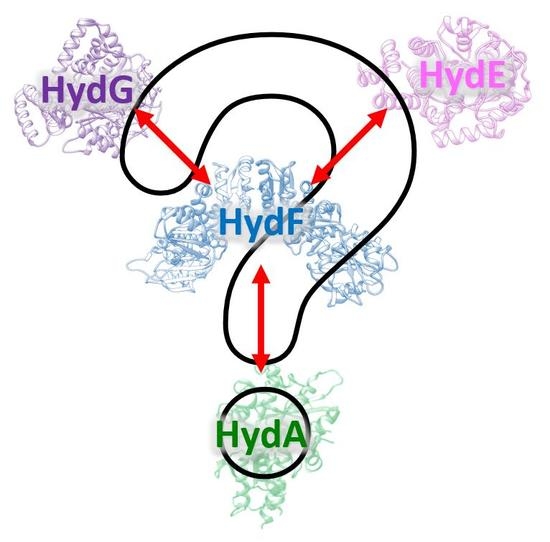
Overview of the Maturation Machinery of the H-Cluster of [FeFe]-Hydrogenases with a Focus on HydF


Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. The maturases
2.1. HydE
2.2. HydG
2.3. HydF
2.3.1. GTPase Domain
2.3.2. Dimerization Domain
2.3.3. Cluster Binding Domain
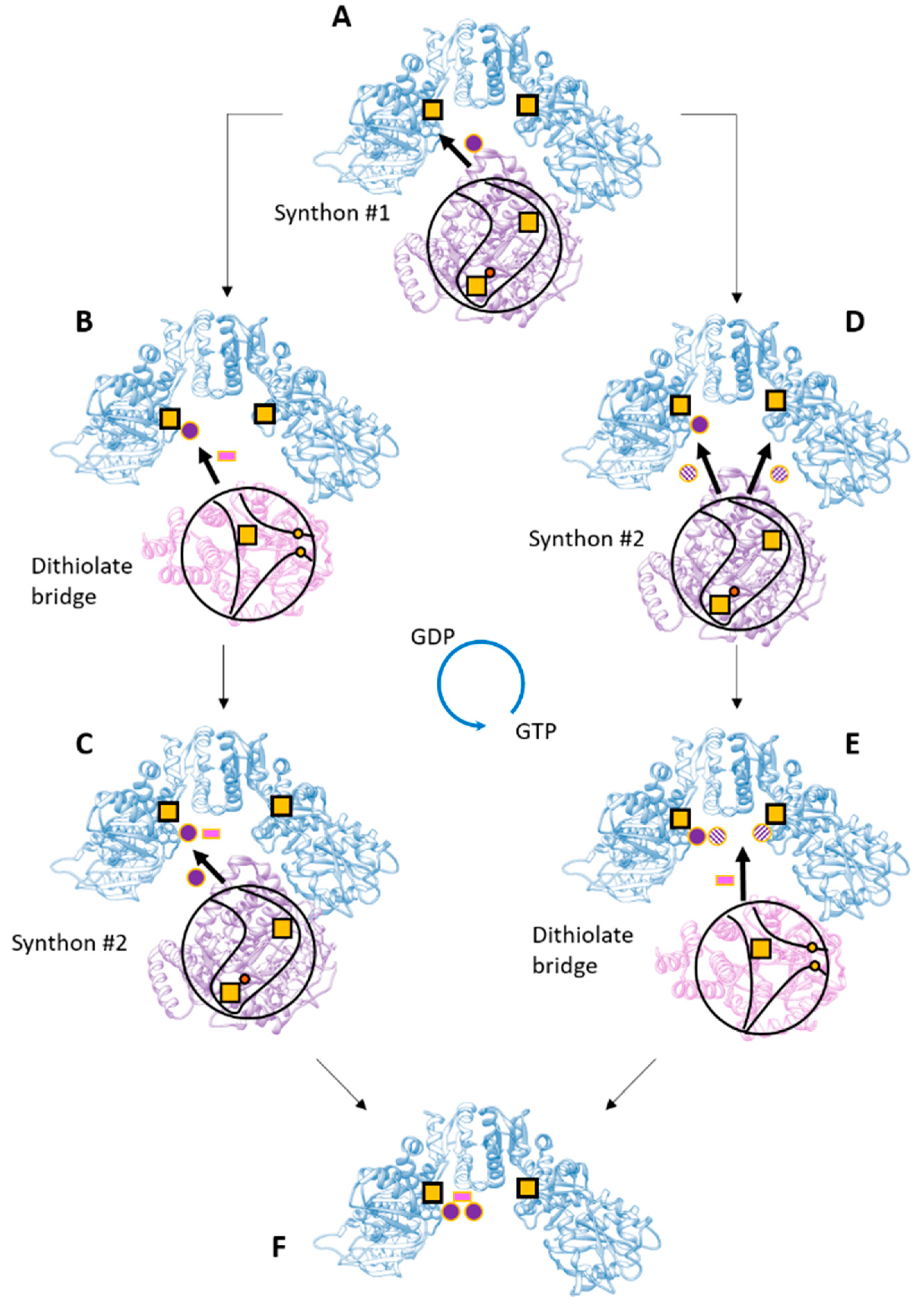
2.3.4. Tetrameric Form
2.3.5. [2Fe]H Cluster Precursor
2.3.6. Role of the GTPase Domain
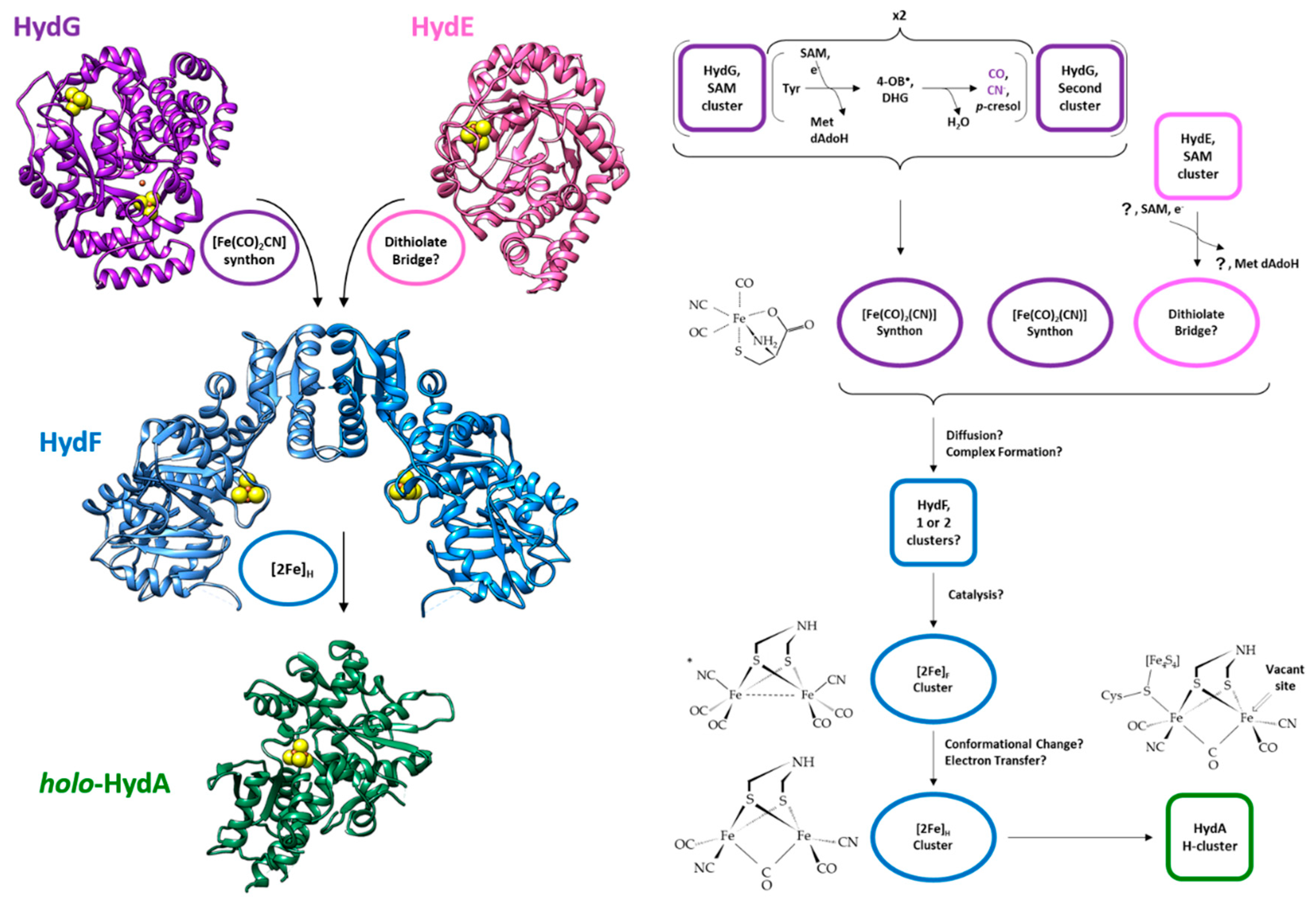
3. The Overall Process
Funding
Conflicts of Interest
Abbreviations
| EPR | Electron Paramagnetic Resonance |
| GTP | Guanosine-5′-Triphosphate |
| GDP | Guanosine-5′-Diphosphate |
| SAM | S-adenosyl-L-Methionine |
| TIM | Triose-Phosphate Isomerase |
| DHG | Dehydroglycine |
| dAdoH | 5′- Deoxyadenosine |
| 4OB• | 4-Oxidobenzyl Radical |
References
- McPherson, I.J.; Vincent, K.A. Electrocatalysis by Hydrogenases: Lessons for Building Bio-Inspired Devices. J. Braz. Chem. Soc. 2014, 25, 1–15. [Google Scholar] [CrossRef]
- Morra, S.; Valetti, F.; Gilardi, G. [FeFe]-hydrogenases as biocatalysts in bio-hydrogen production. Rend. Lincei 2017, 28, 183–194. [Google Scholar] [CrossRef]
- Khan, M.A.; Ngo, H.H.; Guo, W.; Liu, Y.; Zhang, X.; Guo, J.; Chang, S.W.; Nguyen, D.D.; Wang, J. Biohydrogen production from anaerobic digestion and its potential as renewable energy. Renew. Energy 2018, 129, 754–768. [Google Scholar] [CrossRef]
- Reeve, H.A.; Ash, P.A.; Park, H.; Huang, A.; Posidias, M.; Tomlinson, C.; Lenz, O.; Vincent, K.A. Enzymes as modular catalysts for redox half-reactions in H2-powered chemical synthesis: From biology to technology. Biochem. J. 2017, 474, 215–230. [Google Scholar] [CrossRef] [PubMed]
- Esmieu, C.; Raleiras, P.; Berggren, G. From protein engineering to artificial enzymes—Biological and biomimetic approaches towards sustainable hydrogen production. Sustain. Energy Fuels 2017, 724–750. [Google Scholar] [CrossRef]
- Greening, C.; Biswas, A.; Carere, C.R.; Jackson, C.J.; Taylor, M.C.; Stott, M.B.; Cook, G.M.; Morales, S.E. Genomic and metagenomic surveys of hydrogenase distribution indicate H2 is a widely utilised energy source for microbial growth and survival. ISME J. 2016, 10, 761–777. [Google Scholar] [CrossRef] [PubMed]
- Vignais, P.M.; Billoud, B. Occurrence, classification, and biological function of hydrogenases: An overview. Chem. Rev. 2007, 107, 4206–4272. [Google Scholar] [CrossRef] [PubMed]
- Lubitz, W.; Ogata, H.; Rüdiger, O.; Reijerse, E. Hydrogenases. Chem. Rev. 2014, 114, 4081–4148. [Google Scholar] [CrossRef] [PubMed]
