Anti-Inflammatory Activity of Populus deltoides Leaf Extract via Modulating NF-κB and p38/JNK Pathways


1
Department of Medical Science, College of Medical Science, Soonchunhyang University, Asan, Chungnam 31538, Korea
2
Department of Medical Biotechnology, College of Medical Science, Soonchunhyang University, Asan, Chungnam 31538, Korea
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(12), 3746; https://doi.org/10.3390/ijms19123746
Submission received: 20 October 2018
/
Revised: 16 November 2018
/
Accepted: 21 November 2018
/
Published: 25 November 2018
(This article belongs to the Special Issue Natural Anti-Inflammatory Agents 2018)
Abstract
:Populus deltoides, known as eastern cottonwood, has been commonly used as a medicinal plant. The aim of the present study was to investigate the mechanism underlying the anti-inflammatory activity of P. deltoides leaf extract (PLE). PLE effectively inhibited the expression of inducible nitric oxide synthase (iNOS) and nitric oxide (NO) production in lipopolysaccharide (LPS)-stimulated RAW 264.7 cells, but not that of cyclooxygenase-2 (COX-2) and prostaglandin E2. Proinflammatory tumor necrosis factor alpha (TNF-α) levels were also reduced by the extract. PLE inhibited the phosphorylation of nuclear factor-kappa B (NF-κB) and inhibitor of Kappa Bα (IκBα), and blunted LPS-triggered enhanced nuclear translocation of NF-κB p65. In mitogen-activated protein kinase (MAPK) signaling, PLE effectively decreased the phosphorylation of p38 and c-Jun N-terminal protein kinase (JNK), but not of extracellular signal-regulated kinase 1/2 (ERK1/2). Taken together, these results suggest that anti-inflammatory activity of P. deltoides leaf extract might be driven by iNOS and NO inhibition mediated by modulation of the NF-κB and p38/JNK signaling pathways.
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
Inflammation is a complex defense mechanism for an organism against a range of harmful stimuli, including bacteria and parasites. Macrophages are important immune cells responsible for innate cellular immunity involved in host defense and immunity against foreign stimuli [1]. Macrophages stimulated by lipopolysaccharide (LPS), a principal component of the outer membrane of Gram-negative bacteria, regulate inflammation by releasing proinflammatory cytokines, such as tumor necrosis factor alpha (TNF-α), interleukin-6 (IL-6), and adhesion enzymes, such as inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) [1,2,3]. In addition, the mitogen-activated protein kinase (MAPK) and the nuclear factor-kappa B (NF-κB) signaling pathways are important intracellular molecular pathways leading to inflammatory responses to LPS stimulation in macrophages [4,5]. However, imbalanced production of proinflammatory cytokines by macrophages causes destructive inflammation in the body and leads to diseases, such as cancer [6], atherosclerosis [7], diabetic nephropathy [8,9], and neurodegenerative disorders [10].
Nitric oxide (NO) induced by iNOS has been known to be a mediator and regulator of inflammatory reactions that can provoke harmful effects to host tissues. Therefore, inhibiting the production of NO, TNF-α, IL-6, and IL-8 secreted by macrophages through blocking the expression of iNOS can serve as the basis of anti-inflammatory drug development [11,12].
MAPK rapidly responds to a variety of extracellular stimuli and regulates essential cellular events to control a vast array of physiological processes, including macrophage-mediated inflammatory responses [13,14]. The MAPK signaling pathway, involving extracellular signal regulated kinase 1/2 (ERK1/2), c-Jun N-terminal protein kinase (JNK), and p38, is activated by sequential phosphorylation events, and mediates signaling cascades leading to the activation of various transcription factors, such as NF-κB [15,16]. Activation of the inhibitor of Kappa B kinase (IKK) complex, especially IKKβ, results in the breakdown of inhibitor of NF-κB (IκB) following inflammatory stimuli through direct phosphorylation of inhibitor of Kappa Bα (IκBα). NF-κB binds to the IκB family in the form of homo- or heterodimers, and the phosphorylated IκBs are subsequently ubiquitinated and degraded by the proteasome, leaving NF-κB free to translocate to the nucleus. Nuclear NF-κB binds to cognate enhancer/promoter elements of inflammation-related target genes [17,18,19]. Various anti-inflammatory agents targeting the MAPK and NF-κB pathways are being developed on the basis of this cascade response.
A variety of natural products with anti-inflammatory activity have been studied worldwide, including those derived from medicinal plants, such as Allium hookeri [20], Moutan cortex [21], Salvia officinalis [22], Perilla frutescens [23], Camellia japonica [24], and Nauclea officinalis [25]. Moreover, the therapeutic potential of plant secondary metabolites is regarded as a promising research target in the pursuit of novel, natural anti-inflammatory drugs. Some plant-based natural drugs are in clinical use, and some are undergoing clinical trials [26]. A wide variety of chemical components in plants, such as phenolic compounds, flavonoids, coumarins, alkaloids, saponins, sterols, terpenoids, and essential oils, have been known to exhibit anti-inflammatory activities by inhibiting the molecular targets of pro-inflammatory mediators in inflammatory responses. Notably, phenolic glycosides were known to possess a wide variety of bioactivities, including antioxidant, antimicrobial, and anti-inflammatory activities [27,28,29,30].
Populus deltoides, also called eastern cottonwood, is distributed worldwide and has been utilized in traditional medicine to treat various inflammatory conditions [31]. The bark tincture of P. deltoides is known to be effective for treating rheumatism and gout, and its buds have been used to treat colds, respiratory problems, myalgia, and soreness [32]. In addition, various chemical components, including salicortin, salicin, salicyl alcohol, pyrocatechol, 1-O-p-coumaroyl-β-d-glucoside, populoside, ω-salicyloylsalicin, chrysin-7-glucoside, deltoidin, and tremulacin, were identified in P. deltoides [33,34,35,36]. However, information on the utility and efficacy of P. deltoides leaf as an anti-inflammatory agent is limited. In this study, we investigated the anti-inflammatory activity of P. deltoides leaf extract, and dissected the mechanism underlying its anti-inflammatory activity in LPS-induced RAW 264.7 macrophages.
