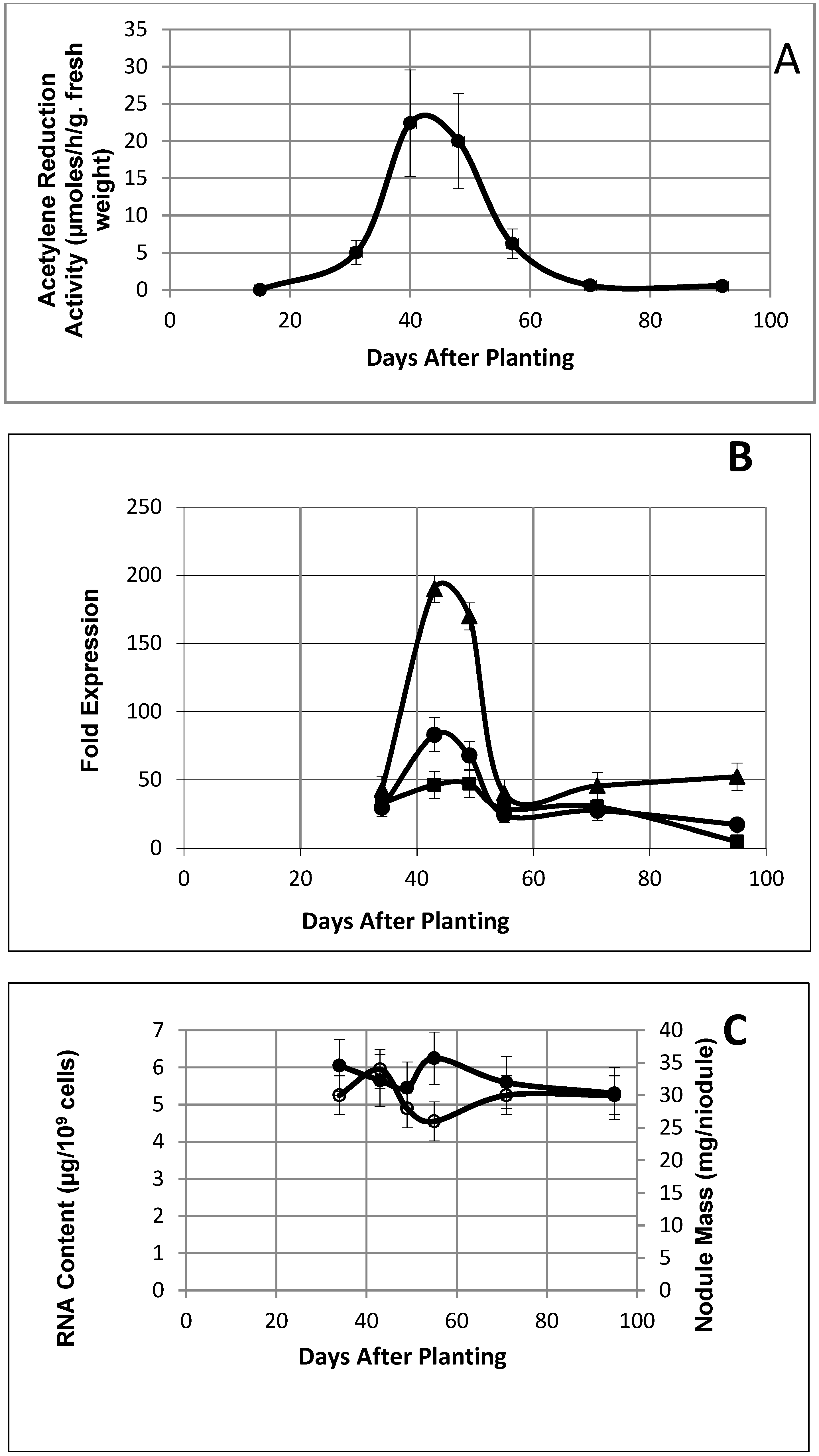
Figure 1.
(A) Acetylene reduction activity of soybean nodules as a function of days after planting. Values are the means of three replicates. (B) Nitrogenase component gene expression from microarray data as a function of days after planting. Symbols represent the molybdenum–iron protein, alpha subunit (●); molybdenum–iron protein, beta subunit (■); iron protein (▲). Each value is the average of three samples ± S.D. (C) Total RNA isolated from soybean nodule bacteroids and soybean nodule mass as a function of days after planting. Values are means of at least three replicates. Solid symbols are RNA content; open symbols are nodule mass.
Figure 1.
(A) Acetylene reduction activity of soybean nodules as a function of days after planting. Values are the means of three replicates. (B) Nitrogenase component gene expression from microarray data as a function of days after planting. Symbols represent the molybdenum–iron protein, alpha subunit (●); molybdenum–iron protein, beta subunit (■); iron protein (▲). Each value is the average of three samples ± S.D. (C) Total RNA isolated from soybean nodule bacteroids and soybean nodule mass as a function of days after planting. Values are means of at least three replicates. Solid symbols are RNA content; open symbols are nodule mass.
