β-Naphtoflavone and Ethanol Induce Cytochrome P450 and Protect towards MPP+ Toxicity in Human Neuroblastoma SH-SY5Y Cells

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
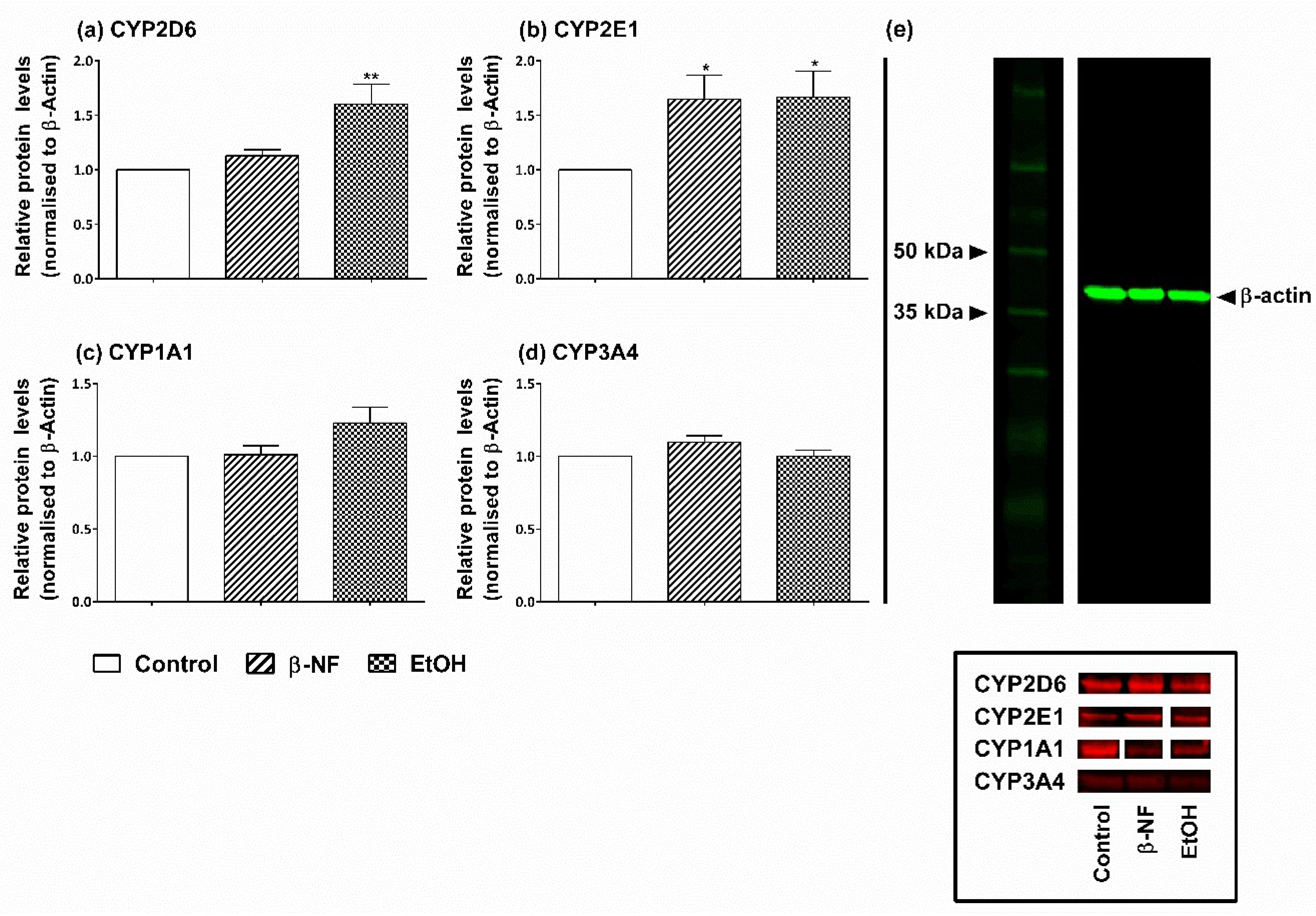
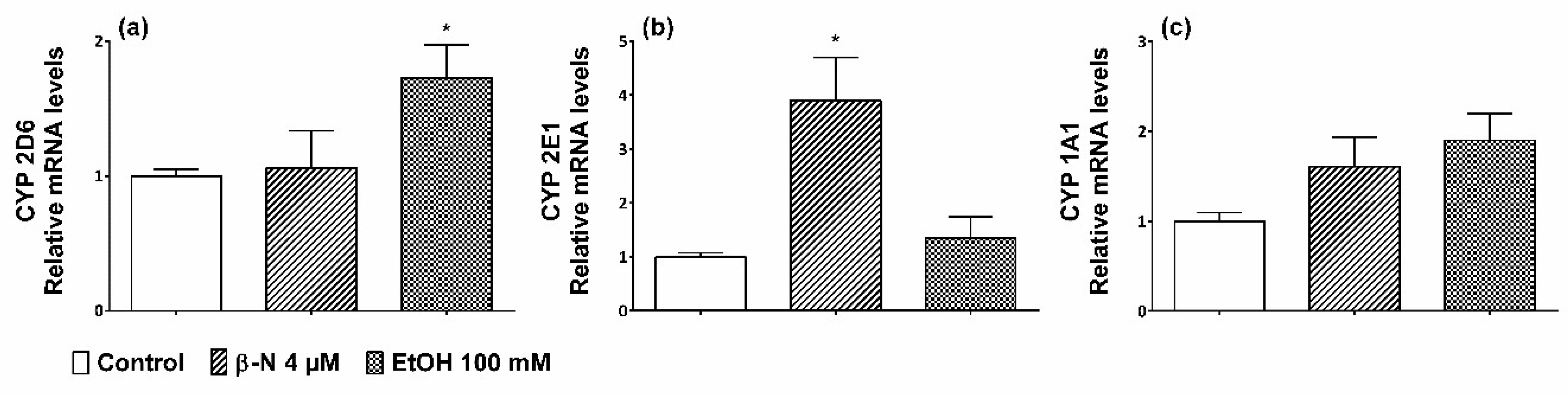
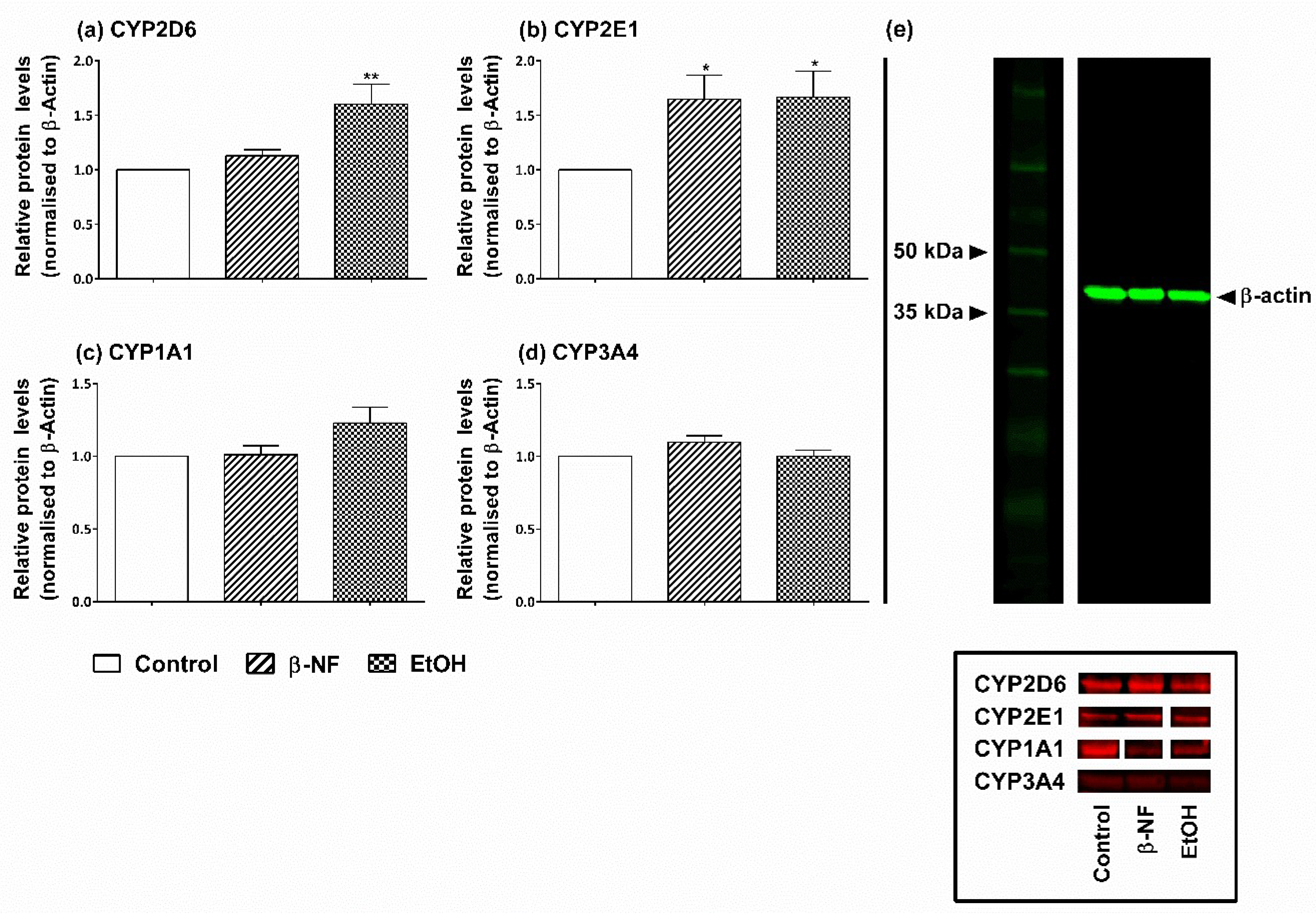
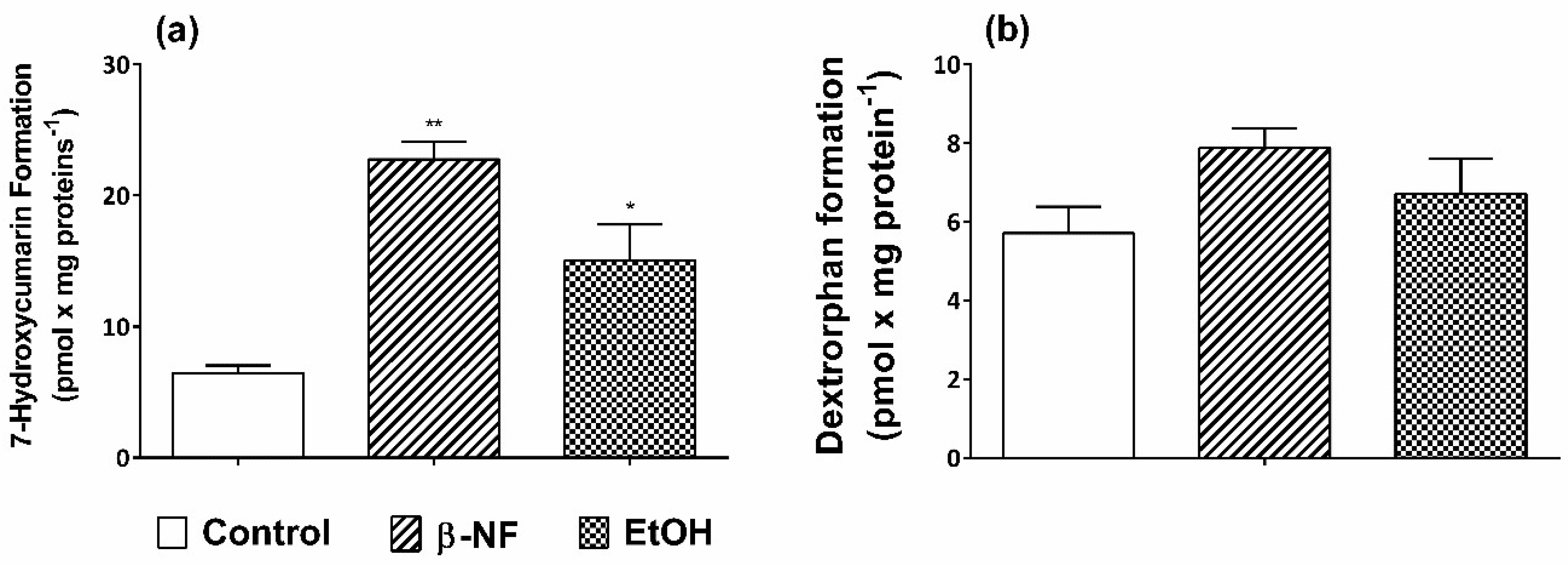
2.1. Induction of CYP2D6 and 2E1
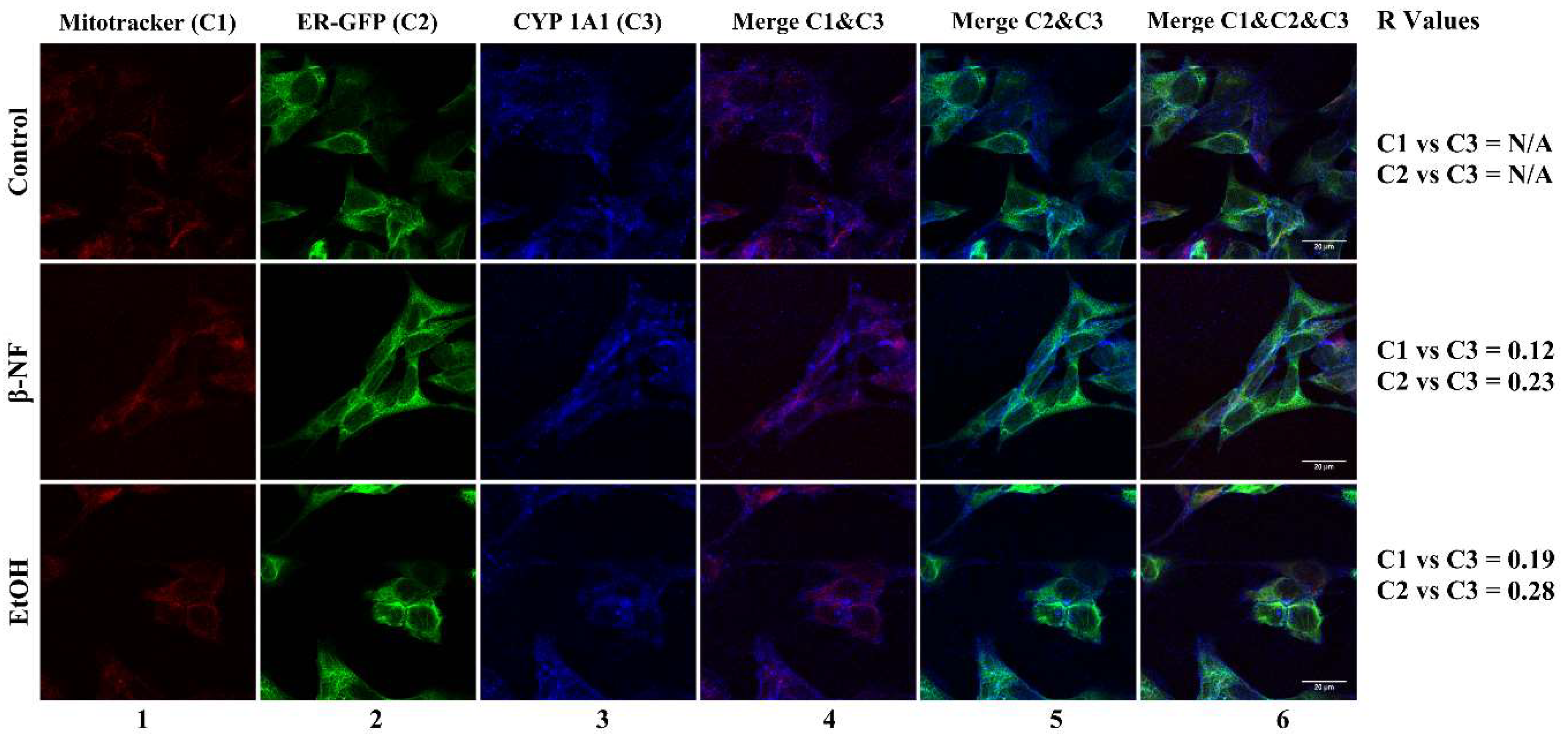
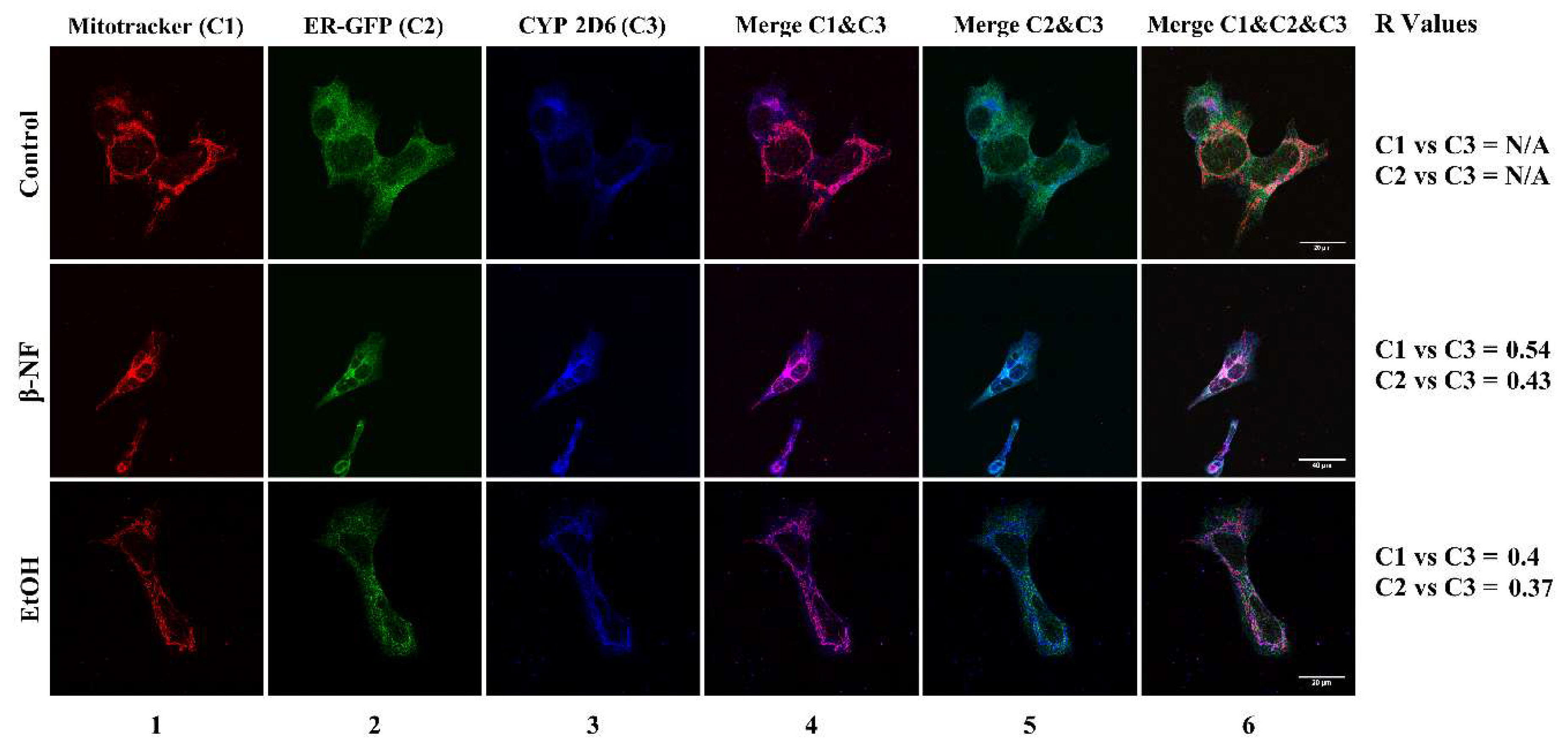
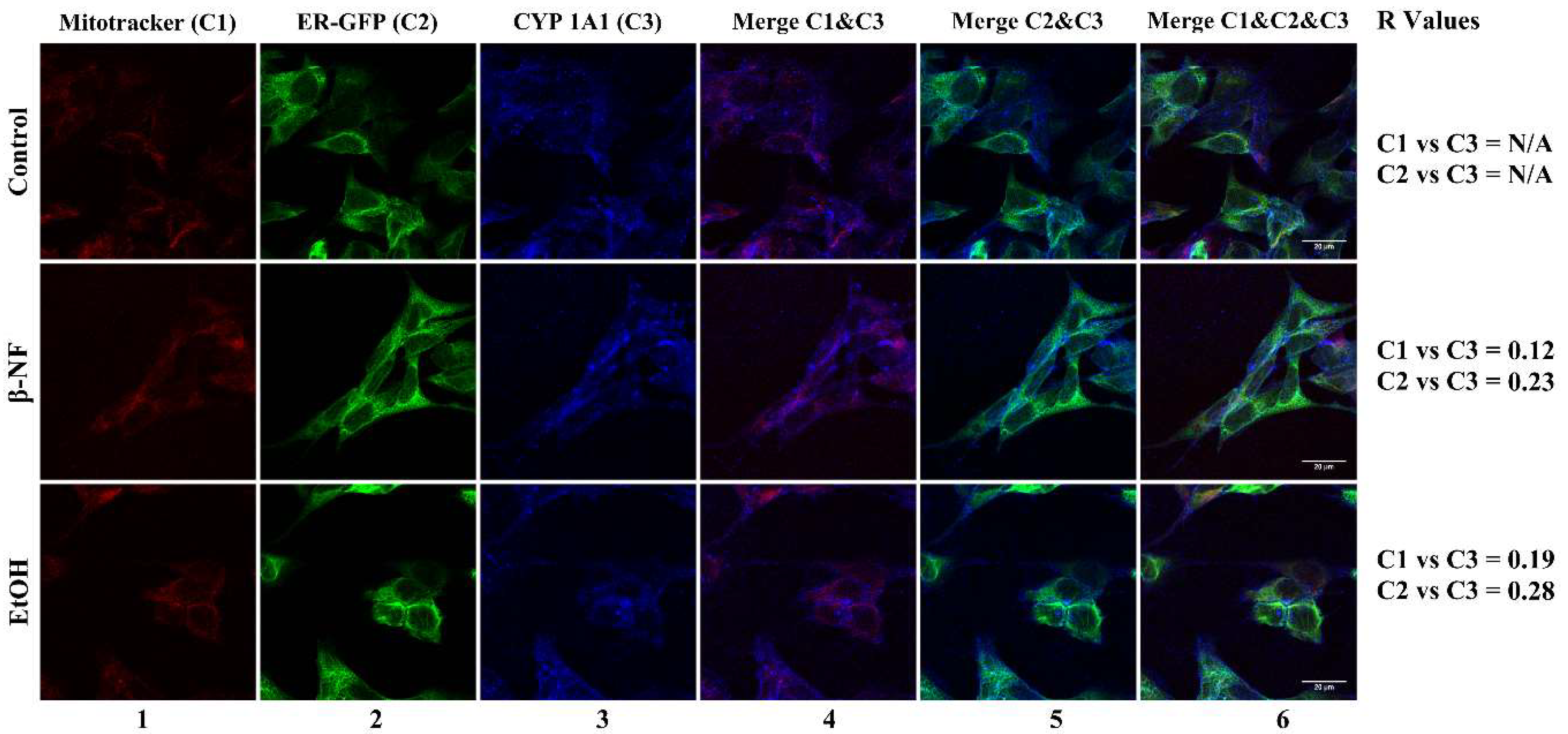
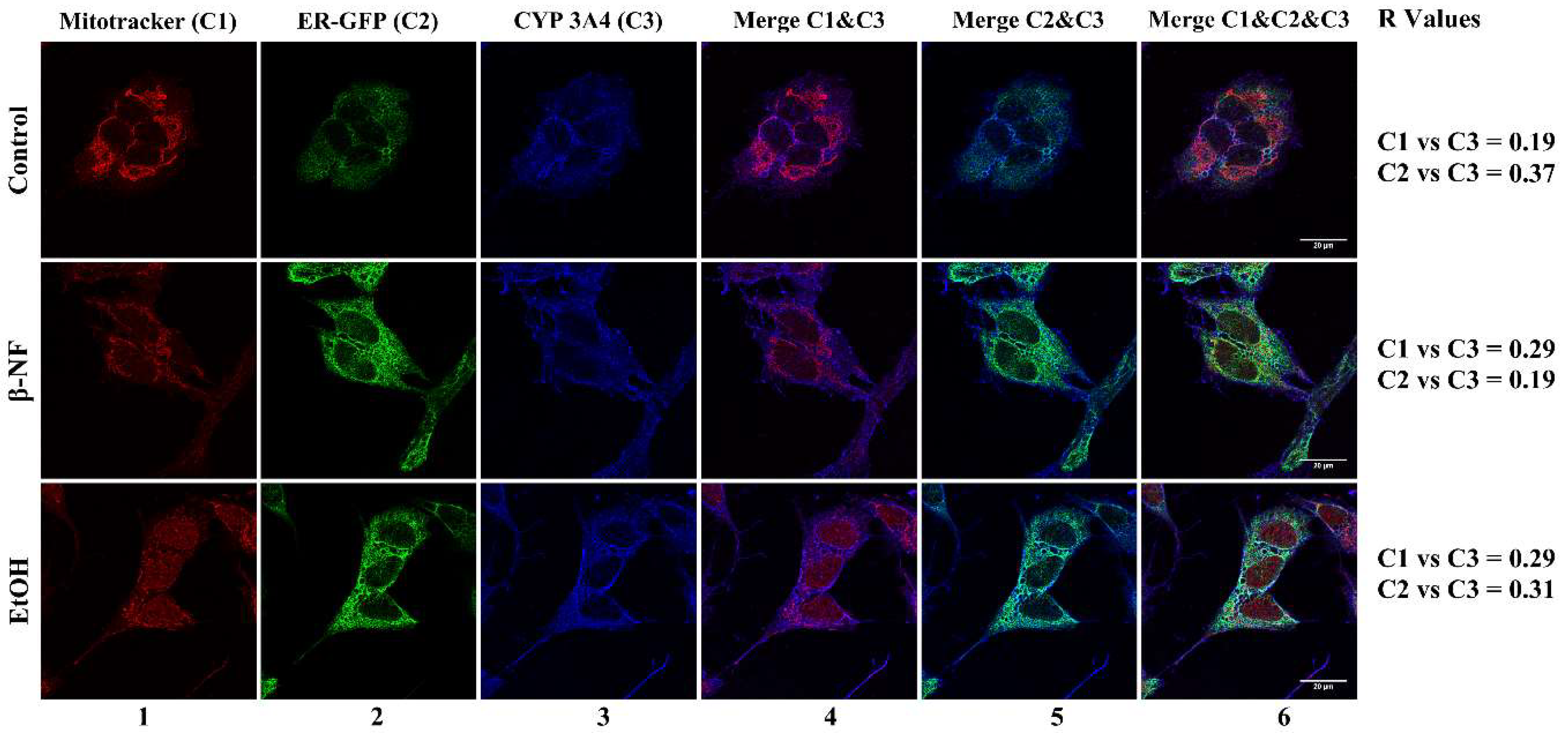
2.2. CYP2D6 Localizes in Mitochondria
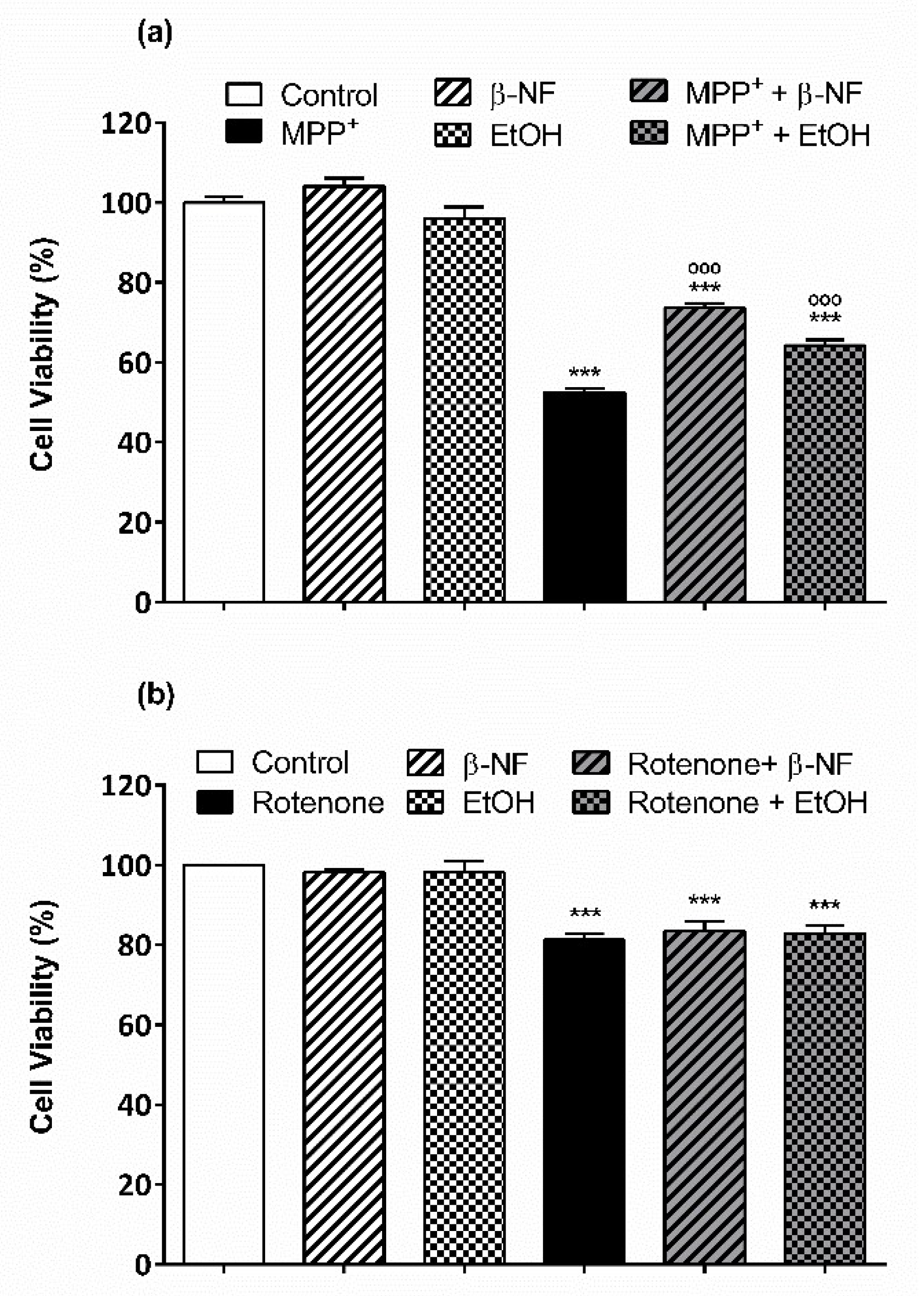
2.3. Protective Effect of CYPs
3. Discussion
4. Materials and Methods
4.1. Cell Culture
4.2. Quantitative Real-Time PCR
4.3. Western Blot and Relative Protein Level Quantification
4.4. Cytochrome P450-Dependent Activity Assay
4.5. Confocal Microscopy
4.6. MTT Assay
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| AhR | Aryl hydrocarbon receptor |
| Ct | Threshold cycle |
| CYP | Cytochrome P450 |
| DMSO | Dimethyl sulfoxide |
| ER | Endoplasmic reticulum |
| ER-GFP | Endoplasmic reticulum-green fluorescent protein |
| EtOH | Ethanol |
| FBS | Fetal bovine serum |
| MPP+ | 1-methyl-4-phenylpyridinium iodide |
| MPTP | 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine |
| qRT-PCR | Real time quantitative-polymerase chain reaction |
| WB | Western blot |
| β-NF | β-naphtoflavone |
References
- Miksys, S.; Tyndale, R.F. Brain drug-metabolizing cytochrome P450 enzymes are active in vivo, demonstrated by mechanism-based enzyme inhibition. Neuropsychopharmacology 2009, 34, 634–640. [Google Scholar] [CrossRef] [PubMed]
- Dutheil, F.; Dauchy, S.; Diry, M.; Sazdovitch, V.; Cloarec, O.; Mellottee, L.; Bieche, I.; Ingelman-Sundberg, M.; Flinois, J.P.; de Waziers, I.; et al. Xenobiotic-metabolizing enzymes and transporters in the normal human brain: Regional and cellular mapping as a basis for putative roles in cerebral function. Drug Metab. Dispos. Biol. Fate Chem. 2009, 37, 1528–1538. [Google Scholar] [CrossRef] [PubMed]
- Santes-Palacios, R.; Ornelas-Ayala, D.; Cabanas, N.; Marroquin-Perez, A.; Hernandez-Magana, A.; Del Rosario Olguin-Reyes, S.; Camacho-Carranza, R.; Espinosa-Aguirre, J.J. Regulation of Human Cytochrome P4501A1 (hCYP1A1): A Plausible Target for Chemoprevention? Biomed. Res. Int. 2016, 2016, 5341081. [Google Scholar] [CrossRef] [PubMed]
- Avadhani, N.G.; Sangar, M.C.; Bansal, S.; Bajpai, P. Bimodal targeting of cytochrome P450s to endoplasmic reticulum and mitochondria: The concept of chimeric signals. FEBS J. 2011, 278, 4218–4229. [Google Scholar] [CrossRef] [PubMed]
- Bansal, S.; Anandatheerthavarada, H.K.; Prabu, G.K.; Milne, G.L.; Martin, M.V.; Guengerich, F.P.; Avadhani, N.G. Human cytochrome P450 2E1 mutations that alter mitochondrial targeting efficiency and susceptibility to ethanol-induced toxicity in cellular models. J. Biol. Chem. 2013, 288, 12627–12644. [Google Scholar] [CrossRef] [PubMed]
- Lavandera, J.; Ruspini, S.; Batlle, A.; Buzaleh, A.M. Cytochrome P450 expression in mouse brain: Specific isoenzymes involved in Phase I metabolizing system of porphyrinogenic agents in both microsomes and mitochondria. Biochem. Cell Biol. 2015, 93, 102–107. [Google Scholar] [CrossRef] [PubMed]