2. Results
2.1. Effect of P. deltoides Leaf Extract on LPS-Stimulated iNOS and NO in RAW 264.7 Cells
Proinflammatory enzyme iNOS produced NO, which has a marked impact on various acute and chronic inflammatory diseases [37]. Figure 1 shows the effect of PLE on the expression of iNOS and COX-2. The LPS-induced expression of iNOS in RAW 264.7 cells was significantly reduced by application of 12.5 and 25 μg/mL of PLE. Conversely, COX-2 expression was not affected by PLE at these concentrations. The LPS-induced iNOS expression was also inhibited by the iNOS inhibitor 1400 W.
Figure 2A shows that PLE significantly reduced LPS-induced NO production. The effect of PLE on the level of TNF-α, a pro-inflammatory cytokine, in LPS-stimulated RAW 264.7 cells was also investigated by ELISA. The level of LPS-induced TNF-α was markedly reduced after application of 12.5 and 25 μg/mL of PLE (Figure 2B). Moreover, iNOS inhibitor 1400W also notably reduced TNF-α expression. Taken together, the results suggest the anti-inflammatory activity of PLE might be exerted by reducing iNOS expression and NO production, not COX-2.
2.2. Effects of P. deltoides Leaf Extract on the NF-κB Signaling in LPS-Stimulated RAW 264.7 Cells
In order to examine whether the NF-κB pathway is involved in the anti-inflammatory effect of PLE, the expression pattern of NF-κB signaling molecules was investigated by western blotting. Figure 3 shows that the phosphorylated IKKα/β, IκBα, and NF-κB p65 were enhanced in LPS-stimulated RAW 264.7 cells. However, the phosphorylation of IKKα/β, IκBα, and NF-κB p65 was significantly reduced upon treatment with PLE. This result suggests that PLE effectively suppresses the NF-κB-associated inflammatory response induced by LPS.
2.3. Effects of P. deltoides Leaf Extract on MAPK Signaling in LPS-Stimulated RAW 264.7 Cells
The mitogen-activated protein kinase (MAPK) family, including ERK1/2, p38, and JNK, play an important role in the regulation of gene expression, cellular growth, and survival through association with the inflammatory response. Figure 5 indicates that the phosphorylation of JNK and p38 was upregulated by LPS, but the phosphorylation of JNK and p38 was markedly down regulated by application of 12.5 and 25 μg/mL of PLE. Interestingly, however, phosphorylation of ERK1/2 was not influenced by PLE application.
3. Discussion
P. deltoides has been widely used as an ancient medicinal herb. However, recent studies on P. deltoides have mainly focused on its response to environmental stressors, including air pollution [39] and water deficits [31], supporting the virtues of P. deltoides as a roadside tree. In this investigation, the mechanism underlying the anti-inflammatory effect of PLE was examined in RAW 264.7 cells stimulated with LPS.
In response to diverse stimulants including microbial infection, immune cells actively release NO, which may lead to chronic or acute inflammatory diseases [40,41]. Our results show that PLE reduced LPS-induced iNOS expression and NO production to a level similar to that of the control. The overproduction of NO is positively linked with excess iNOS expression. Thus, the release of NO was reduced as iNOS expression was blocked [42]. Accordingly, selective inhibitors of iNOS in inflammatory macrophages are considered to be effective therapeutics for inflammatory diseases [43]. In this investigation, PLE was very effective in inhibiting iNOS expression and NO production, showing its superior anti-inflammatory activity. In addition to NO, prostaglandin E2 (PGE2) released from immune cells plays a major role in a variety of inflammatory responses catalyzed by the expression of COX-2 [44]. NO produced by iNOS in the inflammatory response contributes to upregulation of COX-2 via crosstalk between NO, COX-2, and PGE2 [45]. However, PLE did not inhibit the expression COX-2 or prostaglandin E2 in the present study.
The genus Populus, including Populus deltoides, is characterized by the presence of salicin, salicortin [33,46,47], pyrocatechol [48], 1-p-coumaroyl-β-d-glucoside [49], and tremulacin [50]. Salicin is a commonly found phenolic glycoside in the bark, leaves, and buds of more than 100 Populus species [35] with anti-inflammatory, antiangiogenic, and antitumor activities [51,52]. Salicin has been widely used in natural medicines without side effects, such as allergic skin reactions and vomiting [53]. However, salicin itself does not possess anti-inflammatory properties until it is metabolized into salicylic acid in the blood and gastrointestinal tract. Interestingly, some derivatives of salicylic acids showed differential inhibitory effects on COX-2 in murine macrophages [54].
Although PLE might include various potential COX-2 inhibitors, the anti-inflammatory effect of PLE was exerted through iNOS and NO, not COX-2, in this study. This result could be explained by the property of plant extracts. The possession of many constituents might be determined by the presence of synergic interaction and cocktail effect of the extracts. Similar effects were found in the extracts of Acanthopanax sessiliflorum, Daphne genkwa, and Thuja orientalis, strongly inhibiting iNOS activity, but weakly inhibiting COX-2 activity [55]. Generally, plant extracts as a mixture of phytochemical components have been suspected to have higher activities than those obtained with single isolated components. In some cases, the activity of a mixture of pure compounds was not higher than that of a single compound [56]. Therefore, minor constituents of plant extracts may play a major role in expressing specific activity of the extracts.
TNF-α is crucial for inducing synergy in NO production in LPS-stimulated macrophages and induces inflammatory responses such as vasodilatation, edema, and fever [57,58]. In this study, PLE significantly inhibited the levels of TNF-α. The MAPK and NF-κB signaling pathways are known to be important in regulating the expression of proinflammatory enzymes and cytokines. NF-κB is a major transcription factor involved in the regulation of proinflammatory cytokines, such as p50, p65, and IκBα [59,60]. In this investigation, the inhibition of p65 and IκBα phosphorylation, along with nuclear translocation by PLE, demonstrated that the inflammatory response of PLE is associated with NF-κB signaling.
The MAPK family of proteins, which consist of p38, JNK, and ERK1/2, positively affect NF-κB activation [61]. Interestingly, we confirmed by western blot that PLE significantly inhibited p38 and JNK phosphorylation dose-dependently, while ERK1/2 phosphorylation was not significantly affected. MAPK kinase (MAPKK), the upstream effector that activates MAPK, is composed of MEK 1-7 [62,63]. MEK1 and MEK2, the upstream regulators of ERK1/2, were not affected by PLE (data not shown). Similar anti-inflammatory properties were reported in sophocarpine from Sophora alopecuroides, angelicin from Psoralea corylifolia L. fruit, and the sulfated derivative of 20(s)-ginsenoside Rh2 from red ginseng, which inhibit p38 and JNK phosphorylation, but not ERK1/2 [12,59,62].