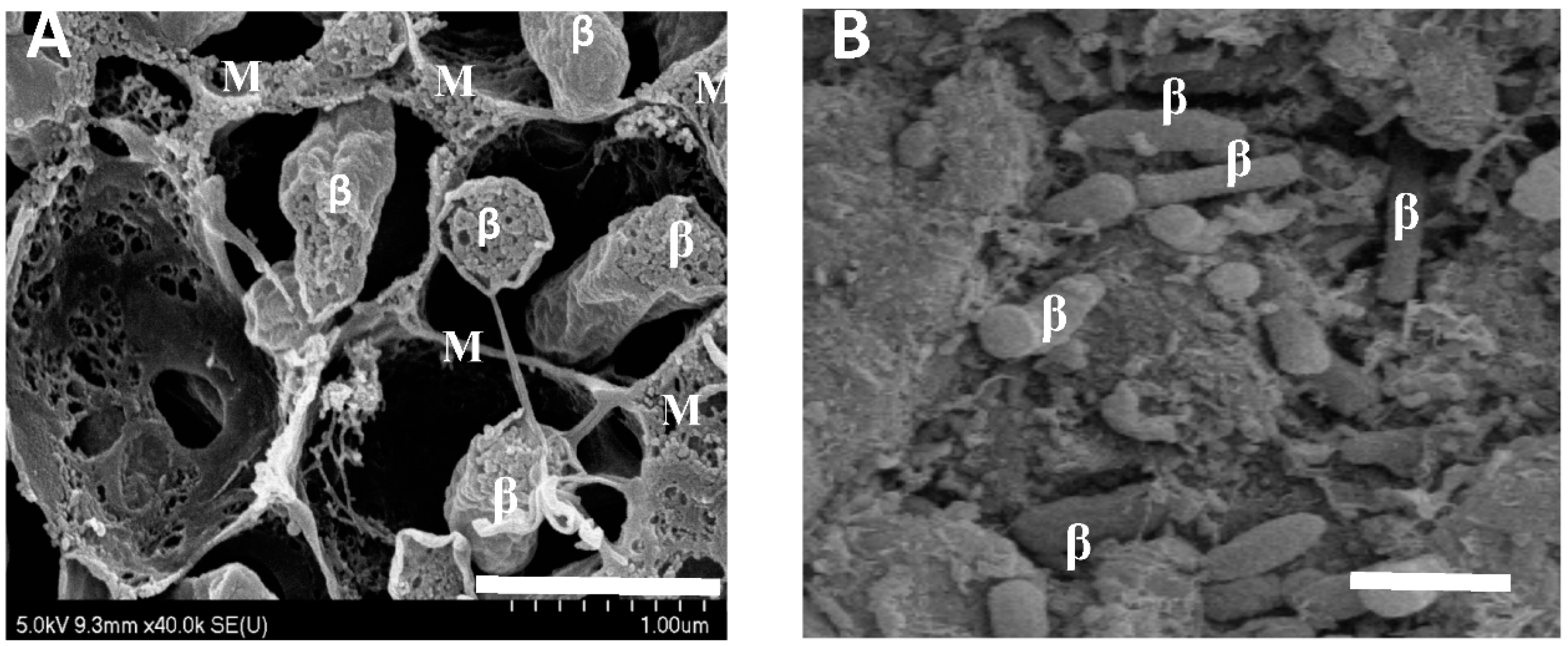
Figure 2.
Scanning electron micrographs of soybean nodules 42 and 95 days after planting. (A) A soybean nodule at 42 days after planting. Bar represents 1 μm. (B) A soybean nodule at 95 days after planting. Bar represents 1 μm. Bacteroids are indicated as “β”; membranes and cellular material are indicated by “M”.
Figure 2.
Scanning electron micrographs of soybean nodules 42 and 95 days after planting. (A) A soybean nodule at 42 days after planting. Bar represents 1 μm. (B) A soybean nodule at 95 days after planting. Bar represents 1 μm. Bacteroids are indicated as “β”; membranes and cellular material are indicated by “M”.
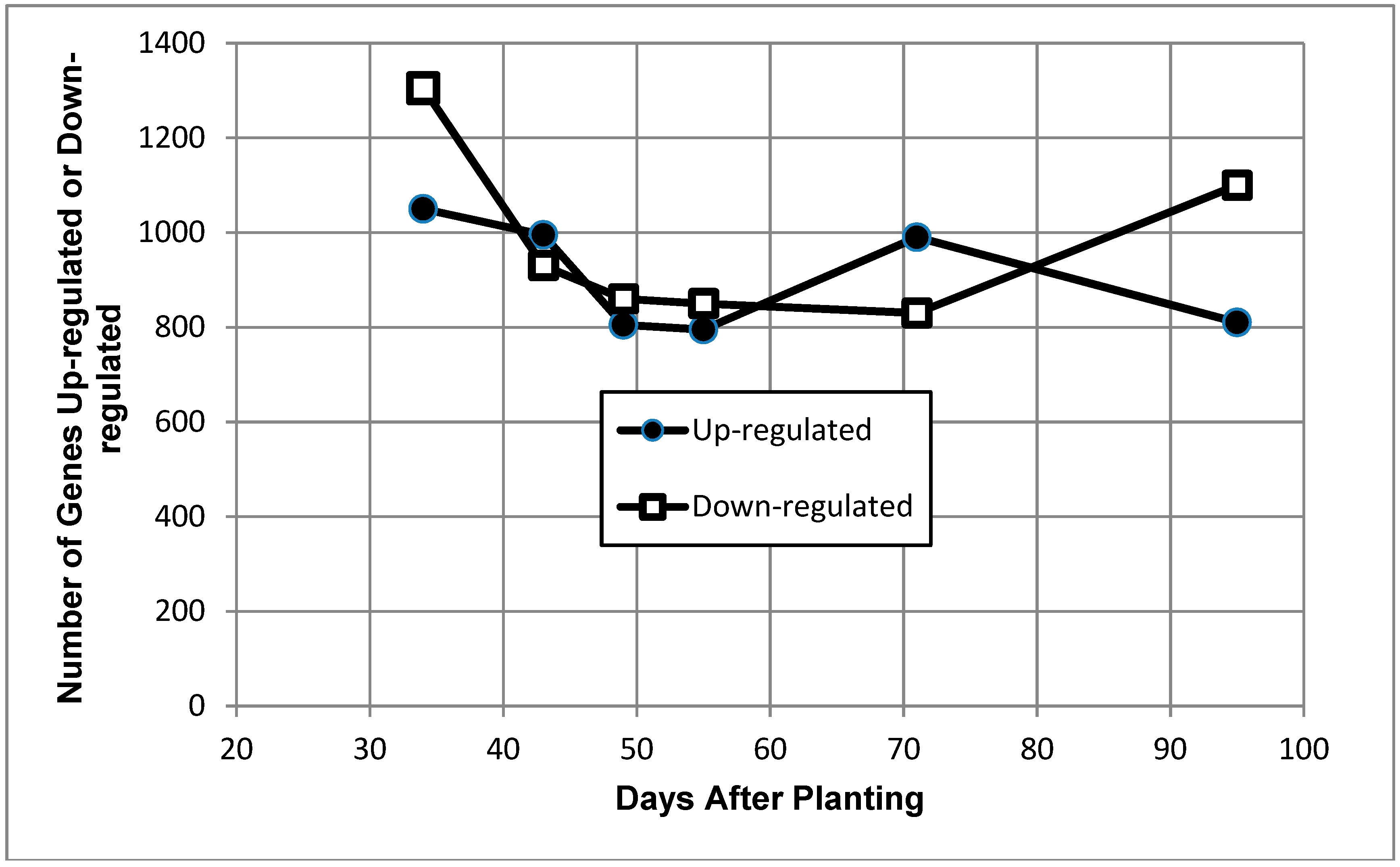
Figure 3.