- Dutheil, F.; Jacob, A.; Dauchy, S.; Beaune, P.; Scherrmann, J.M.; Decleves, X.; Loriot, M.A. ABC transporters and cytochromes P450 in the human central nervous system: Influence on brain pharmacokinetics and contribution to neurodegenerative disorders. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1161–1174. [Google Scholar] [CrossRef] [PubMed]
- Miksys, S.; Rao, Y.; Sellers, E.M.; Kwan, M.; Mendis, D.; Tyndale, R.F. Regional and cellular distribution of CYP2D subfamily members in rat brain. Xenobiotica 2000, 30, 547–564. [Google Scholar] [CrossRef] [PubMed]
- Gambaro, S.E.; Robert, M.C.; Tiribelli, C.; Gazzin, S. Role of brain cytochrome P450 mono-oxygenases in bilirubin oxidation-specific induction and activity. Arch. Toxicol. 2016, 90, 279–290. [Google Scholar] [CrossRef] [PubMed]
- Riedl, A.G.; Watts, P.M.; Brown, C.T.; Jenner, P. P450 and heme oxygenase enzymes in the basal ganglia and their roles in Parkinson’s disease. Adv. Neurol. 1999, 80, 271–286. [Google Scholar] [PubMed]
- Shahabi, H.N.; Andersson, D.R.; Nissbrandt, H. Cytochrome P450 2E1 in the substantia nigra: Relevance for dopaminergic neurotransmission and free radical production. Synapse 2008, 62, 379–388. [Google Scholar] [CrossRef] [PubMed]
- Goldman, S.M. Environmental toxins and Parkinson’s disease. Annu. Rev. Pharmacol. Toxicol. 2014, 54, 141–164. [Google Scholar] [CrossRef] [PubMed]
- Kab, S.; Moisan, F.; Elbaz, A. Farming and incidence of motor neuron disease: French nationwide study. Eur. J. Neurol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Feldman, R.G.; Ratner, M.H. The pathogenesis of neurodegenerative disease: Neurotoxic mechanisms of action and genetics. Curr. Opin. Neurol. 1999, 12, 725–731. [Google Scholar] [CrossRef] [PubMed]
- Cacabelos, R. Parkinson’s Disease: From Pathogenesis to Pharmacogenomics. Int. J. Mol. Sci. 2017, 18. [Google Scholar] [CrossRef] [PubMed]
- Boopathi, E.; Anandatheerthavarada, H.K.; Bhagwat, S.V.; Biswas, G.; Fang, J.K.; Avadhani, N.G. Accumulation of mitochondrial P450MT2, NH2-terminal truncated cytochrome P4501A1 in rat brain during chronic treatment with β-naphthoflavone. A role in the metabolism of neuroactive drugs. J. Biol. Chem. 2000, 275, 34415–34423. [Google Scholar] [CrossRef] [PubMed]
- Jenner, P. Parkinson’s disease, pesticides and mitochondrial dysfunction. Trends Neurosci. 2001, 24, 245–247. [Google Scholar] [CrossRef]
- Ravindranath, V.; Strobel, H.W. Cytochrome P450-mediated metabolism in brain: Functional roles and their implications. Expert Opin. Drug Metab. Toxicol. 2013, 9, 551–558. [Google Scholar] [CrossRef] [PubMed]
- Lamba, V.; Lamba, J.; Yasuda, K.; Strom, S.; Davila, J.; Hancock, M.L.; Fackenthal, J.D.; Rogan, P.K.; Ring, B.; Wrighton, S.A.; et al. Hepatic CYP2B6 expression: Gender and ethnic differences and relationship to CYP2B6 genotype and CAR (constitutive androstane receptor) expression. J. Pharmacol. Exp. Ther. 2003, 307, 906–922. [Google Scholar] [CrossRef] [PubMed]
- Li, L.; Li, D.; Heyward, S.; Wang, H. Transcriptional Regulation of CYP2B6 Expression by Hepatocyte Nuclear Factor 3β in Human Liver Cells. PLoS ONE 2016, 11, e0150587. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, C.S.; Tyndale, R.F. Cytochrome P450 enzymes in the brain: Emerging evidence of biological significance. Trends Pharmacol. Sci. 2011, 32, 708–714. [Google Scholar] [CrossRef] [PubMed]
- Lee, A.M.; Miksys, S.; Palmour, R.; Tyndale, R.F. CYP2B6 is expressed in African Green monkey brain and is induced by chronic nicotine treatment. Neuropharmacology 2006, 50, 441–450. [Google Scholar] [CrossRef] [PubMed]
