and neurotrophins in the brain of adult offspring induced to an animal model of schizophrenia: The effects of FA deficient or FA supplemented diet during the neurodevelopmental phase. Prog. Neuropsychopharmacol. Biol. Psychiatry 2018, 86, 52–64. [Google Scholar] [CrossRef] [PubMed]
- Dal-Pont, G.C.; Resende, W.R.; Varela, R.B.; Menegas, S.; Trajano, K.S.; Peterle, B.R.; Quevedo, J.; Valvassori, S.S. Inhibition of GSK-3β on Behavioral Changes and Oxidative Stress in an Animal Model of Mania. Mol. Neurobiol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Boström, M.; Ek, C.J.; Li, T.; Xie, C.; Xu, Y.; Sun, Y.; Blomgren, K.; Zhu, C. Radiation induces progenitor cell death, microglia activation, and blood-brain barrier damage in the juvenile rat cerebellum. Sci. Rep. 2017, 7, 46181. [Google Scholar] [CrossRef] [PubMed]
- Lei, R.; Zhao, T.; Li, Q.; Wang, X.; Ma, H.; Deng, Y. Carbon Ion Irradiated Neural Injury Induced the Peripheral Immune Effects in Vitro or in Vivo. Int. J. Mol. Sci. 2015, 16, 28334–28346. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kubo, N.; Noda, S.-E.; Takahashi, A.; Yoshida, Y.; Oike, T.; Murata, K.; Musha, A.; Suzuki, Y.; Ohno, T.; Takahashi, T.; et al. Radiosensitizing effect of carboplatin and paclitaxel to carbon-ion beam irradiation in the non-small-cell lung cancer cell line H460. J. Radiat. Res. 2015, 56, 229–238. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kanai, T.; Furusawa, Y.; Fukutsu, K.; Itsukaichi, H.; Eguchi-Kasai, K.; Ohara, H. Irradiation of mixed beam and design of spread-out Bragg peak for heavy-ion radiotherapy. Radiat. Res. 1997, 147, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Mizoe, J.-E. Review of carbon ion radiotherapy for skull base tumors (especially chordomas). Rep. Pract. Oncol. Radiother. 2016, 21, 356–360. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagace, D.C.; Whitman, M.C.; Noonan, M.A.; Ables, J.L.; DeCarolis, N.A.; Arguello, A.A.; Donovan, M.H.; Fischer, S.J.; Farnbauch, L.A.; Beech, R.D.; et al. Dynamic contribution of nestin-expressing stem cells to adult neurogenesis. J. Neurosci. 2007, 27, 12623–12629. [Google Scholar] [CrossRef] [PubMed]
- Kempermann, G.; Jessberger, S.; Steiner, B.; Kronenberg, G. Milestones of neuronal development in the adult hippocampus. Trends Neurosci. 2004, 27, 447–452. [Google Scholar] [CrossRef] [PubMed]
- DeCarolis, N.A.; Mechanic, M.; Petrik, D.; Carlton, A.; Ables, J.L.; Malhotra, S.; Bachoo, R.; Gotz, M.; Lagace, D.C.; Eisch, A.J. In vivo contribution of nestin- and GLAST-lineage cells to adult hippocampal neurogenesis. Hippocampus 2013, 23, 708–719. [Google Scholar] [CrossRef] [PubMed]
- BNL|NASA Space Radiation Laboratory (NSRL). Available online: http://www.bnl.gov/medical/NASA/NSRL_description.asp (accessed on 9 March 2017).