Number of up- and down-regulated genes of Bradyrhizobium diazoefficiens bacteroids as a function of days after planting. The transcripts included at each time point were determined independently of their values at other time points; thus, transcripts are typically included in multiple time points and may occur in either category. Solid symbols are up-regulated genes, and open symbols are down-regulated genes.
Figure 3.
Number of up- and down-regulated genes of Bradyrhizobium diazoefficiens bacteroids as a function of days after planting. The transcripts included at each time point were determined independently of their values at other time points; thus, transcripts are typically included in multiple time points and may occur in either category. Solid symbols are up-regulated genes, and open symbols are down-regulated genes.
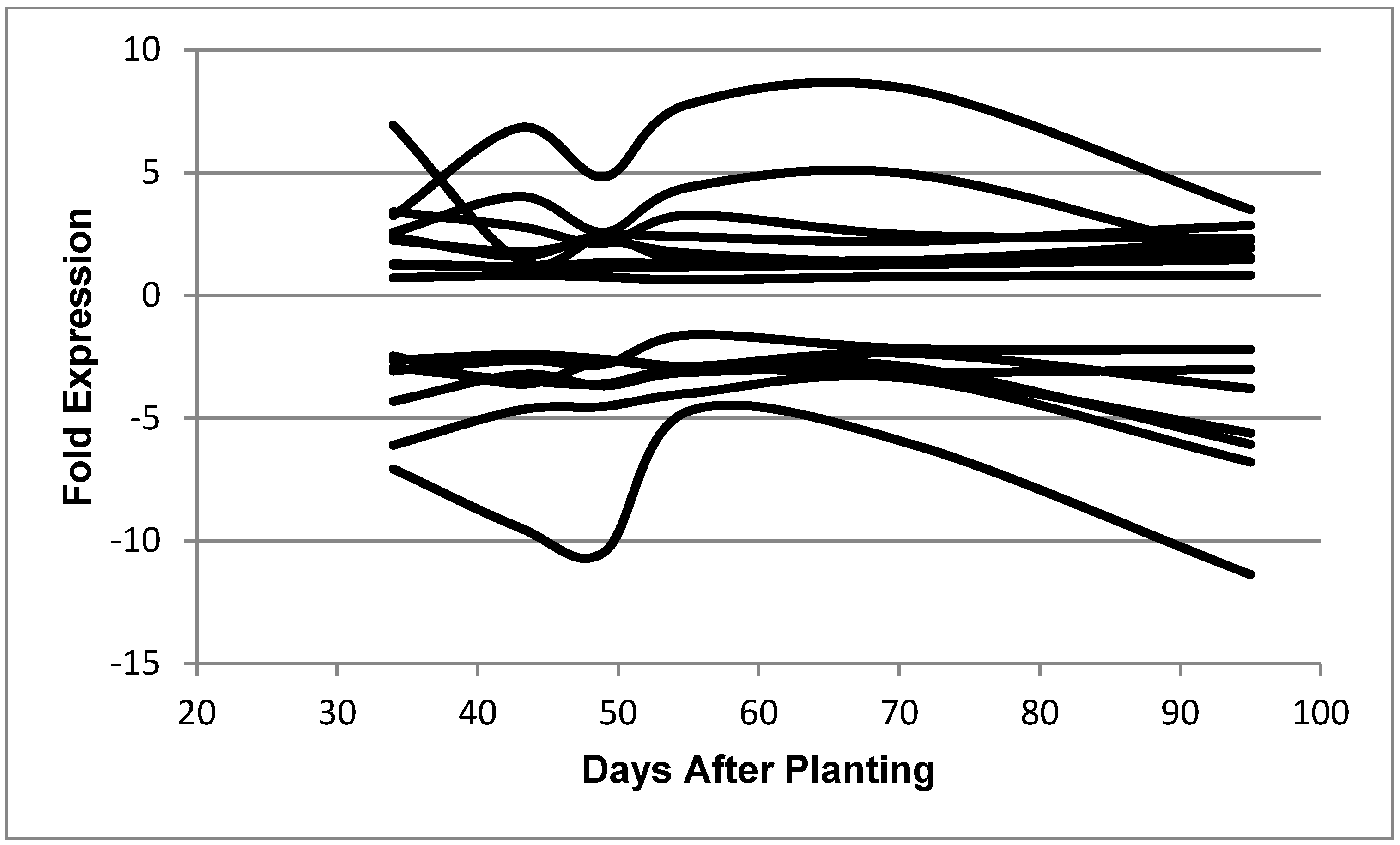
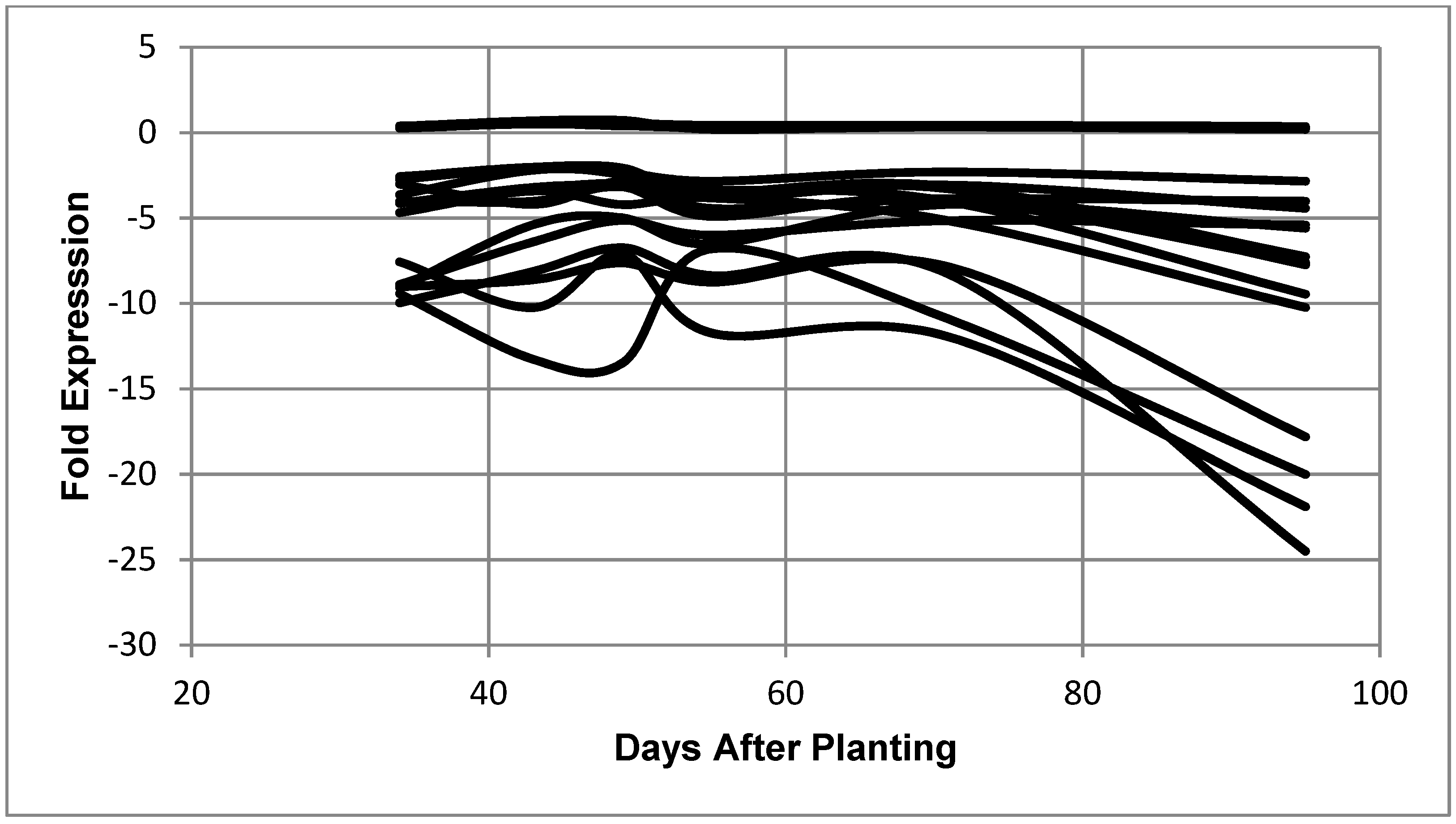
Figure 4.
Expression of B. diazoefficiens bacteroid genes related to transcription as a function of days after planting. Fold expression of the transcription-related genes at 95 days after planting: RNA polymerase sigma-54 subunit (3.50), putative RNA polymerase sigma factor protein (2.86), RNA polymerase sigma-54 subunit (2.33), sigma32-like factor (2.25), RNA polymerase sigma-E factor (1.93), sigma-54 modulation protein (1.55), ECF sigma factor (1.49), ECF family sigma factor (1.47), RNA polymerase (0.82), cold-shock dead-box protein A (−2.20), DNA-directed RNA polymerase beta chain (−3.01), DNA-directed RNA polymerase beta chain (−3.79), RNA polymerase omega subunit (−5.60), transcription anti-termination protein (−6.06), DNA-directed RNA polymerase alpha subunit (−6.78), and ATP-dependent RNA helicase (−11.38).