- Corr, M.J.; Murphy, J.A. Evolution in the understanding of [Fe]-hydrogenase. Chem. Soc. Rev. 2011, 40, 2279–2292. [Google Scholar] [CrossRef] [PubMed]
- Ogata, H.; Lubitz, W.; Higuchi, Y. Structure and function of [NiFe] hydrogenases. J. Biochem. 2016, 160, 251–258. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mulder, D.W.; Shepard, E.M.; Meuser, J.E.; Joshi, N.; King, P.W.; Posewitz, M.C.; Broderick, J.B.; Peters, J.W. Insights into [FeFe]-hydrogenase structure, mechanism, and maturation. Structure 2011, 19, 1038–1052. [Google Scholar] [CrossRef] [PubMed]
- Baltazar, C.S.A.; Marques, M.C.; Soares, C.M.; DeLacey, A.M.; Pereira, I.A.C.; Matias, P.M. Nickel-iron-selenium hydrogenases—An overview. Eur. J. Inorg. Chem. 2011, 948–962. [Google Scholar] [CrossRef] [Green Version]
- Vincent, K.A.; Parkin, A.; Armstrong, F.A. Investigating and exploiting the electrocatalytic properties of hydrogenases. Chem. Rev. 2007, 107, 4366–4413. [Google Scholar] [CrossRef] [PubMed]
- Marques, M.C.; Coelho, R.; De Lacey, A.L.; Pereira, I.A.C.; Matias, P.M. The three-dimensional structure of [nifese] hydrogenase from desulfovibrio vulgaris hildenborough: A hydrogenase without a bridging ligand in the active site in its oxidised, “as-isolated” state. J. Mol. Biol. 2010, 396, 893–907. [Google Scholar] [CrossRef] [PubMed]
- De Lacey, A.L.; Fernández, V.M.; Rousset, M.; Cammack, R. Activation and inactivation of hydrogenase function and the catalytic cycle: Spectroelectrochemical studies. Chem. Rev. 2007, 107, 4304–4330. [Google Scholar] [CrossRef] [PubMed]
- Goldet, G.; Brandmayr, C.; Stripp, S.T.; Happe, T.; Cavazza, C.; Fontecilla-Camps, J.C.; Armstrong, F.A. Electrochemical kinetic investigations of the reactions of [FeFe]-hydrogenases with carbon monoxide and oxygen: Comparing the importance of gas tunnels and active-site electronic/redox effects. J. Am. Chem. Soc. 2009, 131, 14979–14989. [Google Scholar] [CrossRef] [PubMed]
- Stripp, S.T.; Goldet, G.; Brandmayr, C.; Sanganas, O.; Vincent, K.A.; Haumann, M.; Armstrong, F.A.; Happe, T. How oxygen attacks [FeFe] hydrogenases from photosynthetic organisms. Proc. Natl. Acad. Sci. USA 2009, 106, 17331–17336. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lambertz, C.; Leidel, N.; Havelius, K.G.V.; Noth, J.; Chernev, P.; Winkler, M.; Happe, T.; Haumann, M. O2 reactions at the six-iron active site (H-cluster) in [FeFe]-hydrogenase. J. Biol. Chem. 2011, 286, 40614–40623. [Google Scholar] [CrossRef] [PubMed]