- Walker, A.K.; Rivera, P.D.; Wang, Q.; Chuang, J.-C.; Tran, S.; Osborne-Lawrence, S.; Estill, S.J.; Starwalt, R.; Huntington, P.; Morlock, L.; et al. The P7C3 class of neuroprotective compounds exerts antidepressant efficacy in mice by increasing hippocampal neurogenesis. Mol. Psychiatry 2015, 20, 500–508. [Google Scholar] [CrossRef] [PubMed]
- Yun, S.; Donovan, M.H.; Ross, M.N.; Richardson, D.R.; Reister, R.; Farnbauch, L.A.; Fischer, S.J.; Riethmacher, D.; Gershenfeld, H.K.; Lagace, D.C.; et al. Stress-Induced Anxiety- and Depressive-Like Phenotype Associated with Transient Reduction in Neurogenesis in Adult Nestin-CreERT2/Diphtheria Toxin Fragment, A. Transgenic Mice. PLoS ONE 2016, 11, e0147256. [Google Scholar] [CrossRef] [PubMed]
- Howard, V.; Reed, M. Unbiased Stereology: Three-Dimensional Measurement in Microscopy; Garland Science: New York, NY, USA, 2004; ISBN 9781135331672. [Google Scholar]
- Mouton, P. Applications of Modern Stereology to Quantitative Light Microscopy. Microsc. Microanal. 2009, 15, 1526–1527. [Google Scholar] [CrossRef] [Green Version]
- West, M.J. Basic Stereology for Biologists and Neuroscientists; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, NY, USA, 2012. [Google Scholar]
- Boyce, R.W.; Dorph-Petersen, K.-A.; Lyck, L.; Gundersen, H.J.G. Design-based stereology: Introduction to basic concepts and practical approaches for estimation of cell number. Toxicol. Pathol. 2010, 38, 1011–1025. [Google Scholar] [CrossRef] [PubMed]
- Gundersen, H.J.; Bagger, P.; Bendtsen, T.F.; Evans, S.M.; Korbo, L.; Marcussen, N.; Møller, A.; Nielsen, K.; Nyengaard, J.R.; Pakkenberg, B. The new stereological tools: Disector, fractionator, nucleator and point sampled intercepts and their use in pathological research and diagnosis. APMIS 1988, 96, 857–881. [Google Scholar] [CrossRef] [PubMed]
- Schmitz, C.; Hof, P.R. Design-based stereology in neuroscience. Neuroscience 2005, 130, 813–831. [Google Scholar] [CrossRef] [PubMed]
- Basler, L.; Gerdes, S.; Wolfer, D.P.; Slomianka, L. Sampling the Mouse Hippocampal Dentate Gyrus. Front. Neuroanat. 2017, 11. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eisch, A.J.; Barrot, M.; Schad, C.A.; Self, D.W.; Nestler, E.J. Opiates inhibit neurogenesis in the adult rat hippocampus. Proc. Natl. Acad. Sci. USA 2000, 97, 7579–7584. [Google Scholar] [CrossRef] [PubMed] [Green Version]









© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zanni, G.; Deutsch, H.M.; Rivera, P.D.; Shih, H.-Y.; LeBlanc, J.A.; Amaral, W.Z.; Lucero, M.J.; Redfield, R.L.; DeSalle, M.J.; Chen, B.P.C.; et al. Whole-Body 12C Irradiation Transiently Decreases Mouse Hippocampal Dentate Gyrus Proliferation and Immature Neuron Number, but Does Not Change New Neuron Survival Rate. Int. J. Mol. Sci. 2018, 19, 3078. https://doi.org/10.3390/ijms19103078
Zanni G, Deutsch HM, Rivera PD, Shih H-Y, LeBlanc JA, Amaral WZ, Lucero MJ, Redfield RL, DeSalle MJ, Chen BPC, et al. Whole-Body 12C Irradiation Transiently Decreases Mouse Hippocampal Dentate Gyrus Proliferation and Immature Neuron Number, but Does Not Change New Neuron Survival Rate. International Journal of Molecular Sciences. 2018; 19(10):3078. https://doi.org/10.3390/ijms19103078
Chicago/Turabian StyleZanni, Giulia, Hannah M. Deutsch, Phillip D. Rivera, Hung-Ying Shih, Junie A. LeBlanc, Wellington Z. Amaral, Melanie J. Lucero, Rachel L. Redfield, Matthew J. DeSalle, Benjamin P. C. Chen, and et al. 2018. "Whole-Body 12C Irradiation Transiently Decreases Mouse Hippocampal Dentate Gyrus Proliferation and Immature Neuron Number, but Does Not Change New Neuron Survival Rate" International Journal of Molecular Sciences 19, no. 10: 3078. https://doi.org/10.3390/ijms19103078