Figure 4.
Expression of B. diazoefficiens bacteroid genes related to transcription as a function of days after planting. Fold expression of the transcription-related genes at 95 days after planting: RNA polymerase sigma-54 subunit (3.50), putative RNA polymerase sigma factor protein (2.86), RNA polymerase sigma-54 subunit (2.33), sigma32-like factor (2.25), RNA polymerase sigma-E factor (1.93), sigma-54 modulation protein (1.55), ECF sigma factor (1.49), ECF family sigma factor (1.47), RNA polymerase (0.82), cold-shock dead-box protein A (−2.20), DNA-directed RNA polymerase beta chain (−3.01), DNA-directed RNA polymerase beta chain (−3.79), RNA polymerase omega subunit (−5.60), transcription anti-termination protein (−6.06), DNA-directed RNA polymerase alpha subunit (−6.78), and ATP-dependent RNA helicase (−11.38).
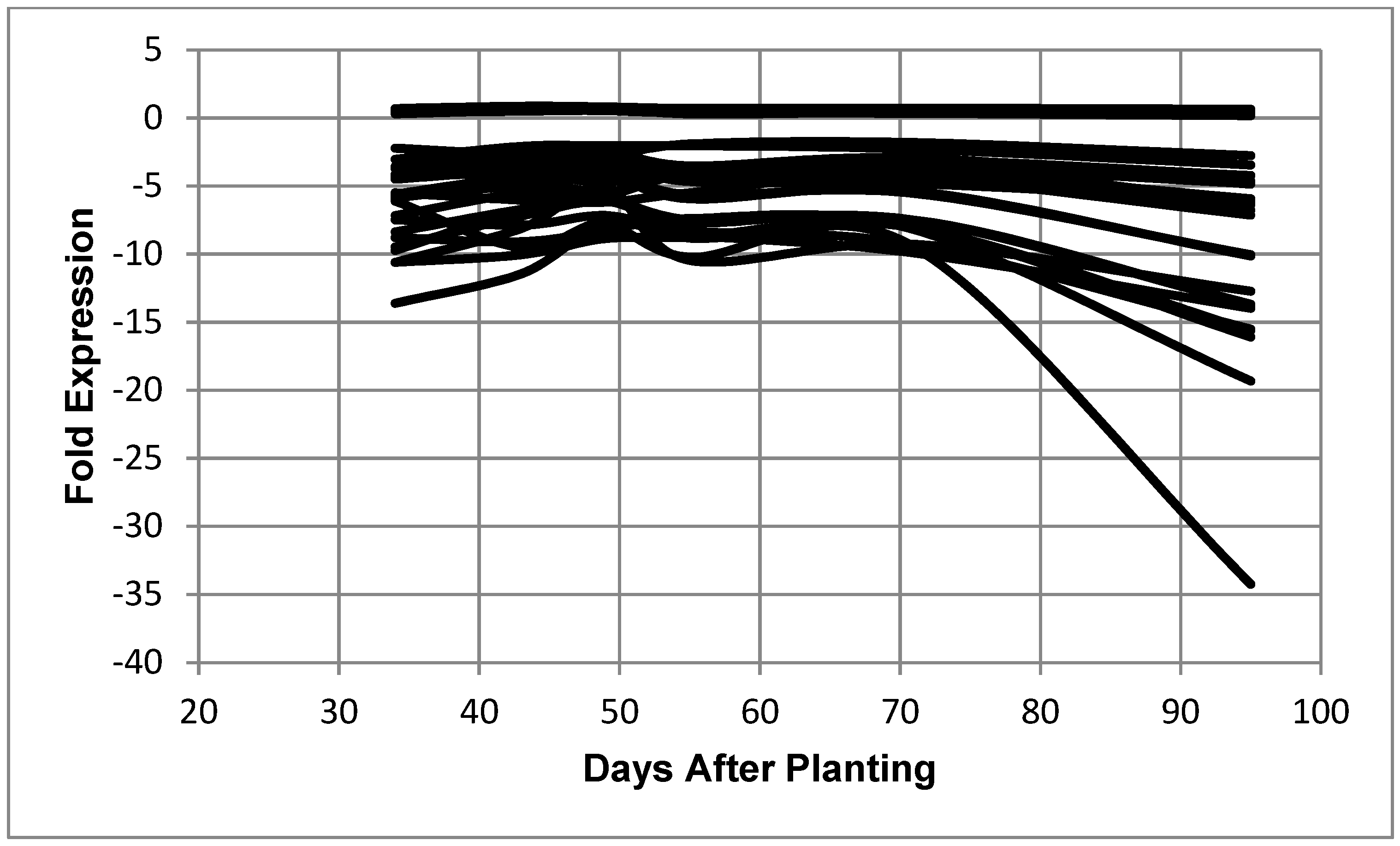
Figure 5.
Expression of B. diazoefficiens bacteroid 30S ribosomal proteins as a function of days after planting. Fold expression value of 30S ribosomal proteins at 95 days after planting: S16 (0.36), S2 (0.29), S20 (0.27), S4 (0.20), S13 (0.19), S12 (−2.85), S19 (4.02), S6 (−4.43), S8 (−5.41), S14 (−5.60), S18 (−7.27), S7 (−7.61), S11 (−7.74), S1 (−9.46), S15 (−10.24), S3 (−17.81), S21 (−20.02), S10 (−21.90), and S17 (−24.51).
Figure 5.