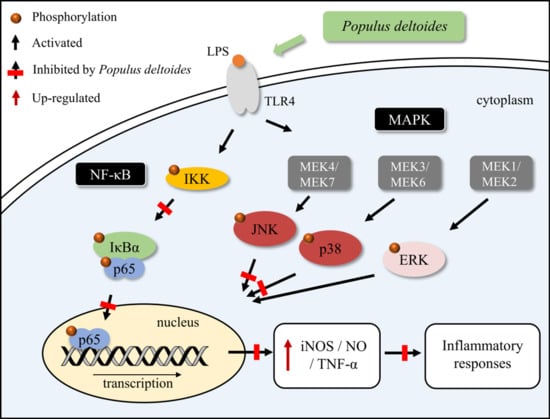
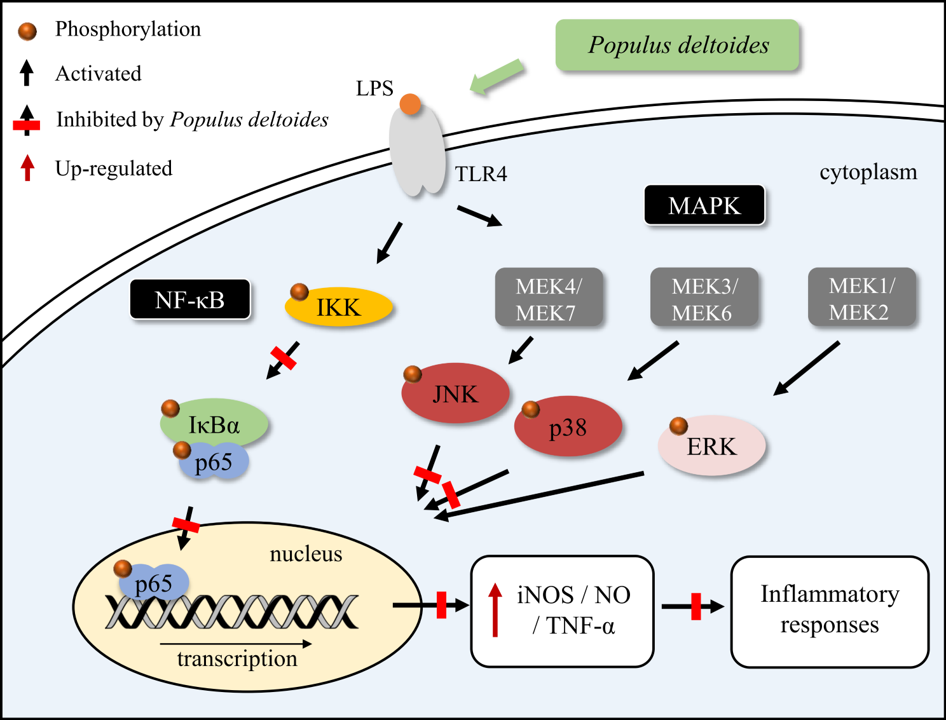
Our data demonstrated that P. deltoides leaf extract exerts an anti-inflammatory effect, which might be attributed to the inhibition of iNOS expression, via suppression of the JNK and p38 MAPK signaling and the inhibition of NF-κB activation (Figure 6).
4. Materials and Methods
4.1. Preparation of Methanol Extract of P. deltoides
The leaf extract of P. deltoides was obtained from the Plant Extract Bank at the Korea Research Institute of Bioscience and Biotechnology (KRIBB, Daejeon, Republic of Korea) and was extracted with 99.9% pure methanol. The extract was dissolved in dimethyl sulfoxide to be 2% and stored at −20 °C until needed. The doses of the extract (12.5 or 25 μg/mL) used here were the highest concentrations that did not show any toxicities on the LPS-stimulated RAW 264.7 cells.
4.2. RAW 264.7 Cell Culture and Cell Viability Assay
RAW 264.7 cells, a murine macrophage cell line, were maintained in Dulbecco’s modified Eagle’s medium (DMEM; Hyclone, Logan, UT, USA) [with 10% fetal bovine serum (FBS; Hyclone, Logan, UT, USA) and 1% penicillin-streptomycin (PS; Hyclone, Logan, UT, USA)] in a humidified atmosphere containing 5% CO2 at 37 °C. Cells were stimulated by replacing the culture medium with medium containing 1 μg/mL LPS (Escherichia coli 011:B4, Sigma Chemical Co., St. Louis, MO, USA), and incubated with P. deltoides leaf extract for 24 h.
Cell viability was assessed by 3-(4,5-dimethyl-thiazol-2-yl)-2,5-diphenyltetrazolium bromide (MTT) assay. Briefly, 4 × 104 RAW 264.7 cells per well of a 96-well plate were incubated with various concentrations of P. deltoides leaf extract (0–200 μg/mL) at 37 °C for 24 h and then the medium was completely removed. The cells were washed and treated with 50 μL of MTT, after which the plates were incubated at 37 °C in the dark for 3 h. The resultant formazan crystals were dissolved in 50 μL of dimethyl sulfoxide, and the absorbance was measured at 570 nm using an ELISA reader (Tecan, Männedorf, Switzerland).
4.3. NO Production
The nitrite content in culture medium was measured as an indicator of nitric oxide (NO) production, according to the Griess reaction. Each supernatant (100 μL) was mixed with the same volume of Griess reagent (1% sulfanilamide in 5% phosphoric acid and 0.1% naphthylethylenediamine dihydrochloride in distilled water); the absorbance of the mixture at 570 nm was determined with an ELISA reader.
4.4. ELISA
The effect of PLE on TNF-α production in LPS-stimulated RAW 264.7 cells was measured by ELISA. TNF-α in the culture supernatant was quantified with an OptEIA mouse TNF-α (Mono/Mono) set (BD Biosciences, San Jose, CA, USA). The TNF-α level was calculated based on the standard curve, using the standard in the mouse TNF (Mono/Mono) set.
