Hydrophilic Astragalin Galactoside Induces T Helper Type 1-Mediated Immune Responses via Dendritic Cells


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
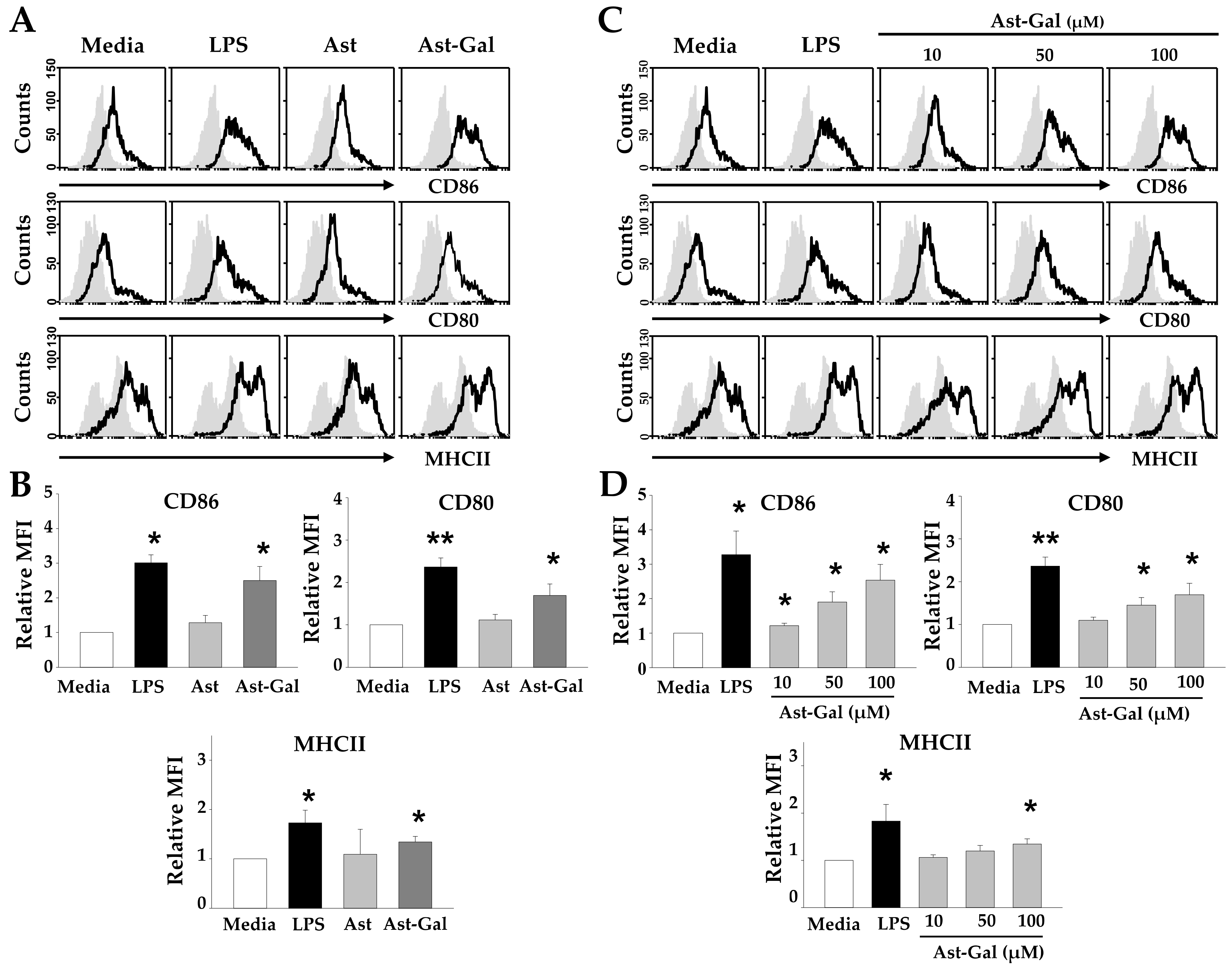
2.1. Hydrophilic Ast-Gal Induces Expression of Surface Molecules on DCs
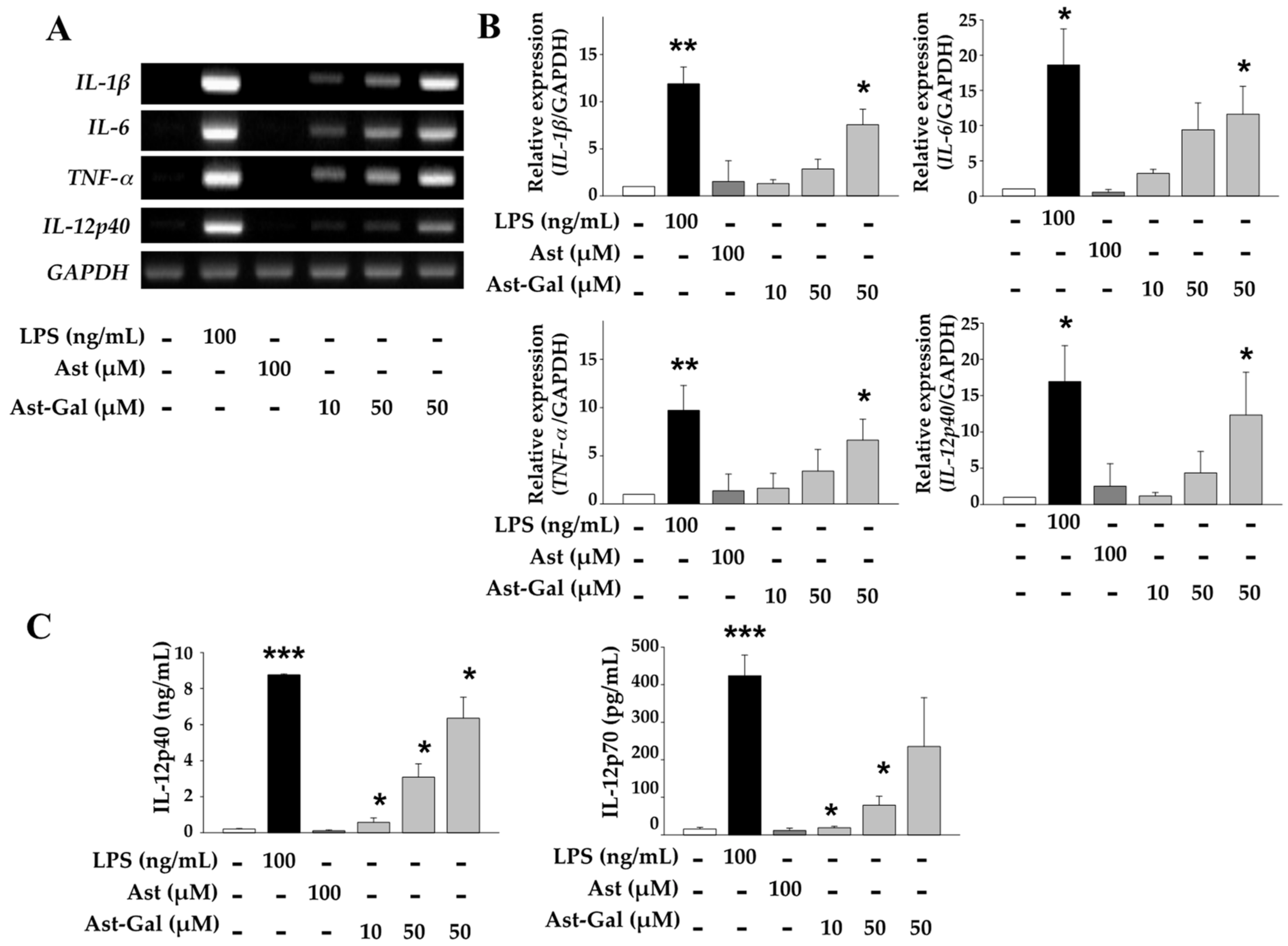
2.2. Increased Expression of Immune-Stimulating Cytokines in Ast-Gal-Treated DCs
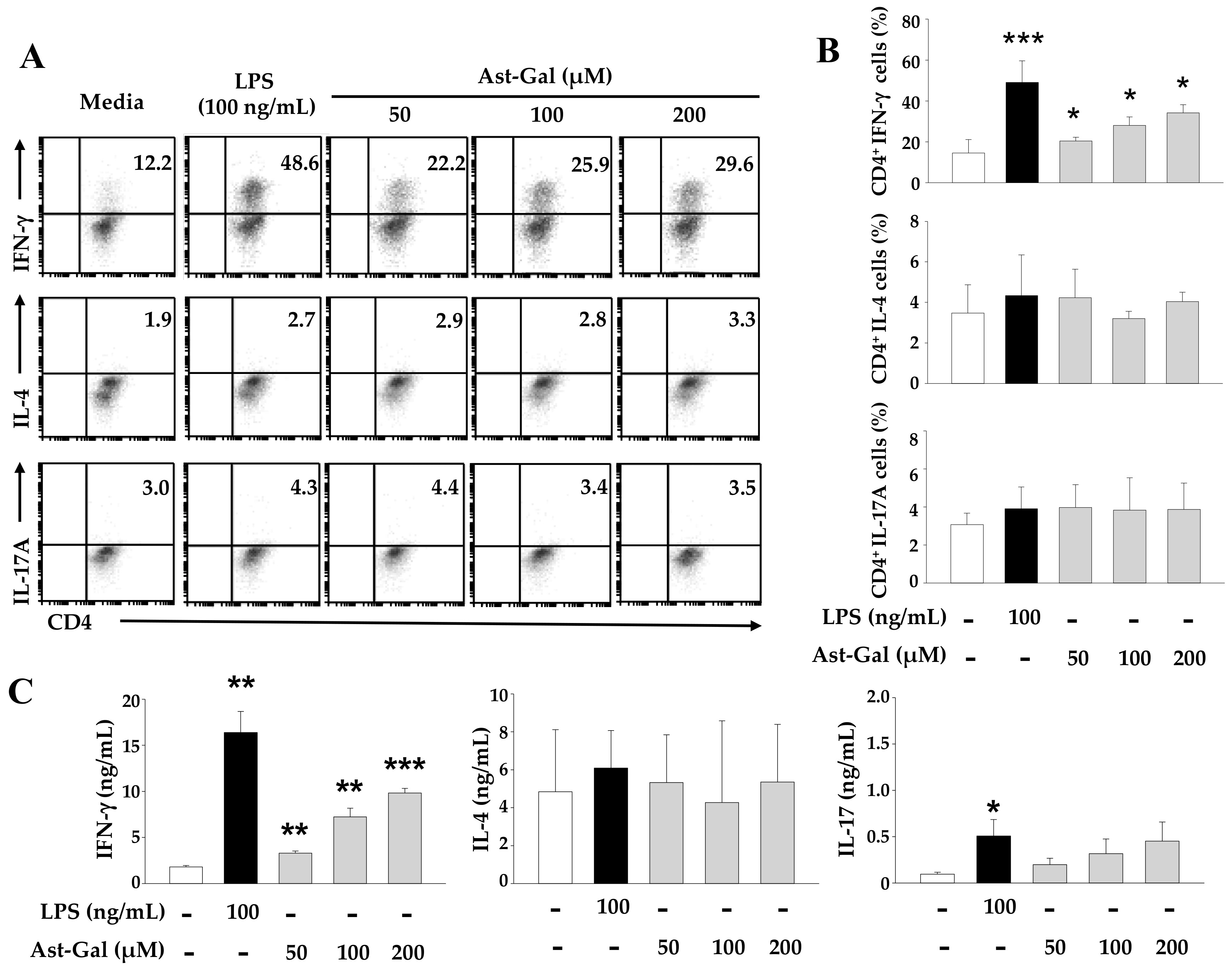
2.3. Ast-Gal-Stimulated DCs Enhance IFN-γ Production in CD4+ T Cells In Vitro
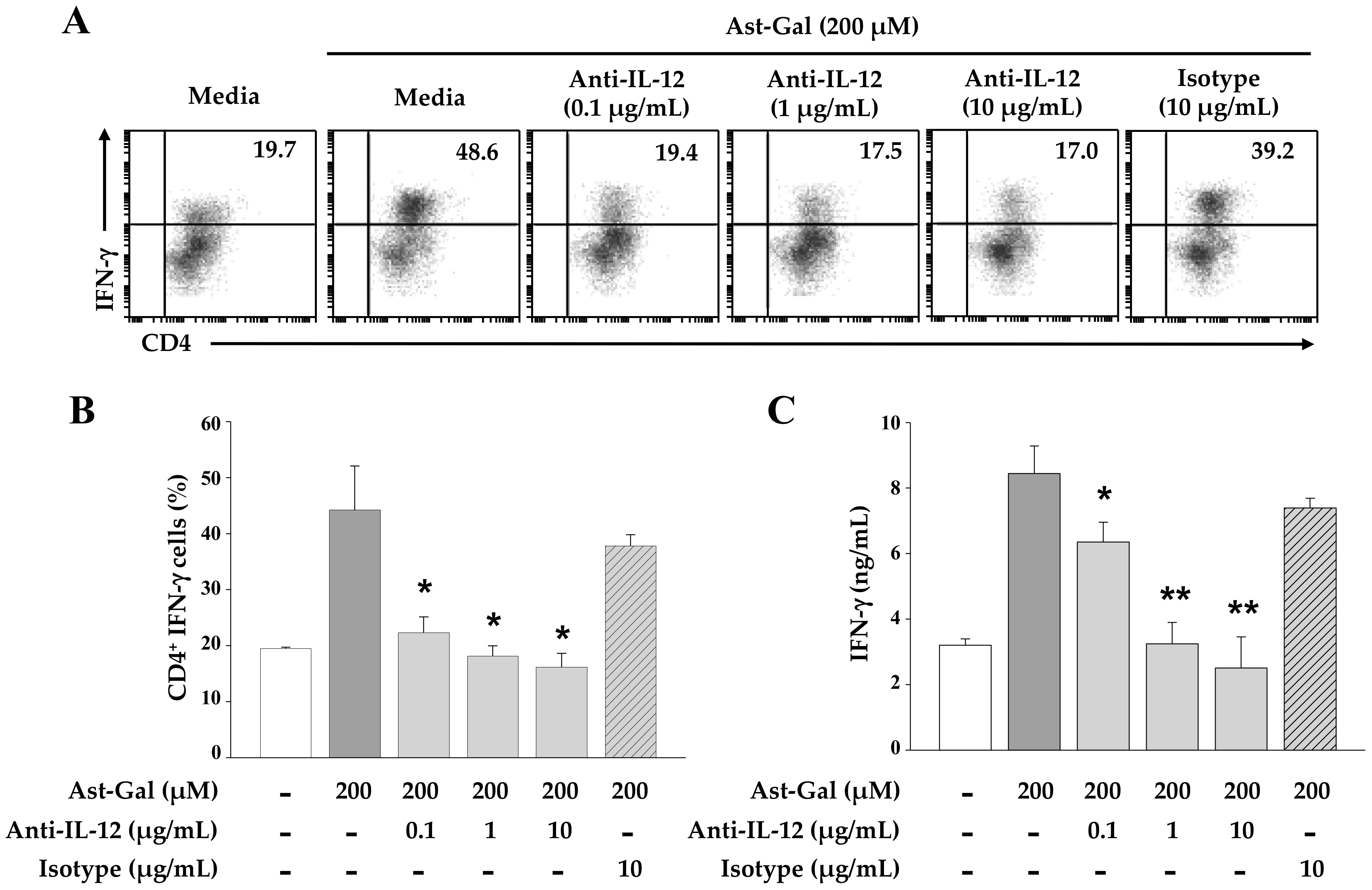
2.4. IL-12 Secreted from the Ast-Gal-Treated DCs Is Involved in the Increased IFN-γ Production in CD4+ T Cells
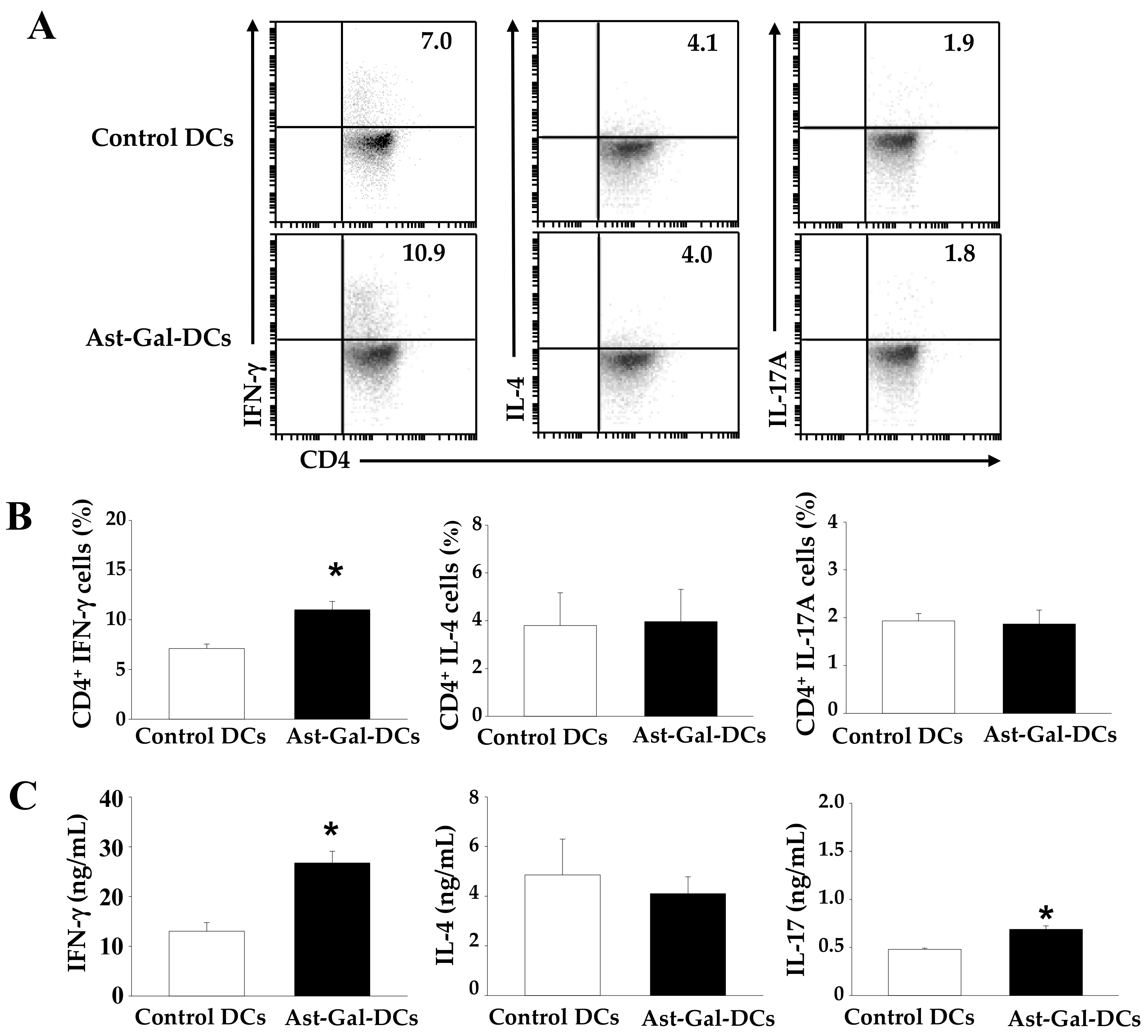
2.5. Ast-Gal-Treated DCs Upregulate IFN-γ Production in CD4+ T Cells In Vivo
3. Discussion
4. Materials and Methods
4.1. Experimental Animals
4.2. Antibodies, Cytokines, and Reagents
4.3. Generation of Bone Marrow DCs
4.4. CD4+ T Cell Isolation and T Cell Polarization In Vitro and In Vivo
4.5. Flow Cytometric Analysis
4.6. Reverse Transcriptase-Polymerase Chain Reaction (RT-PCR)
4.7. Enzyme-Linked Immunosorbent Assay (ELISA)