- Rodríguez-Maciá, P.; Reijerse, E.J.; Van Gastel, M.; Debeer, S.; Lubitz, W.; Rüdiger, O.; Birrell, J.A. Sulfide Protects [FeFe] Hydrogenases from O2. J. Am. Chem. Soc. 2018, 140, 9346–9350. [Google Scholar] [CrossRef] [PubMed]
- Oughli, A.A.; Vélez, M.; Birrell, J.A.; Schuhmann, W.; Lubitz, W.; Plumeré, N.; Rüdiger, O. Viologen-modified electrodes for protection of hydrogenases from high potential inactivation while performing H2oxidation at low overpotential. Dalt. Trans. 2018, 47, 10685–10691. [Google Scholar] [CrossRef] [PubMed]
- Caserta, G.; Papini, C.; Adamska-Venkatesh, A.; Pecqueur, L.; Sommer, C.; Reijerse, E.; Lubitz, W.; Gauquelin, C.; Meynial-Salles, I.; Pramanik, D.; et al. Engineering an [FeFe]-Hydrogenase: Do Accessory Clusters Influence O2 Resistance and Catalytic Bias? J. Am. Chem. Soc. 2018, 140, 5516–5526. [Google Scholar] [CrossRef] [PubMed]
- Apfel, U.P.; Weigand, W. Biomimetic assembly of the [FeFe] hydrogenase: Synthetic mimics in a biological shell. ChemBioChem 2013, 14, 2237–2238. [Google Scholar] [CrossRef] [PubMed]
- Artero, V.; Berggren, G.; Atta, M.; Caserta, G.; Roy, S.; Pecqueur, L.; Fontecave, M. From Enzyme Maturation to Synthetic Chemistry: The Case of Hydrogenases. Acc. Chem. Res. 2015, 48, 2380–2387. [Google Scholar] [CrossRef] [PubMed]
- Birrell, J.A.; Rüdiger, O.; Reijerse, E.J.; Lubitz, W. Semisynthetic Hydrogenases Propel Biological Energy Research into a New Era. Joule 2017, 1, 61–76. [Google Scholar] [CrossRef]
- Stripp, S.T.; Happe, T. How algae produce hydrogen—News from the photosynthetic hydrogenase. Dalt. Trans. 2009, 45, 9960–9969. [Google Scholar] [CrossRef] [PubMed]
- Haumann, M.; Stripp, S.T. The Molecular Proceedings of Biological Hydrogen Turnover. Acc. Chem. Res. 2018, 51, 1755–1763. [Google Scholar] [CrossRef] [PubMed]
- Schilter, D.; Rauchfuss, T.B. And the winner is...azadithiolate: An amine proton relay in the [FeFe] hydrogenases. Angew. Chem. Int. Ed. 2013, 52, 13518–13520. [Google Scholar] [CrossRef] [PubMed]
- Adamska-Venkatesh, A.; Roy, S.; Siebel, J.F.; Simmons, T.R.; Fontecave, M.; Artero, V.; Reijerse, E.; Lubitz, W. Spectroscopic Characterization of the Bridging Amine in the Active Site of [FeFe] Hydrogenase Using Isotopologues of the H.-Cluster. J. Am. Chem. Soc. 2015, 137, 12744–12747. [Google Scholar] [CrossRef] [PubMed]
- Lill, R. Function and biogenesis of iron-sulphur proteins. Nature 2009, 460, 831–838. [Google Scholar] [CrossRef] [PubMed]