- Sridar, C.; Kenaan, C.; Hollenberg, P.F. Inhibition of bupropion metabolism by selegiline: Mechanism-based inactivation of human CYP2B6 and characterization of glutathione and peptide adducts. Drug Metab. Dispos. Biol. Fate Chem. 2012, 40, 2256–2266. [Google Scholar] [CrossRef] [PubMed]
- Zhu, A.Z.; Cox, L.S.; Nollen, N.; Faseru, B.; Okuyemi, K.S.; Ahluwalia, J.S.; Benowitz, N.L.; Tyndale, R.F. CYP2B6 and bupropion’s smoking-cessation pharmacology: The role of hydroxybupropion. Clin. Pharmacol. Ther. 2012, 92, 771–777. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Faucette, S.; Sueyoshi, T.; Moore, R.; Ferguson, S.; Negishi, M.; LeCluyse, E.L. A novel distal enhancer module regulated by pregnane X receptor/constitutive androstane receptor is essential for the maximal induction of CYP2B6 gene expression. J. Biol. Chem. 2003, 278, 14146–14152. [Google Scholar] [CrossRef] [PubMed]
- Elshenawy, O.H.; Abdelhamid, G.; Soshilov, A.A.; Denison, M.S.; El-Kadi, A.O. Down-regulation of cytochrome P450 1A1 by monomethylarsonous acid in human HepG2 cells. Toxicol. Lett. 2017, 270, 34–50. [Google Scholar] [CrossRef] [PubMed]
- Abiko, Y.; Lin, F.Y.; Lee, H.; Puga, A.; Kumagai, Y. Quinone-mediated induction of cytochrome P450 1A1 in HepG2 cells through increased interaction of aryl hydrocarbon receptor with aryl hydrocarbon receptor nuclear translocator. J. Toxicol. Sci. 2016, 41, 775–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toselli, F.; de Waziers, I.; Dutheil, M.; Vincent, M.; Wilce, P.A.; Dodd, P.R.; Beaune, P.; Loriot, M.A.; Gillam, E.M. Gene expression profiling of cytochromes P450, ABC transporters and their principal transcription factors in the amygdala and prefrontal cortex of alcoholics, smokers and drug-free controls by qRT-PCR. Xenobiotica 2015, 45, 1129–1137. [Google Scholar] [CrossRef] [PubMed]
- Nannelli, A.; Rossignolo, F.; Tolando, R.; Rossato, P.; Longo, V.; Gervasi, P.G. Effect of β-naphthoflavone on AhR-regulated genes (CYP1A1, 1A2, 1B1, 2S1, Nrf2, and GST) and antioxidant enzymes in various brain regions of pig. Toxicology 2009, 265, 69–79. [Google Scholar] [CrossRef] [PubMed]
- Zanger, U.M.; Schwab, M. Cytochrome P450 enzymes in drug metabolism: Regulation of gene expression, enzyme activities, and impact of genetic variation. Pharmacol. Ther. 2013, 138, 103–141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, R.; Xu, C.; Chen, X.; Chen, P.; Wang, Y.; Zhou, X.; Jin, J.; Niu, L.; Ying, M.; Huang, M.; et al. Resveratrol suppresses the inducible expression of CYP3A4 through the pregnane X receptor. J. Pharmacol. Sci. 2014, 126, 146–154. [Google Scholar] [CrossRef] [PubMed]
- Du, Z.H.; Xia, J.; Sun, X.C.; Li, X.N.; Zhang, C.; Zhao, H.S.; Zhu, S.Y.; Li, J.L. A novel nuclear xenobiotic receptors (AhR/PXR/CAR)-mediated mechanism of DEHP-induced cerebellar toxicity in quails (Coturnix japonica) via disrupting CYP enzyme system homeostasis. Environ. Pollut. 2017. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, C.; Hossain, M.; Spriggs, A.; Ghosh, A.; Grant, G.A.; Marchi, N.; Perucca, E.; Janigro, D. Sertraline-induced potentiation of the CYP3A4-dependent neurotoxicity of carbamazepine: An in vitro study. Epilepsia 2015, 56, 439–449. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ghosh, C.; Marchi, N.; Desai, N.K.; Puvenna, V.; Hossain, M.; Gonzalez-Martinez, J.; Alexopoulos, A.V.; Janigro, D. Cellular localization and functional significance of CYP3A4 in the human epileptic brain. Epilepsia 2011, 52, 562–571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mann, A.; Tyndale, R.F. Cytochrome P450 2D6 enzyme neuroprotects against 1-methyl-4-phenylpyridinium toxicity in SH-SY5Y neuronal cells. Eur. J. Neurosci. 2010, 31, 1185–1193. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Uehara, S.; Uno, Y.; Inoue, T.; Murayama, N.; Shimizu, M.; Sasaki, E.; Yamazaki, H. Activation and deactivation of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine by cytochrome P450 enzymes and flavin-containing monooxygenases in common marmosets (Callithrix jacchus). Drug Metab. Dispos. Biol. Fate Chem. 2015, 43, 735–742. [Google Scholar] [CrossRef] [PubMed]
- Bromek, E.; Haduch, A.; Daniel, W.A. The ability of cytochrome P450 2D isoforms to synthesize dopamine in the brain: An in vitro study. Eur. J. Neurosci. 2010, 626, 171–178. [Google Scholar] [CrossRef] [PubMed]
- Hiroi, T.; Imaoka, S.; Funae, Y. Dopamine formation from tyramine by CYP2D6. Biochem. Biophys. Res. Commun. 1998, 249, 838–843. [Google Scholar] [CrossRef] [PubMed]
- Niwa, T.; Shizuku, M.; Yamano, K. Effect of genetic polymorphism on the inhibition of dopamine formation from p-tyramine catalyzed by brain cytochrome P450 2D6. Arch. Biochem. Biophys. 2017, 620, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Peng, Q.; Zeng, Z.; Wang, J.; Deng, Y.; Xie, L.; Mo, C.; Zeng, J.; Qin, X.; Li, S. CYP2D6 phenotypes and Parkinson’s disease risk: A meta-analysis. J. Neurol. Sci. 2014, 336, 161–168. [Google Scholar] [CrossRef] [PubMed]
- Mann, A.; Miksys, S.; Lee, A.; Mash, D.C.; Tyndale, R.F. Induction of the drug metabolizing enzyme CYP2D in monkey brain by chronic nicotine treatment. Neuropharmacology 2008, 55, 1147–1155. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yue, J.; Miksys, S.; Hoffmann, E.; Tyndale, R.F. Chronic nicotine treatment induces rat CYP2D in the brain but not in the liver: An investigation of induction and time course. J. Psychiatry Neurosci. 2008, 33, 54–63. [Google Scholar] [PubMed]
- Bertilsson, L. Metabolism of antidepressant and neuroleptic drugs by cytochrome p450s: Clinical and interethnic aspects. Clin. Pharmacol. Ther. 2007, 82, 606–609. [Google Scholar] [CrossRef] [PubMed]
- Cheng, J.; Zhen, Y.; Miksys, S.; Beyoglu, D.; Krausz, K.W.; Tyndale, R.F.; Yu, A.; Idle, J.R.; Gonzalez, F.J. Potential role of CYP2D6 in the central nervous system. Xenobiotica 2013, 43, 973–984. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, Z.X.; Chen, X.W.; Zhou, Z.W.; Zhou, S.F. Impact of physiological, pathological and environmental factors on the expression and activity of human cytochrome P450 2D6 and implications in precision medicine. Drug Metab. Rev. 2015, 47, 470–519. [Google Scholar] [CrossRef] [PubMed]
- Schmid, Y.; Vizeli, P.; Hysek, C.M.; Prestin, K.; Meyer Zu Schwabedissen, H.E.; Liechti, M.E. CYP2D6 function moderates the pharmacokinetics and pharmacodynamics of 3,4-methylene-dioxymethamphetamine in a controlled study in healthy individuals. Pharmacogenet. Genom. 2016, 26, 397–401. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dragoni, S.; Bellik, L.; Frosini, M.; Matteucci, G.; Sgaragli, G.; Valoti, M. Cytochrome P450-dependent metabolism of l-deprenyl in monkey (Cercopithecus aethiops) and C57BL/6 mouse brain microsomal preparations. J. Neurochem. 2003, 86, 1174–1180. [Google Scholar] [PubMed]