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| Ast | astragalin |
| Ast-gal | astragalin-galactoside |
| DC | dendritic cell |
| Th1 | T helper type 1 |
| OVA | ovalbumin |
References
- Banchereau, J.; Steinman, R.M. Dendritic cells and the control of immunity. Nature 1998, 392, 245–252. [Google Scholar] [CrossRef] [PubMed]
- Banchereau, J.; Briere, F.; Caux, C.; Davoust, J.; Lebecque, S.; Liu, Y.J.; Pulendran, B.; Palucka, K. Immunobiology of dendritic cells. Annu. Rev. Immunol. 2000, 18, 767–811. [Google Scholar] [CrossRef] [PubMed]
- Steinman, R.M. The dendritic cell system and its role in immunogenicity. Annu. Rev. Immunol. 1991, 9, 271–296. [Google Scholar] [CrossRef] [PubMed]
- Akira, S.; Hemmi, H. Recognition of pathogen-associated molecular patterns by TLR family. Immunol. Lett. 2003, 85, 85–95. [Google Scholar] [CrossRef]
- Guermonprez, P.; Valladeau, J.; Zitvogel, L.; Thery, C.; Amigorena, S. Antigen presentation and T cell stimulation by dendritic cells. Annu. Rev. Immunol. 2002, 20, 621–667. [Google Scholar] [CrossRef] [PubMed]
- Curtsinger, J.M.; Lins, D.C.; Mescher, M.F. Signal 3 determines tolerance versus full activation of naive CD8 T cells: Dissociating proliferation and development of effector function. J. Exp. Med. 2003, 197, 1141–1151. [Google Scholar] [CrossRef] [PubMed]