- Posewitz, M.C.; King, P.W.; Smolinski, S.L.; Zhang, L.; Seibert, M.; Ghirardi, M.L. Discovery of two novel radical S-adenosylmethionine proteins required for the assembly of an active [Fe] hydrogenase. J. Biol. Chem. 2004, 279, 25711–25720. [Google Scholar] [CrossRef] [PubMed]
- Mulder, D.W.; Ortillo, D.O.; Gardenghi, D.J.; Naumov, A.V.; Ruebush, S.S.; Szilagyi, R.K.; Huynh, B.; Broderick, J.B.; Peters, J.W. Activation of HydAΔEFG requires a preformed [4Fe-4S] cluster. Biochemistry 2009, 48, 6240–6248. [Google Scholar] [CrossRef] [PubMed]
- Nicolet, Y.; Fontecilla-Camps, J.C. Structure-function relationships in [FeFe]-hydrogenase active site maturation. J. Biol. Chem. 2012, 287, 13532–13540. [Google Scholar] [CrossRef] [PubMed]
- Peters, J.W.; Schut, G.J.; Boyd, E.S.; Mulder, D.W.; Shepard, E.M.; Broderick, J.B.; King, P.W.; Adams, M.W.W. [FeFe]- and [NiFe]-hydrogenase diversity, mechanism, and maturation. Biochim. Biophys. Acta-Mol. Cell. Res. 2015, 1853, 1350–1369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, P.W.; Posewitz, M.C.; Ghirardi, M.L.; Seibert, M.; Al, K.E.T.; Acteriol, J.B. Functional Studies of [FeFe] Hydrogenase Maturation in an Escherichia coli Biosynthetic System Functional Studies of [FeFe] Hydrogenase Maturation in an Escherichia coli Biosynthetic System. J. Bacteriol. 2006, 188, 2163–2172. [Google Scholar] [CrossRef] [PubMed]
- Bourne, H.R.; Sanders, D.A.; McCormick, F. The GTPase superfamily: Conserved structure and molecular mechanism. Nature 1991, 349, 117–127. [Google Scholar] [CrossRef] [PubMed]
- McGlynn, S.E.; Shepard, E.M.; Winslow, M.A.; Naumov, A.V.; Duschene, K.S.; Posewitz, M.C.; Broderick, W.E.; Broderick, J.B.; Peters, J.W. HydF as a scaffold protein in [FeFe] hydrogenase H-cluster biosynthesis. FEBS Lett. 2008, 582, 2183–2187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Czech, I.; Silakov, A.; Lubitz, W.; Happe, T. The [FeFe]-hydrogenase maturase HydF from Clostridium acetobutylicum contains a CO and CN-ligated iron cofactor. FEBS Lett. 2010, 584, 638–642. [Google Scholar] [CrossRef] [PubMed]
- Betz, J.N.; Boswell, N.W.; Fugate, C.J.; Holliday, G.L.; Akiva, E.; Scott, A.G.; Babbitt, P.C.; Peters, J.W.; Shepard, E.M.; Broderick, J.B. Hydrogenase maturation: Insights into the role hyde plays in dithiomethylamine biosynthesis. Biochemistry 2015, 54, 1807–1818. [Google Scholar] [CrossRef] [PubMed]
- Nicolet, Y.; Rubach, J.K.; Posewitz, M.C.; Amara, P.; Mathevon, C.; Atta, M.; Fontecave, M.; Fontecilla-Camps, J.C. X-ray structure of the [FeFe]-hydrogenase maturase HydE from Thermotoga maritima. J. Biol. Chem. 2008, 283, 18861–18872. [Google Scholar] [CrossRef] [PubMed]