Expression of B. diazoefficiens bacteroid 30S ribosomal proteins as a function of days after planting. Fold expression value of 30S ribosomal proteins at 95 days after planting: S16 (0.36), S2 (0.29), S20 (0.27), S4 (0.20), S13 (0.19), S12 (−2.85), S19 (4.02), S6 (−4.43), S8 (−5.41), S14 (−5.60), S18 (−7.27), S7 (−7.61), S11 (−7.74), S1 (−9.46), S15 (−10.24), S3 (−17.81), S21 (−20.02), S10 (−21.90), and S17 (−24.51).
Figure 6.
Expression of B. diazoefficiens bacteroid 50S ribosomal proteins as a function of days after planting. Fold expression value of 50S ribosomal proteins at 95 days after planting: L18 (0.65), L27 (0.40), L7/12 (0.38), L31 (0.33), L19 (0.20), L28 (0.17), L10 (−2.76), L21 (−3.44), L25 (−4.22), L13 (−4.65), L11 (−4.82), L2 (−4.86), L23 (−5.93), L9 (−6.07), L17 (−6.21), L15 (−6.38), L22 (−6.76), L20 (−7.13), L1 (−10.04), L35 (−10.13), L6 (−12.72), L24 (−13.68), L14 (−13.71), L3 (−13.98), L29 (−15.50), L5 (−15.63), L4 (−16.09), L16 (−19.32), and L30 (−34.25).
Figure 6.
Expression of B. diazoefficiens bacteroid 50S ribosomal proteins as a function of days after planting. Fold expression value of 50S ribosomal proteins at 95 days after planting: L18 (0.65), L27 (0.40), L7/12 (0.38), L31 (0.33), L19 (0.20), L28 (0.17), L10 (−2.76), L21 (−3.44), L25 (−4.22), L13 (−4.65), L11 (−4.82), L2 (−4.86), L23 (−5.93), L9 (−6.07), L17 (−6.21), L15 (−6.38), L22 (−6.76), L20 (−7.13), L1 (−10.04), L35 (−10.13), L6 (−12.72), L24 (−13.68), L14 (−13.71), L3 (−13.98), L29 (−15.50), L5 (−15.63), L4 (−16.09), L16 (−19.32), and L30 (−34.25).