- Gett, A.V.; Sallusto, F.; Lanzavecchia, A.; Geginat, J. T cell fitness determined by signal strength. Nat. Immunol. 2003, 4, 355–360. [Google Scholar] [CrossRef] [PubMed]
- Lanzavecchia, A.; Sallusto, F. Antigen decoding by T lymphocytes: From synapses to fate determination. Nat. Immunol. 2001, 2, 487–492. [Google Scholar] [CrossRef] [PubMed]
- Valenzuela, J.; Schmidt, C.; Mescher, M. The roles of IL-12 in providing a third signal for clonal expansion of naive CD8 T cells. J. Immunol. 2002, 169, 6842–6849. [Google Scholar] [CrossRef] [PubMed]
- Adams, D.O.; Hamilton, T.A. The cell biology of macrophage activation. Annu. Rev. Immunol. 1984, 2, 283–318. [Google Scholar] [CrossRef] [PubMed]
- Damsker, J.M.; Hansen, A.M.; Caspi, R.R. Th1 and Th17 cells: Adversaries and collaborators. Ann. N. Y. Acad. Sci. 2010, 1183, 211–221. [Google Scholar] [CrossRef] [PubMed]
- Korn, T.; Bettelli, E.; Oukka, M.; Kuchroo, V.K. IL-17 and Th17 Cells. Annu. Rev. Immunol. 2009, 27, 485–517. [Google Scholar] [CrossRef] [PubMed]