4.5. Western Blotting
Proteins were separated by 10% SDS–PAGE, then transferred onto polyvinylidene fluoride membrane (Bio-Rad, Hercules, CA, USA). After blocking for 2 h at room temperature with 5% bovine serum albumin (BSA), the membranes were incubated overnight at 4 °C with primary antibodies (iNOS, COX-2, IKKβ, p-IKKα/β, IκBα, p-IκBα, NF-κB p65, p-NF-κB p65, p38, p-p38, ERK1/2, p-ERK1/2, JNK, p-JNK (Cell Signaling Technology, Inc., MA, USA), and β-actin (Santa Cruz Biotechnology, Inc., Dallas, TX, USA)), which were diluted following the manufacturer’s recommendations. The membranes were then washed in Tris-buffered saline Tween 20 (TBST) and incubated with the appropriate HRP-conjugated secondary antibody (1:5000) at room temperature for 1 h. Protein bands were visualized using the Sensi-Q 2000 (Lugen Sci, Bucheon, Korea). The intensity of the bands was analyzed using ImageJ and normalized to β-actin.
4.6. Confocal Microscope Analysis
LPS-stimulated RAW 264.7 cells were treated with 12.5 or 25 μg/mL of P. deltoides leaf extract, after which the cells were fixed with 4% paraformaldehyde in phosphate-buffered saline (PBS) for 10 min. The cells were washed 3 times with PBS containing 0.5 mM MgCl2 and 1 mM CaCl2 for 5 min. After permeabilization with 0.1% Triton X-100 for 10 min, samples were incubated with blocking buffer (5% BSA in PBS). The cells were incubated with primary antibodies (p-NF-κB p65) (1:200) in 1% BSA for overnight at 4 °C. The cells were washed 3 times in PBS for 5 min and were stained for another 3 h with goat anti-rabbit IgG Texas red (1:1000) (Santa Cruz Biotechnology, Dallas, TX, USA). Cells were counterstained with 4, 6-diamidino-2-phenylindole dihydrochloride (DAPI) (Bio-Rad, Hercules, CA, USA) for 10 min to label the nuclei. The prepared cells were then observed under a confocal microscope (Olympus FV10i, Olympus, Tokyo, Japan), and images were recorded.
4.7. Statistical Analysis
Data were entered into a Microsoft Excel datasheet, then transferred into the IBM Statistical Package for Social Sciences (SPSS, version 20, IBM SPSS statistics, Chicago, IL, USA). Data from each group are expressed as mean ± standard deviation (SD) of triplicate experiments. Group differences were analyzed using one-way analysis of variance (ANOVA), followed by a Bonferroni test and p-values less than 0.05 were considered statistically significant.
Author Contributions
Investigation, Y.E.J.; Supervision, M.-Y.L.; Writing—original draft, Y.E.J.; Writing—review & editing, M.-Y.L.; all authors approved the final draft.
Funding
This research received no external funding.
Acknowledgments
This study was supported by Soonchunhyang University.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Zhang, L.; Wang, C.-C. Inflammatory response of macrophages in infection. HBPD Int. 2014, 13, 138–152. [Google Scholar] [CrossRef]
- Guha, M.; Mackman, N. LPS induction of gene expression in human monocytes. Cell. Signal. 2001, 13, 85–94. [Google Scholar] [CrossRef]
- Tsai, S.C.; Liang, Y.H.; Chiang, J.H.; Liu, F.C.; Lin, W.H.; Chang, S.J.; Lin, W.Y.; Wu, C.H.; Weng, J.R. Anti-inflammatory effects of Calophyllum inophyllum L. in RAW264.7 cells. Oncol. Rep. 2012, 28, 1096–1102. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.; Cai, W.; Wang, X.; Liu, Y.; Hou, B.; Zhu, X.; Qiu, L. Vaccaria hypaphorine alleviates lipopolysaccharide-induced inflammation via inactivation of NFκB and ERK pathways in Raw 264.7 cells. BMC Complement. Altern. Med. 2017, 17. [Google Scholar] [CrossRef] [PubMed]
- Ivashkiv, L.B. Inflammatory signaling in macrophages: Transitions from acute to tolerant and alternative activation states. Eur. J. Immunol. 2011, 41, 2477–2481. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biswas, S.K.; Allavena, P.; Mantovani, A. Tumor-associated macrophages: Functional diversity, clinical significance, and open questions. Semin. Immunopathol. 2013, 35, 585–600. [Google Scholar] [CrossRef] [PubMed]
- Moore, K.J.; Sheedy, F.J.; Fisher, E.A. Macrophages in atherosclerosis: A dynamic balance. Nat. Rev. Immunol. 2013, 13, 709–721. [Google Scholar] [CrossRef] [PubMed]
- Navarro-Gonzalez, J.F.; Mora-Fernandez, C. The role of inflammatory cytokines in diabetic nephropathy. J. Am. Soc. Nephrol. 2008, 19, 433–442. [Google Scholar] [CrossRef] [PubMed]
- Donate-Correa, J.; Martin-Nunez, E.; Muros-de-Fuentes, M.; Mora-Fernandez, C.; Navarro-Gonzalez, J.F. Inflammatory cytokines in diabetic nephropathy. J. Diabetes. Res. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.A.; Das, A.; Ray, S.K.; Banik, N.L. Role of pro-inflammatory cytokines released from microglia in neurodegenerative diseases. Brain Res. Bull. 2012, 87, 10–20. [Google Scholar] [CrossRef] [PubMed]
- Korhonen, R.; Lahti, A.; Kankaanranta, H.; Moilanen, E. Nitric oxide production and signaling in inflammation. Curr. Drug. Targets. Inflamm. Allergy. 2005, 4, 471–479. [Google Scholar] [CrossRef] [PubMed]