- Nicolet, Y.; Rohac, R.; Martin, L.; Fontecilla-Camps, J.C. X-ray snapshots of possible intermediates in the time course of synthesis and degradation of protein-bound Fe4S4 clusters. Proc. Natl. Acad. Sci. USA 2013, 110, 7188–7192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rao, G.; Tao, L.; Suess, D.L.M.; Britt, R.D. A [4Fe–4S]-Fe(CO)(CN)-l-cysteine intermediate is the first organometallic precursor in [FeFe] hydrogenase H-cluster bioassembly. Nat. Chem. 2018, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Nicolet, Y.; Pagnier, A.; Zeppieri, L.; Martin, L.; Amara, P.; Fontecilla-Camps, J.C. Crystal structure of HydG from carboxydothermus hydrogenoformans: A trifunctional [FeFe]-Hydrogenase maturase. Chem. Biol. Chem. 2015, 16, 397–402. [Google Scholar] [CrossRef] [PubMed]
- Dinis, P.; Suess, D.L.M.; Fox, S.J.; Harmer, J.E.; Driesener, R.C.; De La Paz, L.; Swartz, J.R.; Essex, J.W.; Britt, R.D.; Roach, P.L. X-ray crystallographic and EPR spectroscopic analysis of HydG, a maturase in [FeFe]-hydrogenase H.-cluster assembly. Proc. Natl. Acad. Sci. USA 2015, 112, 1362–1367. [Google Scholar] [CrossRef] [PubMed]
- Kuchenreuther, J.M.; Myers, W.K.; Stich, T.A.; George, S.J.; Nejatyjahromy, Y.; Swartz, J.R.; Britt, R.D. A Radical Intermediate in Tyrosine of FeFe Hydrogenase. Science 2013, 342, 472–475. [Google Scholar] [CrossRef] [PubMed]
- Kuchenreuther, J.M. The HydG Enzyme Generates an. Science 2014, 343, 424. [Google Scholar] [CrossRef] [PubMed]
- Suess, D.L.M.; Kuchenreuther, J.M.; De La Paz, L.; Swartz, J.R.; Britt, R.D. Biosynthesis of the [FeFe] Hydrogenase H Cluster: A Central Role for the Radical SAM Enzyme HydG. Inorg. Chem. 2016, 55, 478–487. [Google Scholar] [CrossRef] [PubMed]
- Suess, D.L.M.; Pham, C.C.; Bürstel, I.; Swartz, J.R.; Cramer, S.P.; Britt, R.D. The Radical SAM Enzyme HydG Requires Cysteine and a Dangler Iron for Generating an Organometallic Precursor to the [FeFe]-Hydrogenase, H.-Cluster. J. Am. Chem. Soc. 2016, 138, 1146–1149. [Google Scholar] [CrossRef] [PubMed]
- Cendron, L.; Berto, P.; D’Adamo, S.; Vallese, F.; Govoni, C.; Posewitz, M.C.; Giacometti, G.M.; Costantini, P.; Zanotti, G. Crystal structure of HydF scaffold protein provides insights into [FeFe]-hydrogenase maturation. J. Biol. Chem. 2011, 286, 43944–43950. [Google Scholar] [CrossRef] [PubMed]
- Shepard, E.M.; Duffus, B.R.; George, S.J.; Mcglynn, S.E.; Challand, M.R.; Swanson, K.D.; Roach, P.L.; Cramer, S.P.; Peters, J.W.; Broderick, J.B. [FeFe]-Hydrogenase Maturation: HydG-Catalyzed Synthesis of Carbon Monoxide. J. Am. Chem. Soc. 2010, 132, 9247–9249. [Google Scholar] [CrossRef] [PubMed]