- Stevens, T.L.; Bossie, A.; Sanders, V.M.; Fernandez-Botran, R.; Coffman, R.L.; Mosmann, T.R.; Vitetta, E.S. Regulation of antibody isotype secretion by subsets of antigen-specific helper T cells. Nature 1988, 334, 255–258. [Google Scholar] [CrossRef] [PubMed]
- Sakaguchi, S. Naturally arising CD4+ regulatory T cells for immunologic self-tolerance and negative control of immune responses. Annu. Rev. Immunol. 2004, 22, 531–562. [Google Scholar] [CrossRef] [PubMed]
- Yasukawa, K.; Takido, M.; Takeuchi, M.; Sato, Y.; Nitta, K.; Nakagawa, S. Inhibitory effects of flavonol glycosides on 12-O-tetradecanoylphorbol-13-acetate-induced tumor promotion. Chem. Pharm. Bull. 1990, 38, 774–776. [Google Scholar] [CrossRef] [PubMed]
- Higa, S.; Hirano, T.; Kotani, M.; Matsumoto, M.; Fujita, A.; Suemura, M.; Kawase, I.; Tanaka, T. Fisetin, a flavonol, inhibits TH2-type cytokine production by activated human basophils. J. Allergy Clin. Immunol. 2003, 111, 1299–1306. [Google Scholar] [CrossRef] [PubMed]
- Hirano, T.; Arimitsu, J.; Higa, S.; Naka, T.; Ogata, A.; Shima, Y.; Fujimoto, M.; Yamadori, T.; Ohkawara, T.; Kuwabara, Y.; et al. Luteolin, a flavonoid, inhibits CD40 ligand expression by activated human basophils. Int. Arch. Allergy Immunol. 2006, 140, 150–156. [Google Scholar] [CrossRef] [PubMed]