- Gao, Y.; Jiang, W.; Dong, C.; Li, C.; Fu, X.; Min, L.; Tian, J.; Jin, H.; Shen, J. Anti-inflammatory effects of sophocarpine in LPS-induced RAW 264.7 cells via NF-κB and MAPKs signaling pathways. Toxicol. In Vitro 2012, 26, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Kim, E.K.; Choi, E.J. Compromised MAPK signaling in human diseases: An update. Arch. Toxicol. 2015, 89, 867–882. [Google Scholar] [CrossRef] [PubMed]
- Neacsu, P.; Mazare, A.; Schmuki, P.; Cimpean, A. Attenuation of the macrophage inflammatory activity byTiO2 nanotubes via inhibition of MAPK and NF-κB pathways. Int. J. Nanomedicine 2015, 10, 6455–6467. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.Y.; Ryu, A.R.; Jin, S.; Jeon, Y.M.; Lee, M.Y. Chlorin e6-mediated photodynamic therapy suppresses P. acnes-induced inflammatory response via NFκB and MAPKs signaling pathway. PLoS ONE 2017, 12, e0170599. [Google Scholar] [CrossRef] [PubMed]
- Limtrakul, P.; Yodkeeree, S.; Pitchakarn, P.; Punfa, W. Anti-inflammatory effects of proanthocyanidin-rich red rice extract via suppression of MAPK, AP-1 and NF-κB pathways in Raw 264.7 macrophages. Nutr. Res. Pract. 2016, 10, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Tripathi, P.; Aggarwal, A. NF-kB transcription factor: A key player in the generation of immune response. Curr. Sci. 2006, 90, 519–531. [Google Scholar]
- Doerks, T.; Copley, R.R.; Schultz, J.; Ponting, C.P.; Bork, P. Systematic identification of novel protein domain families associated with nuclear functions. Genome. Res. 2002, 12, 47–56. [Google Scholar] [CrossRef] [PubMed]
- Perkins, N.D. Integrating cell-signalling pathways with NF-κB and IKK function. Nat. Rev. Mol. Cell. Biol. 2007, 8, 49–62. [Google Scholar] [CrossRef] [PubMed]
- Jang, J.Y.; Lee, M.J.; You, B.R.; Jin, J.S.; Lee, S.H.; Yun, Y.R.; Kim, H.J. Allium hookeri root extract exerts anti-inflammatory effects by nuclear factor-κB down-regulation in lipopolysaccharide-induced RAW264.7 cells. BMC Complement. Altern. Med. 2017, 17. [Google Scholar] [CrossRef]
- Chun, S.C.; Jee, S.Y.; Lee, S.G.; Park, S.J.; Lee, J.R.; Kim, S.C. Anti-inflammatory activity of the methanol extract of moutan cortex in LPS-activated Raw264.7 cells. Evid. Based Complement. Alternat. Med. 2007, 4, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Kolac, U.K.; Ustuner, M.C.; Tekin, N.; Ustuner, D.; Colak, E.; Entok, E. The anti-inflammatory and antioxidant effects of Salvia officinalis on lipopolysaccharide-induced inflammation in rats. J. Med. Food 2017, 20, 1193–1200. [Google Scholar] [CrossRef] [PubMed]
- Huang, B.P.; Lin, C.H.; Chen, Y.C.; Kao, S.H. Anti-inflammatory effects of Perilla frutescens leaf extract on lipopolysaccharide-stimulated RAW264.7 cells. Mol. Med. Rep. 2014, 10, 1077–1083. [Google Scholar] [CrossRef] [PubMed]
- Akanda, M.R.; Park, B.Y. Involvement of MAPK/NF-κB signal transduction pathways: Camellia japonica mitigates inflammation and gastric ulcer. Biomed. Pharmacother. 2017, 95, 1139–1146. [Google Scholar] [CrossRef] [PubMed]
- Zhai, X.T.; Zhang, Z.Y.; Jiang, C.H.; Chen, J.Q.; Ye, J.Q.; Jia, X.B.; Yang, Y.; Ni, Q.; Wang, S.X.; Song, J.; et al. Nauclea officinalis inhibits inflammation in LPS-mediated RAW 264.7 macrophages by suppressing the NF-κB signaling pathway. J. Ethnopharmacol. 2016, 183, 159–165. [Google Scholar] [CrossRef] [PubMed]
- Bernstein, N.; Akram, M.; Daniyal, M.; Koltai, H.; Fridlender, M.; Gorelick, J. Antiinflammatory potential of medicinal plants: A source for therapeutic secondary metabolites. Adv. Agron. 2018, 150, 131–183. [Google Scholar] [CrossRef]
- Boeckler, G.A.; Gershenzon, J.; Unsicker, S.B. Phenolic glycosides of the Salicaceae and their role as anti-herbivore defenses. Phytochemistry 2011, 72, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Thuong, P.T.; Min, B.-S.; Ngoc, T.M.; Hung, T.M.; Lee, I.S.; Na, M.; Seong, Y.-H.; Song, K.-S.; Bae, K. Phenolic glycosides with antioxidant activity from the stem bark of Populus davidiana. J. Nat. Prod. 2006, 69, 1370–1373. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.J.; Huang, H.T.; Huang, S.Y.; Lin, Z.H.; Shen, C.C.; Tsai, W.J.; Kuo, Y.H. Antioxidant and anti-inflammatory phenolic glycosides from Clematis tashiroi. J. Nat. Prod. 2015, 78, 1586–1592. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Wang, H.; Gong, Z.; Huang, J.; Pei, W.; Wang, X.; Zhang, J.; Tang, X. Antimicrobial and cytotoxic phenolics and phenolic glycosides from Sargentodoxa cuneata. Fitoterapia 2015, 101, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Abraham, P.E.; Garcia, B.J.; Gunter, L.E.; Jawdy, S.S.; Engle, N.; Yang, X.; Jacobson, D.A.; Hettich, R.L.; Tuskan, G.A.; Tschaplinski, T.J. Quantitative proteome profile of water deficit stress responses in eastern cottonwood (Populus deltoides) leaves. PLoS ONE 2018, 13, e0190019. [Google Scholar] [CrossRef] [PubMed]
- Duke, J.A. Amerindian medicinal plants; Typescript: Fulton, ML, USA, 1983. [Google Scholar]
- Pearl, I.A.; Darling, S.F. Studies of the hot water extractives of the bark and leaves of Populus deltoides bartr. Can. J. Chem. 1971, 49, 49–55. [Google Scholar] [CrossRef]
- Picard, S.; Chenault, J.; Augustin, S.; Venot, C. Isolation of a new phenolic compound from leaves of Populus deltoides. J. Nat. Prod. 1994, 57, 808–810. [Google Scholar] [CrossRef]