- Brazzolotto, X.; Rubach, J.K.; Gaillard, J.; Gambarelli, S.; Atta, M.; Fontecave, M. The [Fe-Fe]-hydrogenase maturation protein HydF from Thermotoga maritima is a GTPase with an iron-sulfur cluster. J. Biol. Chem. 2006, 281, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Czech, I.; Stripp, S.; Sanganas, O.; Leidel, N.; Happe, T.; Haumann, M. The [FeFe]-hydrogenase maturation protein HydF contains a H-cluster like [4Fe4S]-2Fe site. FEBS Lett. 2011, 585, 225–230. [Google Scholar] [CrossRef] [PubMed]
- Berto, P.; Di Valentin, M.; Cendron, L.; Vallese, F.; Albertini, M.; Salvadori, E.; Giacometti, G.M.; Carbonera, D.; Costantini, P. The [4Fe-4S]-cluster coordination of [FeFe]-hydrogenase maturation protein HydF as revealed by EPR and HYSCORE spectroscopies. Biochim. Biophys. Acta-Bioenergy 2012, 1817, 2149–2157. [Google Scholar] [CrossRef] [PubMed]
- Berggren, G.; Garcia-Serres, R.; Brazzolotto, X.; Clemancey, M.; Gambarelli, S.; Atta, M.; Latour, J.M.; Hernández, H.L.; Subramanian, S.; Johnson, M.K.; et al. An EPR/HYSCORE, Mössbauer, and resonance Raman study of the hydrogenase maturation enzyme HydF: A model for N-coordination to [4Fe-4S] clusters. J. Biol. Inorg. Chem. 2014, 19, 75–84. [Google Scholar] [CrossRef] [PubMed]
- Albertini, M.; Vallese, F.; Di Valentin, M.; Berto, P.; Giacometti, G.M.; Costantini, P.; Carbonera, D. The proton iron-sulfur cluster environment of the [FeFe]-hydrogenase maturation protein HydF from Thermotoga neapolitana. Int. J. Hydrogen Energy 2014, 39, 18574–18582. [Google Scholar] [CrossRef]
- Albertini, M.; Galazzo, L.; Maso, L.; Vallese, F.; Berto, P.; De Rosa, E.; Di Valentin, M.; Costantini, P.; Carbonera, D. Characterization of the [FeFe]-Hydrogenase Maturation Protein HydF by EPR Techniques: Insights into the Catalytic Mechanism. Top. Catal. 2015, 58, 708–718. [Google Scholar] [CrossRef]
- Caserta, G.; Pecqueur, L.; Adamska-Venkatesh, A.; Papini, C.; Roy, S.; Artero, V.; Atta, M.; Reijerse, E.; Lubitz, W.; Fontecave, M. Structural and functional characterization of the hydrogenase-maturation HydF protein. Nat. Chem. Biol. 2017, 13, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Galazzo, L.; Maso, L.; De Rosa, E.; Bortolus, M.; Doni, D.; Acquasaliente, L.; Filippis, V.D.V.D.; Costantini, P.; Carbonera, D. Identifying conformational changes with site-directed spin labeling reveals that the GTPase domain of HydF is a molecular switch. Sci. Rep. 2017, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ash, M.R.; Maher, M.J.; Mitchell Guss, J.; Jormakka, M. The cation-dependent G-proteins: In a class of their own. FEBS Lett. 2012, 586, 2218–2224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera - A visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed]