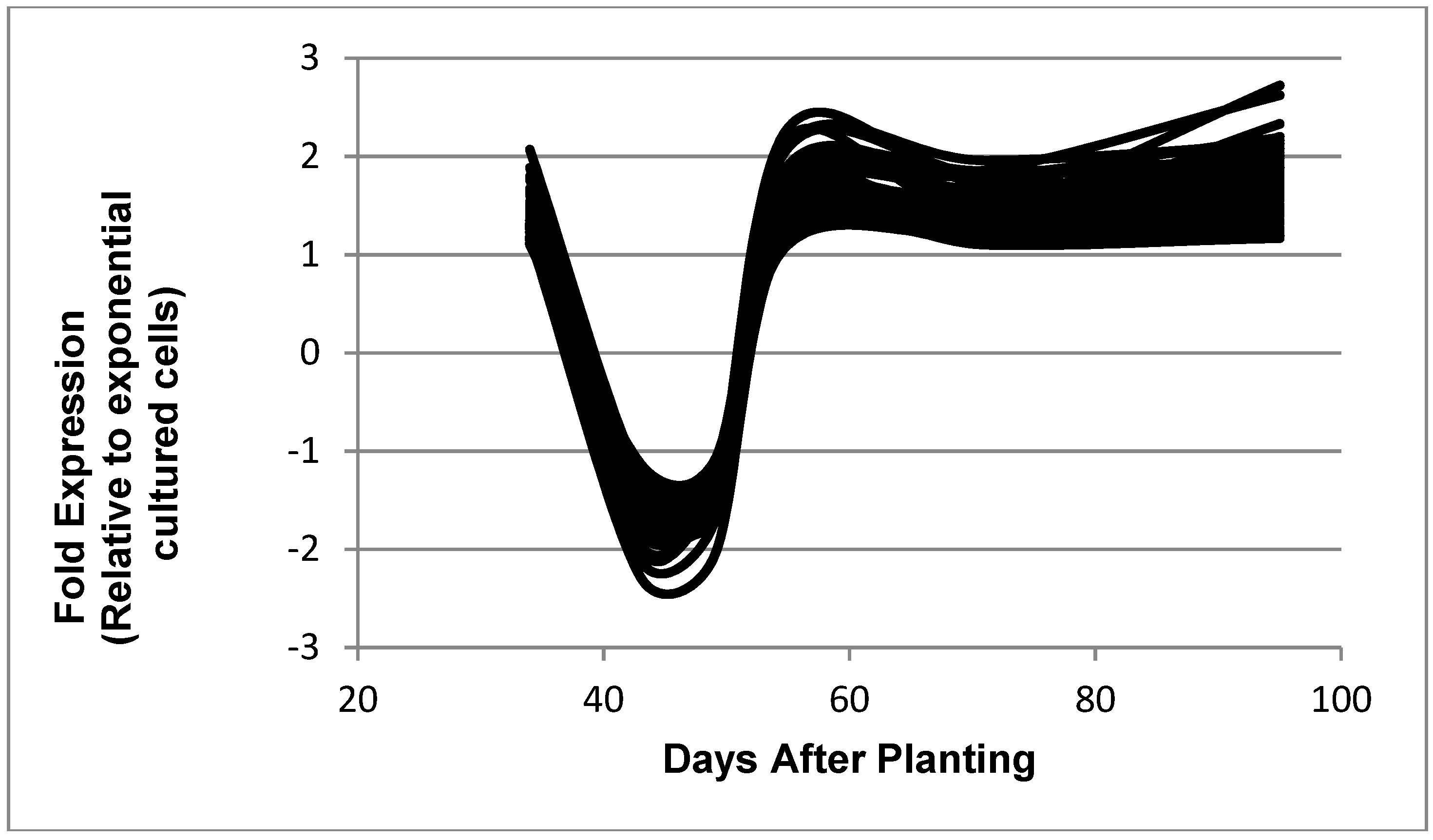
Figure 7.
Expressed genes representing the reciprocal symbiosis pattern. The 68 genes shown in the figure are: bll0116, bll0195, bll0776, bll0979, bll1109, bll1422, bll2116, bll2410, bll2494, bll2675, bll3374, bll3444, bll3470, bll3505, bll3783, bll4141, bll4201, bll4403, bll4596, bll4793, bll5240, bll5296, bll5959, bll6302, bll6471, bll6498, bll6891, bll7298, bll7311, bll7402, bll7610, bll7835, bll7964, blr0310, blr0925, blr1457, blr2533, blr2715, blr2809, blr3324, blr3400, blr3517, blr4155, blr4311, blr4566, blr4772, blr5162, blr5169, blr5229, blr6118, blr6544, blr6569, blr6661, blr6771, blr7060, blr7216, blr7281, blr7496, blr7595, blr7717, blr7815, blr8000, blr8060, blr8194, bsl3895, bsl7758, bsr1887, and bsr3197.
Figure 7.
Expressed genes representing the reciprocal symbiosis pattern. The 68 genes shown in the figure are: bll0116, bll0195, bll0776, bll0979, bll1109, bll1422, bll2116, bll2410, bll2494, bll2675, bll3374, bll3444, bll3470, bll3505, bll3783, bll4141, bll4201, bll4403, bll4596, bll4793, bll5240, bll5296, bll5959, bll6302, bll6471, bll6498, bll6891, bll7298, bll7311, bll7402, bll7610, bll7835, bll7964, blr0310, blr0925, blr1457, blr2533, blr2715, blr2809, blr3324, blr3400, blr3517, blr4155, blr4311, blr4566, blr4772, blr5162, blr5169, blr5229, blr6118, blr6544, blr6569, blr6661, blr6771, blr7060, blr7216, blr7281, blr7496, blr7595, blr7717, blr7815, blr8000, blr8060, blr8194, bsl3895, bsl7758, bsr1887, and bsr3197.
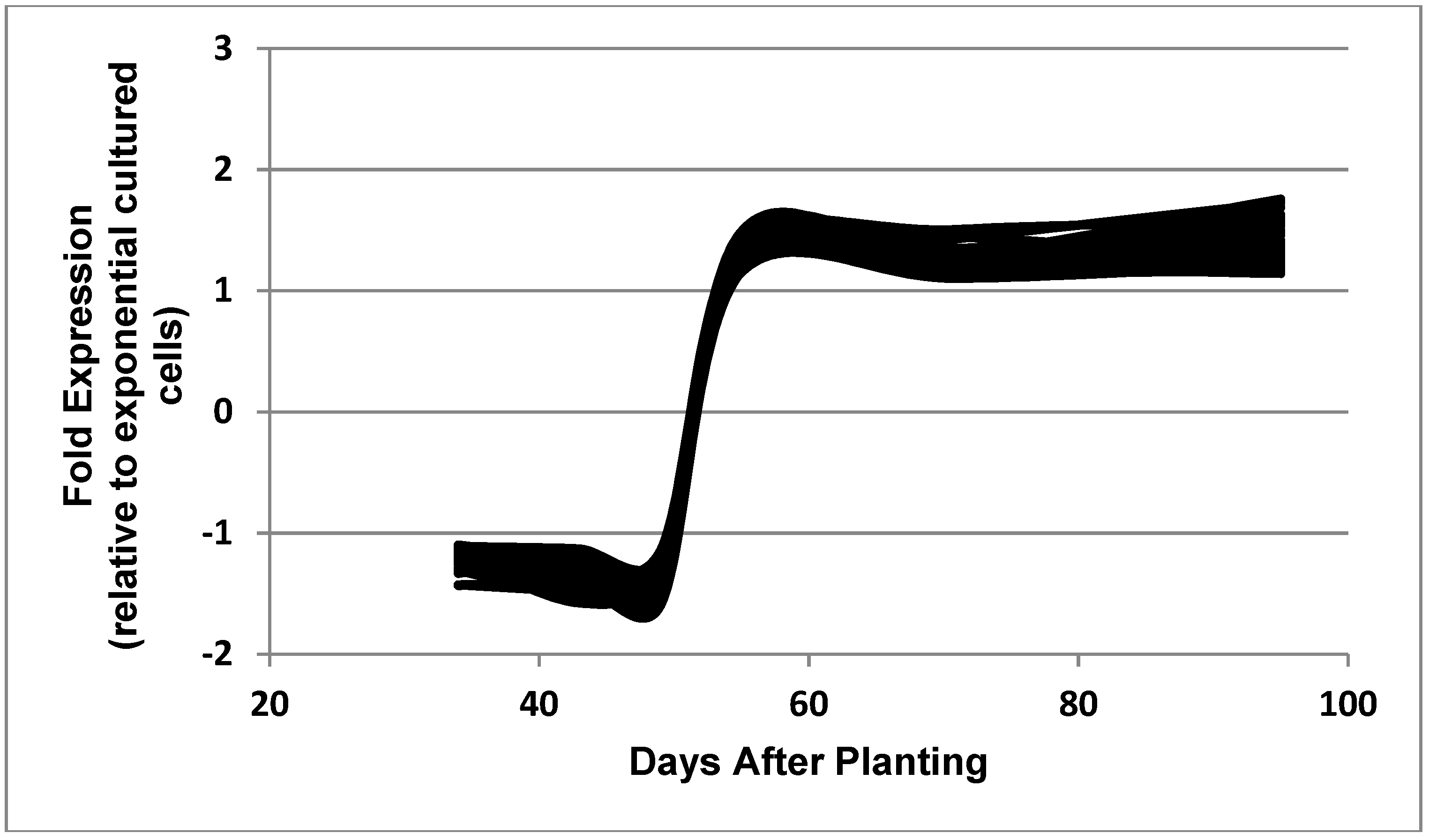
Figure 8.
Expressed genes representing the post-symbiotic category. The 33 genes shown in the figure are: bll3172, bll4771, bll6387, bll6597, bll8078, blr0490, blr0998, blr2666, blr2834, blr2882, blr2923, blr3110, blr3499, blr3603, blr3634, blr3898, blr4361, blr5112, blr7482, blr7579, bsr1885, bll0052, bll0242, bll0244, bll0493, bll1533, bll2075, bll2188, bll4292, bll6051, blr4337, blr7208, and blr7482.
Figure 8.
Expressed genes representing the post-symbiotic category. The 33 genes shown in the figure are: bll3172, bll4771, bll6387, bll6597, bll8078, blr0490, blr0998, blr2666, blr2834, blr2882, blr2923, blr3110, blr3499, blr3603, blr3634, blr3898, blr4361, blr5112, blr7482, blr7579, bsr1885, bll0052, bll0242, bll0244, bll0493, bll1533, bll2075, bll2188, bll4292, bll6051, blr4337, blr7208, and blr7482.