- Clausen, T.P.; Reichardt, P.B.; Bryant, J.P.; Werner, R.A.; Post, K.; Frisby, K. Chemical model for short-term induction in quaking aspen (Populus tremuloides) foliage against herbivores. J. Chem. Ecol. 1989, 15, 2335–2346. [Google Scholar] [CrossRef] [PubMed]
- Snyder, D.T.; Schilling, M.C.; Hochwender, C.G.; Kaufman, A.D. Profiling phenolic glycosides in Populus deltoides and Populus grandidentata by leaf spray ionization tandem mass spectrometry. Anal. Methods 2015, 7, 870–876. [Google Scholar] [CrossRef]
- Kanwar, J.R.; Kanwar, R.K.; Burrow, H.; Baratchi, S. Recent advances on the roles of NO in cancer and chronic inflammatory disorders. Curr. Med. Chem. 2009, 16, 2373–2394. [Google Scholar] [CrossRef] [PubMed]
- Basak, S.; Hoffmann, A. Crosstalk via the NF-κB signaling system. Cytokine Growth Factor Rev. 2008, 19, 187–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guerra, F.; Duplessis, S.; Kohler, A.; Martin, F.; Tapia, J.; Lebed, P.; Zamudio, F.; González, E. Gene expression analysis of Populus deltoides roots subjected to copper stress. Environ. Exp. Bot. 2009, 67, 335–344. [Google Scholar] [CrossRef]
- Uslu, C.; Karasen, R.M.; Sahin, F.; Taysi, S.; Akcay, F. Effects of aqueous extracts of Satureja hortensis L. On rhinosinusitis treatment in rabbit. J. Ethnopharmacol. 2003, 88, 225–228. [Google Scholar] [CrossRef]
- Bogdan, C.; Röllinghoff, M.; Diefenbach, A. Reactive oxygen and reactive nitrogen intermediates in innate and specific immunity. Curr. Opin. Immunol. 2000, 12, 64–76. [Google Scholar] [CrossRef]
- Baatar, D.; Siddiqi, M.Z.; Im, W.T.; Ul Khaliq, N.; Hwang, S.G. Anti-inflammatory effect of Ginsenoside Rh2-mix on lipopolysaccharide-stimulated RAW 264.7 murine macrophage cells. J. Med. Food 2018, 21, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Wen, Z.S.; Xiang, X.W.; Jin, H.X.; Guo, X.Y.; Liu, L.J.; Huang, Y.N.; OuYang, X.K.; Qu, Y.L. Composition and anti-inflammatory effect of polysaccharides from Sargassum horneri in RAW264.7 macrophages. Int. J. Biol. Macromol. 2016, 88, 403–413. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Leite, C.; Nunes, C.; Jamal, S.K.; Cuccovia, I.M.; Reis, S. Nonsteroidal anti-inflammatory therapy: A journey toward safety. Med. Res. Rev. 2017, 37, 802–859. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, Y.; Sobajima, J.; Higashi, M.; Ishiguro, T.; Fukuchi, M.; Ishibashi, K.; Mochiki, E.; Yakabi, K.; Kawano, T.; Tamaru, J.; et al. Coexpression of COX-2 and iNOS in angiogenesis of superficial esophageal squamous cell carcinoma. Int. Surg. 2015, 100, 733–743. [Google Scholar] [CrossRef] [PubMed]
- Thieme, H. ber die phenolglykoside der gattung Populus. Planta Medica 1967, 15, 35–40. [Google Scholar] [CrossRef] [PubMed]
- Zhang, C.-p.; Zheng, H.-q.; Liu, G.; Hu, F.-l. Development and validation of HPLC method for determination of salicin in poplar buds: Application for screening of counterfeit propolis. Food Chem. 2011, 127, 345–350. [Google Scholar] [CrossRef]
- Chang, M.-C.; Chang, H.-H.; Wang, T.-M.; Chan, C.-P.; Lin, B.-R.; Yeung, S.-Y.; Yeh, C.-Y.; Cheng, R.-H.; Jeng, J.-H. Antiplatelet effect of catechol is related to inhibition of cyclooxygenase, reactive oxygen species, ERK/p38 signaling and thromboxane A2 production. PLoS ONE 2014, 9, e104310. [Google Scholar] [CrossRef] [PubMed]
- Vo, V.A.; Lee, J.-W.; Kim, J.-Y.; Park, J.-H.; Lee, H.J.; Kim, S.-S.; Kwon, Y.-S.; Chun, W. Phosphorylation of Akt mediates anti-inflammatory activity of 1-p-coumaroyl β-D-glucoside against lipopolysaccharide-induced inflammation in RAW264. 7 cells. Korean. J. Physiol. Pharmacol. 2014, 18, 79–86. [Google Scholar] [CrossRef] [PubMed]
- Cheng, G.; Liu, D.; Yang, D.; He, K.; Bai, J.; Zhu, X. Antiinflammatory effects of tremulacin, a salicin-related substance isolated from Populus tomentosa carr. leaves. Phytomedicine 1994, 1, 209–211. [Google Scholar] [CrossRef]
- Li, Y.; Wu, Q.; Deng, Y.; Lv, H.; Qiu, J.; Chi, G.; Feng, H.D. (−)-salicin inhibits the LPS-induced inflammation in RAW264.7 cells and mouse models. Int. Immunopharmacol. 2015, 26, 286–294. [Google Scholar] [CrossRef] [PubMed]
- Kong, C.S.; Kim, K.H.; Choi, J.S.; Kim, J.E.; Park, C.; Jeong, J.W. Salicin, an extract from white willow bark, inhibits angiogenesis by blocking the ROS-ERK pathways. Phytother. Res. 2014, 28, 1246–1251. [Google Scholar] [CrossRef] [PubMed]
- Wick, J. Aspirin: A history, a love story. Consult. Pharm. 2012, 27, 322–329. [Google Scholar] [CrossRef] [PubMed]
- Hinz, B.; Kraus, V.; Pahl, A.; Brune, K. Salicylate metabolites inhibit cyclooxygenase-2-dependent prostaglandin E2 synthesis in murine macrophages. Biochem. Biophys. Res. Commun. 2000, 274, 197–202. [Google Scholar] [CrossRef] [PubMed]
- Hong, C.H.; Hur, S.K.; Oh, O.-J.; Kim, S.S.; Nam, K.A.; Lee, S.K. Evaluation of natural products on inhibition of inducible cyclooxygenase (COX-2) and nitric oxide synthase (iNOS) in cultured mouse macrophage cells. J. Ethnopharmacol. 2002, 83, 153–159. [Google Scholar] [CrossRef]