- Ash, M.R.; Guilfoyle, A.; Clarke, R.J.; Guss, J.M.; Maher, M.J.; Jormakka, M. Potassium-activated GTPase reaction in the G protein-coupled ferrous iron transporter B. J. Biol. Chem. 2010, 285, 14594–14602. [Google Scholar] [CrossRef] [PubMed]
- Broderick, J.B.; Byer, A.S.; Duschene, K.S.; Duffus, B.R.; Betz, J.N.; Shepard, E.M.; Peters, J.W. H-Cluster assembly during maturation of the [FeFe]-hydrogenase. J. Biol. Inorg. Chem. 2014, 19, 747–757. [Google Scholar] [CrossRef] [PubMed]
- Shepard, E.M.; Byer, A.S.; Aggarwal, P.; Betz, J.N.; Scott, A.G.; Shisler, K.A.; Usselman, R.J.; Eaton, G.R.; Eaton, S.S.; Broderick, J.B. Electron Spin Relaxation and Biochemical Characterization of the Hydrogenase Maturase HydF: Insights into [2Fe-2S] and [4Fe-4S] Cluster Communication and Hydrogenase Activation. Biochemistry 2017, 56, 3234–3247. [Google Scholar] [CrossRef] [PubMed]
- Shepard, E.M.; Byer, A.S.; Broderick, J.B. Iron-Sulfur Cluster States of the Hydrogenase Maturase HydF. Biochemistry 2017, 56, 4733–4734. [Google Scholar] [CrossRef] [PubMed]
- Shepard, E.M.; Byer, A.S.; Betz, J.N.; Peters, J.W.; Broderick, J.B. A redox active [2Fe-2S] cluster on the hydrogenase maturase HydF. Biochemistry 2016, 55, 3514–3527. [Google Scholar] [CrossRef] [PubMed]
- Scott, A.G.; Szilagyi, R.K.; Mulder, D.W.; Ratzlo, W.; Byer, A.S.; King, P.W.; Broderick, W.E.; Shepard, M.; Broderick, J.B. Compositional and structural insights into the nature of the H-cluster precursor on HydF. Dalt. Trans. 2018. [Google Scholar] [CrossRef] [PubMed]
- Vallese, F.; Berto, P.; Ruzzene, M.; Cendron, L.; Sarno, S.; De Rosa, E.; Giacometti, G.M.; Costantini, P. Biochemical analysis of the interactions between the proteins involved in the [FeFe]-hydrogenase maturation process. J. Biol. Chem. 2012, 287, 36544–36555. [Google Scholar] [CrossRef] [PubMed]
- Leipe, D.D.; Wolf, Y.I.; Koonin, E.V.; Aravind, L. Classification and evolution of P-loop GTPases and related ATPases. J. Mol. Biol. 2002, 317, 41–72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shepard, E.M.; McGlynn, S.E.; Bueling, A.L.; Grady-Smith, C.S.; George, S.J.; Winslow, M.A.; Cramer, S.P.; Peters, J.W.; Broderick, J.B. Synthesis of the 2Fe subcluster of the [FeFe]-hydrogenase H cluster on the HydF scaffold. Proc. Natl. Acad. Sci. USA 2010, 107, 10448–10453. [Google Scholar] [CrossRef] [PubMed]