- Dragoni, S.; Bellik, L.; Frosini, M.; Sgaragli, G.; Marini, S.; Gervasi, P.G.; Valoti, M. l-Deprenyl metabolism by the cytochrome P450 system in monkey (Cercopithecus aethiops) liver microsomes. Xenobiotica 2003, 33, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Valoti, M.; Fusi, F.; Frosini, M.; Pessina, F.; Tipton, K.F.; Sgaragli, G.P. Cytochrome P450-dependent N-dealkylation of l-deprenyl in C57BL mouse liver microsomes: Effects of in vivo pretreatment with ethanol, phenobarbital, beta-naphthoflavone and l-deprenyl. Eur. J. Pharmacol. 2000, 391, 199–206. [Google Scholar] [CrossRef]
- Vaglini, F.; Viaggi, C.; Piro, V.; Pardini, C.; Gerace, C.; Scarselli, M.; Corsini, G.U. Acetaldehyde and Parkinsonism: Role of CYP450 2E1. Front. Behav. Neurosci. 2013, 7, 71. [Google Scholar] [CrossRef] [PubMed]
- Na, S.; Li, J.; Zhang, H.; Li, Y.; Yang, Z.; Zhong, Y.; Dong, G.; Yang, J.; Yue, J. The induction of cytochrome P450 2E1 by ethanol leads to the loss of synaptic proteins via PPARalpha down-regulation. Toxicology 2017, 385, 18–27. [Google Scholar] [CrossRef] [PubMed]
- Pardini, C.; Vaglini, F.; Viaggi, C.; Caramelli, A.; Corsini, G.U. Role of CYP2E1 in the mouse model of MPTP toxicity. Parkinsonism Relat. Disord. 2008, 14, S119–S123. [Google Scholar] [CrossRef] [PubMed]
- Garcia-Suastegui, W.A.; Ramos-Chavez, L.A.; Rubio-Osornio, M.; Calvillo-Velasco, M.; Atzin-Mendez, J.A.; Guevara, J.; Silva-Adaya, D. The Role of CYP2E1 in the Drug Metabolism or Bioactivation in the Brain. Oxidat. Med. Cell. Longev. 2017, 2017, 4680732. [Google Scholar] [CrossRef] [PubMed]
- Vaglini, F.; Pardini, C.; Viaggi, C.; Corsini, G.U. Cytochrome p450 and Parkinsonism: Protective role of CYP2E1. Funct. Neurol. 2001, 16, 107–112. [Google Scholar] [PubMed]
- Naselli, F.; Catanzaro, I.; Bellavia, D.; Perez, A.; Sposito, L.; Caradonna, F. Role and importance of polymorphisms with respect to DNA methylation for the expression of CYP2E1 enzyme. Gene 2014, 536, 29–39. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xicoy, H.; Wieringa, B.; Martens, G.J. The SH-SY5Y cell line in Parkinson’s disease research: A systematic review. Mol. Neurodegener. 2017, 12, 10. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.K.; Kumar, V.; Pandey, A.; Vatsa, P.; Dhasmana, A.; Singh, R.P.; Appikonda, S.H.C.; Hwang, I.; Lohani, M. Monocrotophos Induces the Expression of Xenobiotic Metabolizing Cytochrome P450s (CYP2C8 and CYP3A4) and Neurotoxicity in Human Brain Cells. Mol. Neurobiol. 2017, 54, 3633–3651. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, V.K.; Kumar, V.; Singh, A.K.; Kashyap, M.P.; Jahan, S.; Pandey, A.; Alam, S.; Khan, F.; Khanna, V.K.; Yadav, S.; et al. Monocrotophos induces the expression and activity of xenobiotic metabolizing enzymes in pre-sensitized cultured human brain cells. PLoS ONE 2014, 9, e91946. [Google Scholar] [CrossRef] [PubMed]
- Brauze, D.; Fijalkiewicz, K.; Szaumkessel, M.; Kiwerska, K.; Bednarek, K.; Rydzanicz, M.; Richter, J.; Grenman, R.; Jarmuz-Szymczak, M. Diversified expression of aryl hydrocarbon receptor dependent genes in human laryngeal squamous cell carcinoma cell lines treated with β-naphthoflavone. Toxicol. Lett. 2014, 231, 99–107. [Google Scholar] [CrossRef] [PubMed]