- Ke, M.; Hu, X.Q.; Ouyang, J.; Dai, B.; Xu, Y. The effect of astragalin on the VEGF production of cultured Muller cells under high glucose conditions. Biomed. Mater. Eng. 2012, 22, 113–119. [Google Scholar] [PubMed]
- Kim, M.S.; Kim, S.H. Inhibitory effect of astragalin on expression of lipopolysaccharide-induced inflammatory mediators through NF-kappaB in macrophages. Arch. Pharm. Res. 2011, 34, 2101–2107. [Google Scholar] [CrossRef] [PubMed]
- Soromou, L.W.; Chen, N.; Jiang, L.; Huo, M.; Wei, M.; Chu, X.; Millimouno, F.M.; Feng, H.; Sidime, Y.; Deng, X. Astragalin attenuates lipopolysaccharide-induced inflammatory responses by down-regulating NF-kappaB signaling pathway. Biochem. Biophys. Res. Commun. 2012, 419, 256–261. [Google Scholar] [CrossRef] [PubMed]
- Choi, J.; Kang, H.J.; Kim, S.Z.; Kwon, T.O.; Jeong, S.I.; Jang, S.I. Antioxidant effect of astragalin isolated from the leaves of Morus alba L. against free radical-induced oxidative hemolysis of human red blood cells. Arch. Pharm. Res. 2013, 36, 912–917. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Cheng, Y.; Zhang, X.; Zhang, X.; Chen, S.; Hu, Z.; Zhou, C.; Zhang, E.; Ma, S. Astragalin Attenuates Allergic Inflammation in a Murine Asthma Model. Inflammation 2015, 38, 2007–2016. [Google Scholar] [CrossRef] [PubMed]
- Le Garrec, D.; Gori, S.; Luo, L.; Lessard, D.; Smith, D.C.; Yessine, M.A.; Ranger, M.; Leroux, J.C. Poly(N-vinylpyrrolidone)-block-poly(d,l-lactide) as a new polymeric solubilizer for hydrophobic anticancer drugs: In vitro and in vivo evaluation. J. Control. Release 2004, 99, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Savjani, K.T.; Gajjar, A.K.; Savjani, J.K. Drug solubility: Importance and enhancement techniques. ISRN Pharm. 2012, 2012, 195727. [Google Scholar] [CrossRef] [PubMed]
- Han, S.; Hanh Nguyen, T.T.; Hur, J.; Kim, N.M.; Kim, S.B.; Hwang, K.H.; Moon, Y.H.; Kang, C.; Chung, B.; Kim, Y.M.; et al. Synthesis and characterization of novel astragalin galactosides using beta-galactosidase from Bacillus circulans. Enzym. Microb. Technol. 2017, 103, 59–67. [Google Scholar] [CrossRef] [PubMed]
- De Jong, E.C.; Smits, H.H.; Kapsenberg, M.L. Dendritic cell-mediated T cell polarization. Springer Semin. Immunopathol. 2005, 26, 289–307. [Google Scholar] [CrossRef] [PubMed]