- Maso, L.; Galazzo, L.; Vallese, F.; Di Valentin, M.; Albertini, M.; De Rosa, E.; Giacometti, G.M.; Costantini, P.; Carbonera, D. A conformational study of the GTPase domain of [FeFe]-hydrogenase maturation protein HydF by PELDOR spectroscopy. Appl. Magn. Reson. 2015, 46, 465–479. [Google Scholar] [CrossRef]
- McGlynn, S.E.; Ruebush, S.S.; Naumov, A.; Nagy, L.E.; Dubini, A.; King, P.W.; Broderick, J.B.; Posewitz, M.C.; Peters, J.W. In vitro activation of [FeFe] hydrogenase: New insights into hydrogenase maturation. J. Biol. Inorg. Chem. 2007, 12, 443–447. [Google Scholar] [CrossRef] [PubMed]
- De Toni, L.; Guidolin, D.; De Filippis, V.; Tescari, S.; Strapazzon, G.; Rocca, M.S.; Ferlin, A.; Plebani, M.; Foresta, C. Osteocalcin and sex hormone binding globulin compete on a specific binding site of GPRC6A. Endocrinology 2016, 157, 4473–4486. [Google Scholar] [CrossRef] [PubMed]
- Schneidman-Duhovny, D.; Inbar, Y.; Nussinov, R.; Wolfson, H.J. PatchDock and SymmDock: Servers for rigid and symmetric docking. Nucleic Acids Res. 2005, 33, 363–367. [Google Scholar] [CrossRef] [PubMed]
- Andrusier, N.; Nussinov, R.; Wolfson, H.J. FireDock: Fast interaction refinement in molecular docking. Proteins Struct. Funct. Genet. 2007, 69, 139–159. [Google Scholar] [CrossRef] [PubMed]
- Pierce, B.G.; Wiehe, K.; Hwang, H.; Kim, B.H.; Vreven, T.; Weng, Z. ZDOCK server: Interactive docking prediction of protein-protein complexes and symmetric multimers. Bioinformatics 2014, 30, 1771–1773. [Google Scholar] [CrossRef] [PubMed]







© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bortolus, M.; Costantini, P.; Doni, D.; Carbonera, D. Overview of the Maturation Machinery of the H-Cluster of [FeFe]-Hydrogenases with a Focus on HydF. Int. J. Mol. Sci. 2018, 19, 3118. https://doi.org/10.3390/ijms19103118
Bortolus M, Costantini P, Doni D, Carbonera D. Overview of the Maturation Machinery of the H-Cluster of [FeFe]-Hydrogenases with a Focus on HydF. International Journal of Molecular Sciences. 2018; 19(10):3118. https://doi.org/10.3390/ijms19103118
Chicago/Turabian StyleBortolus, Marco, Paola Costantini, Davide Doni, and Donatella Carbonera. 2018. "Overview of the Maturation Machinery of the H-Cluster of [FeFe]-Hydrogenases with a Focus on HydF" International Journal of Molecular Sciences 19, no. 10: 3118. https://doi.org/10.3390/ijms19103118