- Pretti, C.; Salvetti, A.; Longo, V.; Giorgi, M.; Gervasi, P.G. Effects of beta-naphthoflavone on the cytochrome P450 system, and phase II enzymes in gilthead seabream (Sparus aurata). Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2001, 130, 133–144. [Google Scholar] [CrossRef]
- Shahi, G.S.; Das, N.P.; Moochhala, S.M. 1-Methyl-4-phenyl-1,2,3,6-tetrahydropyridine-induced neurotoxicity: Partial protection against striato-nigral dopamine depletion in C57BL/6J mice by cigarette smoke exposure and by beta-naphthoflavone-pretreatment. Neurosci. Lett. 1991, 127, 247–250. [Google Scholar] [CrossRef]
- Waxman, D.J.; Chang, T.K. Use of 7-ethoxycoumarin to monitor multiple enzymes in the human CYP1, CYP2, and CYP3 families. Methods Mol. Biol. 2006, 320, 153–156. [Google Scholar] [CrossRef] [PubMed]
- Yu, A.; Haining, R.L. Comparative contribution to dextromethorphan metabolism by cytochrome P450 isoforms in vitro: Can dextromethorphan be used as a dual probe for both CTP2D6 and CYP3A activities? Drug Metab. Dispos. Biol. Fate Chem. 2001, 29, 1514–1520. [Google Scholar] [PubMed]
- Imran, S.; Ferretti, P.; Vrzal, R. Different regulation of aryl hydrocarbon receptor-regulated genes in response to dioxin in undifferentiated and neuronally differentiated human neuroblastoma SH-SY5Y cells. Toxicol. Mech. Methods 2015, 25, 689–697. [Google Scholar] [CrossRef] [PubMed]
- Hellum, B.H.; Nilsen, O.G. The in vitro inhibitory potential of trade herbal products on human CYP2D6-mediated metabolism and the influence of ethanol. Basic Clin. Pharmacol. Toxicol. 2007, 101, 350–358. [Google Scholar] [CrossRef] [PubMed]
- Zhang, F.; Li, J.; Na, S.; Wu, J.; Yang, Z.; Xie, X.; Wan, Y.; Li, K.; Yue, J. The Involvement of PPARs in the Selective Regulation of Brain CYP2D by Growth Hormone. Neuroscience 2018, 379, 115–125. [Google Scholar] [CrossRef] [PubMed]
- Roberts, B.J.; Song, B.J.; Soh, Y.; Park, S.S.; Shoaf, S.E. Ethanol induces CYP2E1 by protein stabilization. Role of ubiquitin conjugation in the rapid degradation of CYP2E1. J. Biol. Chem. 1995, 270, 29632–29635. [Google Scholar] [PubMed]
- Ledesma, J.C.; Miquel, M.; Pascual, M.; Guerri, C.; Aragon, C.M. Induction of brain cytochrome P450 2E1 boosts the locomotor-stimulating effects of ethanol in mice. Neuropharmacology 2014, 85, 36–44. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McGehee, R.E., Jr.; Ronis, M.J.; Badger, T.M. Regulation of the hepatic CYP 2E1 gene during chronic alcohol exposure: Lack of an ethanol response element in the proximal 5′-flanking sequence. DNA Cell Biol. 1997, 16, 725–736. [Google Scholar] [CrossRef] [PubMed]
- Howard, L.A.; Miksys, S.; Hoffmann, E.; Mash, D.; Tyndale, R.F. Brain CYP2E1 is induced by nicotine and ethanol in rat and is higher in smokers and alcoholics. Br. J. Pharmacol. 2003, 138, 1376–1386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gambaro, S.E.; Moretti, R.; Tiribelli, C.; Gazzin, S. Brain Cytochrome p450 enzymes: A possible Therapeutic Targets for Neurological Diseases. Ther. Targets Neurol. Dis. 2015, 2. [Google Scholar] [CrossRef]
- Singh, N.K.; Banerjee, B.D.; Bala, K.; Basu, M.; Chhillar, N. Polymorphism in Cytochrome P450 2D6, Glutathione S-Transferases Pi 1 Genes, and Organochlorine Pesticides in Alzheimer Disease: A Case-Control Study in North Indian Population. J. Geriatr. Psychiatry Neurol. 2014, 27, 119–127. [Google Scholar] [CrossRef] [PubMed]