- Muturi, E.J.; Ramirez, J.L.; Doll, K.M.; Bowman, M.J. Combined toxicity of three essential oils against Aedes aegypti (Diptera: Culicidae) larvae. J. Med. Entomol. 2017, 54, 1684–1691. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.-B.; Han, A.-R.; Park, E.-Y.; Kim, J.-Y.; Cho, W.; Lee, J.; Seo, E.-K.; Lee, K.-T. Inhibition of LPS-induced iNOS, COX-2 and cytokines expression by poncirin through the NF-κB inactivation in RAW 264.7 macrophage cells. Biol. Pharm. Bull. 2007, 30, 2345–2351. [Google Scholar] [CrossRef] [PubMed]
- Zelova, H.; Hosek, J. TNF-α signalling and inflammation: Interactions between old acquaintances. Inflamm. Res. 2013, 62, 641–651. [Google Scholar] [CrossRef] [PubMed]
- Liu, F.; Sun, G.Q.; Gao, H.Y.; Li, R.S.; Soromou, L.W.; Chen, N.; Deng, Y.H.; Feng, H.H. Angelicin regulates LPS-induced inflammation via inhibiting MAPK/NF-κB pathways. J. Surg. Res. 2013, 185, 300–309. [Google Scholar] [CrossRef] [PubMed]
- Kuo, M.Y.; Liao, M.F.; Chen, F.L.; Li, Y.C.; Yang, M.L.; Lin, R.H.; Kuan, Y.H. Luteolin attenuates the pulmonary inflammatory response involves abilities of antioxidation and inhibition of MAPK and NFκB pathways in mice with endotoxin-induced acute lung injury. Food Chem. Toxicol. 2011, 49, 2660–2666. [Google Scholar] [CrossRef] [PubMed]
- Choi, R.J.; Chun, J.; Khan, S.; Kim, Y.S. Desoxyrhapontigenin, a potent anti-inflammatory phytochemical, inhibits LPS-induced inflammatory responses via suppressing NF-κB and MAPK pathways in RAW 264.7 cells. Int. Immunopharmacol. 2014, 18, 182–190. [Google Scholar] [CrossRef] [PubMed]
- Bi, W.Y.; Fu, B.D.; Shen, H.Q.; Wei, Q.; Zhang, C.; Song, Z.; Qin, Q.Q.; Li, H.P.; Lv, S.; Wu, S.C.; et al. Sulfated derivative of 20(s)-ginsenoside Rh2 inhibits inflammatory cytokines through MAPKs and NF-kappa B pathways in LPS-induced RAW264.7 macrophages. Inflammation 2012, 35, 1659–1668. [Google Scholar] [CrossRef] [PubMed]
- Gerits, N.; Kostenko, S.; Moens, U. In vivo functions of mitogen-activated protein kinases: Conclusions from knock-in and knock-out mice. Transgenic Res. 2007, 16, 281–314. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Effects of P. deltoides leaf extract on inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) expression in LPS-stimulated RAW 264.7 cells. The cells were treated with P. deltoides leaf extract (12.5 and 25 μg/mL) in the presence of 1 μg/mL LPS for 24 h. RAW 264.7 cells were treated with 10 μM of iNOS inhibitor 1400 W simultaneously with LPS treatment. Error bars represent the mean ± SD. *** p < 0.001 compared to LPS alone.
Figure 1.
Effects of P. deltoides leaf extract on inducible nitric oxide synthase (iNOS) and cyclooxygenase-2 (COX-2) expression in LPS-stimulated RAW 264.7 cells. The cells were treated with P. deltoides leaf extract (12.5 and 25 μg/mL) in the presence of 1 μg/mL LPS for 24 h. RAW 264.7 cells were treated with 10 μM of iNOS inhibitor 1400 W simultaneously with LPS treatment. Error bars represent the mean ± SD. *** p < 0.001 compared to LPS alone.

Figure 2.
Effect of P. deltoides leaf extract on the production of nitric oxide (NO) and tumor necrosis factor alpha (TNF-α) in LPS-stimulated RAW 264.7 cells. P. deltoides leaf extract (12.5 or 25 μg/mL) suppressed (A) NO production and (B) TNF-α expression in RAW 264.7 cells stimulated by 1 μg/mL LPS. The expressions of TNF-α were also down regulated by treatment of iNOS inhibitor 1400 W (10 μM). Error bars represent the mean ± SD. * p < 0.05 and *** p < 0.001 compared to LPS alone.
Figure 2.
Effect of P. deltoides leaf extract on the production of nitric oxide (NO) and tumor necrosis factor alpha (TNF-α) in LPS-stimulated RAW 264.7 cells. P. deltoides leaf extract (12.5 or 25 μg/mL) suppressed (A) NO production and (B) TNF-α expression in RAW 264.7 cells stimulated by 1 μg/mL LPS. The expressions of TNF-α were also down regulated by treatment of iNOS inhibitor 1400 W (10 μM). Error bars represent the mean ± SD. * p < 0.05 and *** p < 0.001 compared to LPS alone.

Figure 3.
Effects of P. deltoides leaf extract on the LPS-induced activation of the nuclear factor-kappa (NF-κB) pathway in RAW 264.7 cells. The cells were treated with P. deltoides leaf extract (12.5 or 25 μg/mL) in the presence of 1 μg/mL LPS for 24 h. Error bars represent the mean ± SD. Values of * p < 0.05 and *** p < 0.001 compared to LPS alone were considered statistically significant.
Figure 3.
Effects of P. deltoides leaf extract on the LPS-induced activation of the nuclear factor-kappa (NF-κB) pathway in RAW 264.7 cells. The cells were treated with P. deltoides leaf extract (12.5 or 25 μg/mL) in the presence of 1 μg/mL LPS for 24 h. Error bars represent the mean ± SD. Values of * p < 0.05 and *** p < 0.001 compared to LPS alone were considered statistically significant.

Figure 4.
The inhibitory effect of P. deltoides leaf extract on NF-κB p65 translocation in LPS-stimulated RAW 264.7 cells. Representative photomicrographs of immune-labelled NF-κB (red) and nuclear counterstaining with DAPI (blue) in LPS-stimulated RAW 264.7 cells treated without and with 12.5 or 25 μg/mL of PLE. LPS increased colocalization of NF-κB and DAPI in RAW 264.7 cells. However, PLE reduced the nuclear translocation and accumulation of NF-κB, which was induced by LPS. Scale bar = 10 μm.
Figure 4.