- Bilenki, L.; Wang, S.; Yang, J.; Fan, Y.; Jiao, L.; Joyee, A.G.; Han, X.; Yang, X. Adoptive transfer of CD8α+ dendritic cells (DC) isolated from mice infected with Chlamydia muridarum are more potent in inducing protective immunity than CD8α− DC. J. Immunol. 2006, 177, 7067–7075. [Google Scholar] [CrossRef] [PubMed]
- Czerniecki, B.J.; Koski, G.K.; Koldovsky, U.; Xu, S.; Cohen, P.A.; Mick, R.; Nisenbaum, H.; Pasha, T.; Xu, M.; Fox, K.R.; et al. Targeting HER-2/neu in early breast cancer development using dendritic cells with staged interleukin-12 burst secretion. Cancer Res. 2007, 67, 1842–1852. [Google Scholar] [CrossRef] [PubMed]
- Hokey, D.A.; Larregina, A.T.; Erdos, G.; Watkins, S.C.; Falo, L.D., Jr. Tumor cell loaded type-1 polarized dendritic cells induce Th1-mediated tumor immunity. Cancer Res. 2005, 65, 10059–10067. [Google Scholar] [CrossRef] [PubMed]
- Wesa, A.; Kalinski, P.; Kirkwood, J.M.; Tatsumi, T.; Storkus, W.J. Polarized type-1 dendritic cells (DC1) producing high levels of IL-12 family members rescue patient TH1-type antimelanoma CD4+ T cell responses in vitro. J. Immunother. 2007, 30, 75–82. [Google Scholar] [CrossRef] [PubMed]