- Ramsay, R.R.; Majekova, M.; Medina, M.; Valoti, M. Key Targets for Multi-Target Ligands Designed to Combat Neurodegeneration. Front. Neurosci. 2016, 10, 375. [Google Scholar] [CrossRef] [PubMed]
- Viaggi, C.; Pardini, C.; Vaglini, F.; Corsini, G.U. Cytochrome P450 and Parkinson’s disease: Protective role of neuronal CYP 2E1 from MPTP toxicity. J. Neural Transm. Suppl. 2006, 70, 173–176. [Google Scholar]
- Viaggi, C.; Vaglini, F.; Pardini, C.; Caramelli, A.; Corsini, G.U. MPTP-induced model of Parkinson’s disease in cytochrome P450 2E1 knockout mice. Neuropharmacology 2009, 56, 1075–1081. [Google Scholar] [CrossRef] [PubMed]
- Kaut, O.; Schmitt, I.; Wullner, U. Genome-scale methylation analysis of Parkinson’s disease patients’ brains reveals DNA hypomethylation and increased mRNA expression of cytochrome P450 2E1. Neurogenetics 2012, 13, 87–91. [Google Scholar] [CrossRef] [PubMed]
- Shimada, T.; Tsumura, F.; Yamazaki, H. Prediction of human liver microsomal oxidations of 7-ethoxycoumarin and chlorzoxazone with kinetic parameters of recombinant cytochrome P-450 enzymes. Drug Metab. Dispos. Biol. Fate Chem. 1999, 27, 1274–1280. [Google Scholar] [PubMed]
- Marini, S.; Nannelli, A.; Sodini, D.; Dragoni, S.; Valoti, M.; Longo, V.; Gervasi, P.G. Expression, microsomal and mitochondrial activities of cytochrome P450 enzymes in brain regions from control and phenobarbital-treated rabbits. Life Sci. 2007, 80, 910–917. [Google Scholar] [CrossRef] [PubMed]
- Miksys, S.L.; Tyndale, R.F. Drug-metabolizing cytochrome P450s in the brain. J. Psychiatry Neurosci. 2002, 27, 406–415. [Google Scholar] [PubMed]
- Bajpai, P.; Sangar, M.C.; Singh, S.; Tang, W.; Bansal, S.; Chowdhury, G.; Cheng, Q.; Fang, J.K.; Martin, M.V.; Guengerich, F.P.; et al. Metabolism of 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine by mitochondrion-targeted cytochrome P450 2D6: Implications in Parkinson disease. J. Biol. Chem. 2013, 288, 4436–4451. [Google Scholar] [CrossRef] [PubMed]
- Sangar, M.C.; Anandatheerthavarada, H.K.; Martin, M.V.; Guengerich, F.P.; Avadhani, N.G. Identification of genetic variants of human cytochrome P450 2D6 with impaired mitochondrial targeting. Mol. Genet. Metab. 2010, 99, 90–97. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sangar, M.C.; Bansal, S.; Avadhani, N.G. Bimodal targeting of microsomal cytochrome P450s to mitochondria: Implications in drug metabolism and toxicity. Expert Opin. Drug Metab. Toxicol. 2010, 6, 1231–1251. [Google Scholar] [CrossRef] [PubMed]
- Booth Depaz, I.M.; Toselli, F.; Wilce, P.A.; Gillam, E.M. Differential expression of human cytochrome P450 enzymes from the CYP3A subfamily in the brains of alcoholic subjects and drug-free controls. Drug Metab. Dispos. Biol. Fate Chem. 2013, 41, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Yamaori, S.; Maeda, C.; Ogawa, R.; Okazaki, H.; Aramaki, H.; Watanabe, K. Expression levels of 39 Cyp mRNAs in the mouse brain and neuroblastoma cell lines, C-1300N18 and NB2a—Strong expression of Cyp1b1. Fund. Toxicol. Sci. 2017, 4, 6. [Google Scholar] [CrossRef]
- Valencia-Olvera, A.C.; Moran, J.; Camacho-Carranza, R.; Prospero-Garcia, O.; Espinosa-Aguirre, J.J. CYP2E1 induction leads to oxidative stress and cytotoxicity in glutathione-depleted cerebellar granule neurons. Toxicol. In Vitro 2014, 28, 1206–1214. [Google Scholar] [CrossRef] [PubMed]