The inhibitory effect of P. deltoides leaf extract on NF-κB p65 translocation in LPS-stimulated RAW 264.7 cells. Representative photomicrographs of immune-labelled NF-κB (red) and nuclear counterstaining with DAPI (blue) in LPS-stimulated RAW 264.7 cells treated without and with 12.5 or 25 μg/mL of PLE. LPS increased colocalization of NF-κB and DAPI in RAW 264.7 cells. However, PLE reduced the nuclear translocation and accumulation of NF-κB, which was induced by LPS. Scale bar = 10 μm.
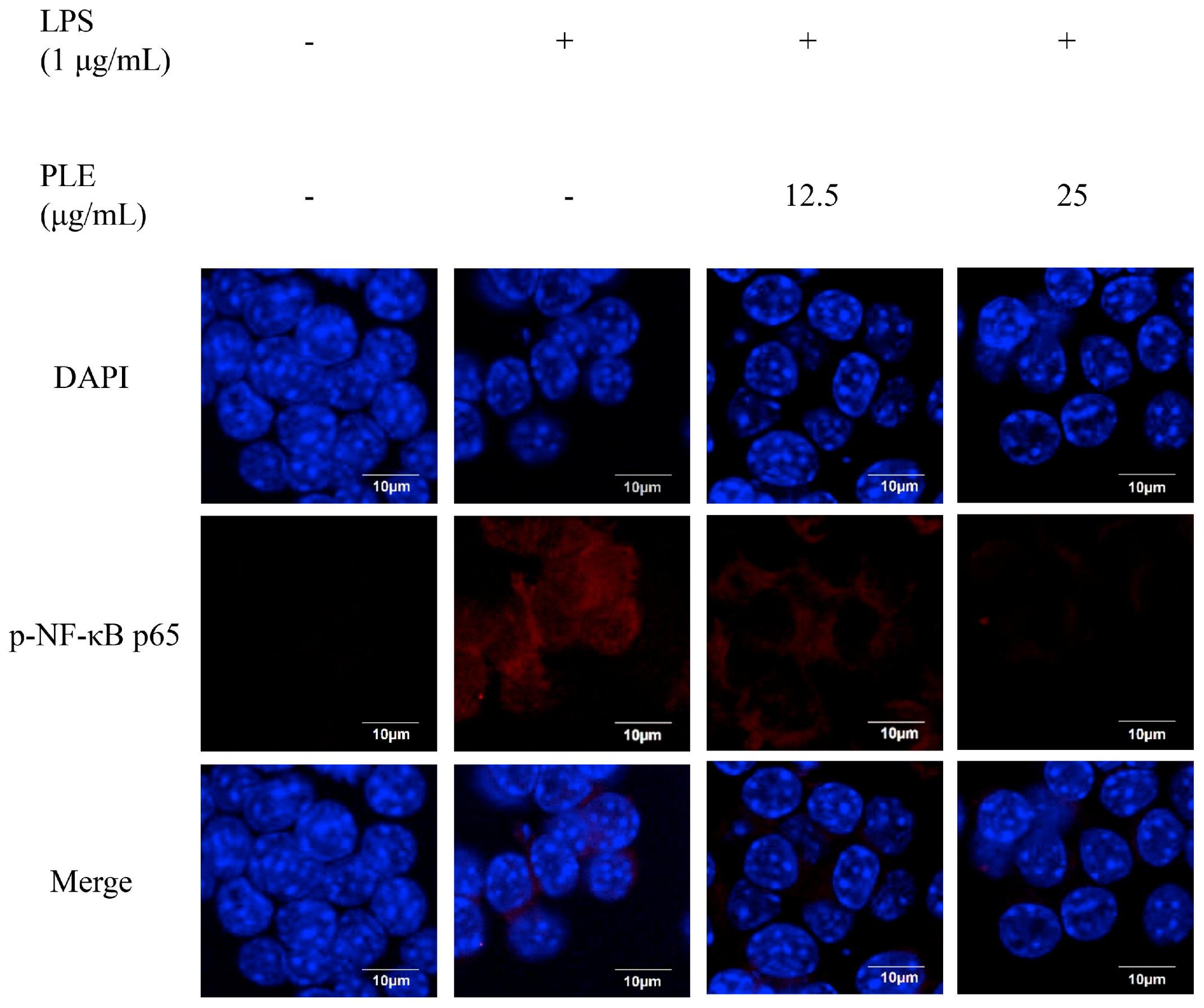
Figure 5.
Effects of PLE on the phosphorylation of mitogen-activated protein kinase (MAPK) cascade (p-ERK1/2, p-JNK, and p-p38) in LPS-stimulated RAW 264.7 cells. The cells were treated with P. deltoides leaf extract (12.5 or 25 μg/mL) in the presence of 1 μg/mL LPS. Error bars represent the mean ± SD. Values of ** p < 0.01, and *** p < 0.001 compared to LPS alone.
Figure 5.
Effects of PLE on the phosphorylation of mitogen-activated protein kinase (MAPK) cascade (p-ERK1/2, p-JNK, and p-p38) in LPS-stimulated RAW 264.7 cells. The cells were treated with P. deltoides leaf extract (12.5 or 25 μg/mL) in the presence of 1 μg/mL LPS. Error bars represent the mean ± SD. Values of ** p < 0.01, and *** p < 0.001 compared to LPS alone.

Figure 6.
Schematic diagram of the suppressive effects of P. deltoides leaf extract on the inflammatory response via modulating NF-κB and p38/JNK pathway in LPS-stimulated RAW 264.7 cells.
Figure 6.
Schematic diagram of the suppressive effects of P. deltoides leaf extract on the inflammatory response via modulating NF-κB and p38/JNK pathway in LPS-stimulated RAW 264.7 cells.

© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Jeong, Y.E.; Lee, M.-Y. Anti-Inflammatory Activity of Populus deltoides Leaf Extract via Modulating NF-κB and p38/JNK Pathways. Int. J. Mol. Sci. 2018, 19, 3746. https://doi.org/10.3390/ijms19123746
AMA Style
Jeong YE, Lee M-Y. Anti-Inflammatory Activity of Populus deltoides Leaf Extract via Modulating NF-κB and p38/JNK Pathways. International Journal of Molecular Sciences. 2018; 19(12):3746. https://doi.org/10.3390/ijms19123746
Chicago/Turabian StyleJeong, Ye Eun, and Mi-Young Lee. 2018. "Anti-Inflammatory Activity of Populus deltoides Leaf Extract via Modulating NF-κB and p38/JNK Pathways" International Journal of Molecular Sciences 19, no. 12: 3746. https://doi.org/10.3390/ijms19123746
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.