- Inaba, K.; Inaba, M.; Romani, N.; Aya, H.; Deguchi, M.; Ikehara, S.; Muramatsu, S.; Steinman, R.M. Generation of large numbers of dendritic cells from mouse bone marrow cultures supplemented with granulocyte/macrophage colony-stimulating factor. J. Exp. Med. 1992, 176, 1693–1702. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeon, J.H.; Lee, B.-C.; Kim, D.; Cho, D.; Kim, T.S. Hydrophilic Astragalin Galactoside Induces T Helper Type 1-Mediated Immune Responses via Dendritic Cells. Int. J. Mol. Sci. 2018, 19, 3120. https://doi.org/10.3390/ijms19103120
Jeon JH, Lee B-C, Kim D, Cho D, Kim TS. Hydrophilic Astragalin Galactoside Induces T Helper Type 1-Mediated Immune Responses via Dendritic Cells. International Journal of Molecular Sciences. 2018; 19(10):3120. https://doi.org/10.3390/ijms19103120
Chicago/Turabian StyleJeon, Jae Hyoung, Byung-Cheol Lee, Doman Kim, Daeho Cho, and Tae Sung Kim. 2018. "Hydrophilic Astragalin Galactoside Induces T Helper Type 1-Mediated Immune Responses via Dendritic Cells" International Journal of Molecular Sciences 19, no. 10: 3120. https://doi.org/10.3390/ijms19103120