Table 1.
Categories of Expressed and Constitutive Genes. Each identified gene was placed in the corresponding category as defined by Kaneko et al. [
22]. The values in parenthesis are the percentage that each category represents of the total number of transcripts in each group (up-regulated, down-regulated, or constitutive). The transcripts included in either the up- (>2-fold) or down-regulated (<2-fold) columns were genes that display the respective values at all six time points.
Table 1.
Categories of Expressed and Constitutive Genes. Each identified gene was placed in the corresponding category as defined by Kaneko et al. [
22]. The values in parenthesis are the percentage that each category represents of the total number of transcripts in each group (up-regulated, down-regulated, or constitutive). The transcripts included in either the up- (>2-fold) or down-regulated (<2-fold) columns were genes that display the respective values at all six time points.
| Category | Upregulated | Downregulated | Constitutive | Total |
|---|
| Amino acid metabolism | 9 (1.4%) | 32 (3.3%) | 17 (1.8%) | 58 |
| Cofactors, prosthetic groups, and carriers | 13 (2.0%) | 20 (2.0%) | 12 (1.3%) | 45 |
| Cell envelope | 4 (0.6%) | 28 (2.9%) | 14 (1.5%) | 46 |
| Cellular processes | 15 (2.3%) | 66 (6.8%) | 60 (6.5%) | 141 |
| Central intermediary metabolism | 40 (6.1%) | 30 (3.1%) | 46 (5.0%) | 116 |
| DNA replication, recombination, and repair | 5 (0.8%) | 11 (1.1%) | 9 (1.0%) | 25 |
| Energy metabolism | 19 (3%) | 54 (5.5%) | 40 (4.3%) | 113 |
| Fatty acid, phospholipid, and sterol metabolism | 6 (0.9%) | 23 (2.4%) | 17 (1.8%) | 46 |
| Purines, pyrimidines, nucleosides, and nucleotides | 2 (0.3%) | 15 (1.5%) | 11 (1.2%) | 28 |
| Regulatory functions | 39 (6.0%) | 56 (5.7%) | 55 (5.9%) | 150 |
| Transcription | 5 (0.8%) | 10 (1.0%) | 11 (1.2%) | 26 |
| Translation | 9 (1.4%) | 65 (6.7%) | 51 (5.5%) | 125 |
| Transport and binding proteins | 46 (7.0%) | 61 (6.3%) | 63 (6.8%) | 170 |
| Other functions | 72 (11.0%) | 81 (8.3%) | 78 (8.4%) | 231 |
| Hypothetical | 369 (56.5%) | 424 (43.4%) | 443 (47.8%) | 1236 |
| Total | 653 | 976 | 927 | 2556 |
Table 2.
Examples of
B. diazoefficiens genes expressed during the symbiosis of soybean root nodules following the pattern displayed in
Figure 1B.
Table 2.