- Hao, C.; Liu, W.; Luan, X.; Li, Y.; Gui, H.; Peng, Y.; Shen, J.; Hu, G.; Yang, J. Aquaporin-4 knockout enhances astrocyte toxicity induced by 1-methyl-4-phenylpyridinium ion and lipopolysaccharide via increasing the expression of cytochrome P4502E1. Toxicol. Lett. 2010, 198, 225–231. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miksys, S.; Tyndale, R.F. Cytochrome P450-mediated drug metabolism in the brain. J. Psychiatry Neurosci. 2013, 38, 152–163. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Gao, S.S.; Yang, H.J.; Wang, M.; Cheng, B.F.; Feng, Z.W.; Wang, L. Neuroprotective Effects of Proanthocyanidins, Natural Flavonoids Derived From Plants, on Rotenone-Induced Oxidative Stress and Apoptotic Cell Death in Human Neuroblastoma SH-SY5Y Cells. Front. Neurosci. 2018, 12, 369. [Google Scholar] [CrossRef] [PubMed]
- Gay, S.C.; Roberts, A.G.; Halpert, J.R. Structural features of cytochromes P450 and ligands that affect drug metabolism as revealed by X-ray crystallography and NMR. Future Med. Chem. 2010, 2, 1451–1468. [Google Scholar] [CrossRef] [PubMed]
- Cheung, Y.T.; Lau, W.K.; Yu, M.S.; Lai, C.S.; Yeung, S.C.; So, K.F.; Chang, R.C. Effects of all-trans-retinoic acid on human SH-SY5Y neuroblastoma as in vitro model in neurotoxicity research. Neurotoxicology 2009, 30, 127–135. [Google Scholar] [CrossRef] [PubMed]
- Presgraves, S.P.; Ahmed, T.; Borwege, S.; Joyce, J.N. Terminally differentiated SH-SY5Y cells provide a model system for studying neuroprotective effects of dopamine agonists. Neurotox. Res. 2004, 5, 579–598. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Dragoni, S.; Franco, G.; Regoli, M.; Bracciali, M.; Morandi, V.; Sgaragli, G.; Bertelli, E.; Valoti, M. Gold nanoparticles uptake and cytotoxicity assessed on rat liver precision-cut slices. Toxicol. Sci. 2012, 128, 186–197. [Google Scholar] [CrossRef] [PubMed]
- De Deurwaerdere, P.; Binda, C.; Corne, R.; Leone, C.; Valeri, A.; Valoti, M.; Ramsay, R.R.; Fall, Y.; Marco-Contelles, J. Comparative Analysis of the Neurochemical Profile and MAO Inhibition Properties of N-(Furan-2-ylmethyl)-N-methylprop-2-yn-1-amine. ACS Chem. Neurosci. 2017, 8, 1026–1035. [Google Scholar] [CrossRef] [PubMed]
- Sylvester, P.W. Optimization of the tetrazolium dye (MTT) colorimetric assay for cellular growth and viability. Methods Mol. Biol. 2011, 716, 157–168. [Google Scholar] [CrossRef] [PubMed]
- Yuan, J.S.; Reed, A.; Chen, F.; Stewart, C.N., Jr. Statistical analysis of real-time PCR data. BMC Bioinform. 2006, 7, 85. [Google Scholar] [CrossRef] [PubMed]

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandez-Abascal, J.; Ripullone, M.; Valeri, A.; Leone, C.; Valoti, M. β-Naphtoflavone and Ethanol Induce Cytochrome P450 and Protect towards MPP+ Toxicity in Human Neuroblastoma SH-SY5Y Cells. Int. J. Mol. Sci. 2018, 19, 3369. https://doi.org/10.3390/ijms19113369
Fernandez-Abascal J, Ripullone M, Valeri A, Leone C, Valoti M. β-Naphtoflavone and Ethanol Induce Cytochrome P450 and Protect towards MPP+ Toxicity in Human Neuroblastoma SH-SY5Y Cells. International Journal of Molecular Sciences. 2018; 19(11):3369. https://doi.org/10.3390/ijms19113369
Chicago/Turabian StyleFernandez-Abascal, Jesus, Mariantonia Ripullone, Aurora Valeri, Cosima Leone, and Massimo Valoti. 2018. "β-Naphtoflavone and Ethanol Induce Cytochrome P450 and Protect towards MPP+ Toxicity in Human Neuroblastoma SH-SY5Y Cells" International Journal of Molecular Sciences 19, no. 11: 3369. https://doi.org/10.3390/ijms19113369