Examples of
B. diazoefficiens genes expressed during the symbiosis of soybean root nodules following the pattern displayed in
Figure 1B.
| Function | Rhizobase | Accession | Location |
|---|
| ABC transporter substrate-binding protein | Bll4544 | BAC49809 | Periplasm |
| FixK2 protein | Blr2757 | CAA06287 | Cytoplasmic |
| C-4 Dicarboxylate transport protein | Bll1718 | NP_768358 | Cytoplasmic membrane |
| Iron response regulator | Bll0768 | AHY55539 | Cytoplasm |
| Citrate synthase | Blr4839 | NP_771479 | Cytoplasm |
| Aspartate aminotransferase | Blr1686 | NP_768326 | Cytoplasm |
| Divalent cation resistant protein | Blr4935 | NP_771575 | Outer Membrane |
| Alcohol dehydrogenase | Bll4482 | NP_771122 | Unknown |
| Taurine Dioxygenase | Bll2125 | APO50734 | Cytoplasm |
| Β-Lactamase | Bll2252 | AND87779 | Cytoplasm |
| Tripartite tricarboxylate transporter TctB | Bll3050 | AND88496 | Cytoplasmic membrane |
| Tripartite tricarboxylate transporter substrate binding protein | Blr3161 | AWO90155 | Periplasm |
| 3-β hydroxysteroid dehydrogenase | Bll4299 | AND89578 | Cytoplasm |
| C4-Dicarboxylate ABC transporter | Blr5025 | AND90243 | Periplasm |
| (2Fe-2S)-binding protein | Bll6238 | AND91348 | Cytoplasm |
| Hemolysin D | Bll6258 | AND91366 | Cytoplasmic membrane |
| Hemolysin secretion protein D | Blr3031 | AND88479 | Unknown |
| Dienelactone hydrolase | Bll7509 | AWO94311 | Unknown |
| Serine hydrolase | Bll8153 | AWO94832 | Cytoplasmic membrane |
| Asp/Glu/hydantoin racemase | Blr3294 | AND88720 | Unknown |
| Phospholipase | Blr5550 | AWO92367 | Unknown |
| Allophanate hydrolase | Blr3633 | AND88989 | Cytoplasm |
| Anhydro-N-acetylmuramic acid kinase | Blr4331 | AND89607 | Unknown |
| Dolichol-phosphate mannosyltransferase | Blr4442 | NP_771082 | Cytoplasmic membrane |
| ATPase | Blr7361 | AND92326 | Unknown |
| 2-Hydroxyhepta-2,4-diene-1,7-dioate isomerase | Blr7891 | AND92773 | Cytoplasm |
Table 3.
Examples of
B. diazoefficiens genes expressed during the senescence of soybean root nodules following the pattern displayed in
Figure 7.
Table 3.
Examples of
B. diazoefficiens genes expressed during the senescence of soybean root nodules following the pattern displayed in
Figure 7.
| Function | Rhizobase | Accession | Location |
|---|
| Glutamate synthetase I | Blr4835 | BAC50100 | Cytoplasm |
| Proline iminopeptidase | Bll4403 | BAC49668 | Cytoplasm |
| Pantoate-beta-alanine ligase | Blr6152 | BAC50427 | Cytoplasm |
| β-ketoadipyl CoA thiolase | Blr0925 | BAC46190 | Unknown |
| Glycerate dehydrogenase | Bll2918 | BAC48183 | Cytoplasm |
| Cytochrome C oxidase | Bll3783 | BAC49048 | Cytoplasmic membrane |
| Murein endopeptidase | Blr8158 | BAC53423 | Periplasm |
| Ribose 5-phosphate isomerase | Blr3755 | BAC49020 | Cytoplasm |
| 3-Deoxy-manno-octulosonate cytidylyltransferase | Bll1422 | BAC46687 | Cytoplasm |
| Sugar ABC transporter permease | Bll2675 | BAC47940 | Cytoplasmic membrane |
| Flagellar protein | Bll6868 | BAC52133 | Extracellular |
| 3-Oxoacyl-(acyl-carrier-protein) synthase II | Bll3809 | BAC49074 | Unknown |
| Branched-chain amino acid ABC transporter substrate-binding protein | Bll0979 | BAC46244 | Periplasm |
| Cytochrome O ubiquinol oxidase | Blr2715 | AND93893 | Cytoplasmic membrane |
| Glycosyl hydrolase | Blr6771 | NP_773411 | Cytoplasm |
| Alkylhydroperoxidase | Bl57595 | AND92531 | Unknown |
| 3-Oxoacyl-ACP reductase | Bll4596 | AWO91529 | Periplasm |
| Methyltransferase | Blr2533 | AWO89558 | Cytoplasm |
| Patatin-like phospholipase | Bll7964 | AWO94740 | Extracellular |
| LysE family translocator | Bll6498 | PDT61394 | Cytoplasmic membrane |
| Amidohydrolase | Bll7610 | AWO94407 | Cytoplasm |
| Phosphodiesterase | Bll4141 | AWO91078 | Cytoplasm |
| Pyridoxamine 5′-phosphate oxidase | Blr4155 | AWO91090 | Cytoplasm |
| Peptide ABC transporter permease | Blr0310 | BAC45575 | Cytoplasmic membrane |
| TetR-like transcriptional regulator | Bll7298 | BAC52563 | Unknown |
| enoyl-CoA hydratase | Bll0116 | AWO87220 | Cytoplasm |
Table 4.
Examples of
B. diazoefficiens genes expressed during the senescence of soybean root nodules following the pattern displayed in
Figure 8.
Table 4.
Examples of
B. diazoefficiens genes expressed during the senescence of soybean root nodules following the pattern displayed in
Figure 8.
| Function | Rhizobase | Accession | Location |
|---|
| Citrate-proton symporter | Bll1864 | WP_06090909 | Cytoplasmic Membrane |
| Ribose 5-phosphate isomerase | Blr3755 | NP_770395 | Cytoplasm |
| Acyltransferase | Blr4337 | AND94080 | Cytoplasmic Membrane |
| Thymydilate kinase | Bll4518 | NP_771158 | Cytoplasm |
| Homoserine O-succinyltransferase | Bll0244 | BAC45509 | Cytoplasm |
| Proline iminopeptidase | Bll4403 | NP_771043 | Cytoplasm |
| 3-Dehydroquinate dehydratase | Bll4292 | NP_770932 | Cytoplasm |
| Epoxide hydrolase | Blr2881 | NP_769521 | Unknown |
| ATP-dependent DNA helicase | Bll0242 | NP_766882 | Cytoplasm |
| Pyridoxamine 5′-phosphate oxidase | Bll7835 | AND92725 | Unknown |
| Beta-ketoadipyl CoA thiolase | Blr0925 | NP_767565 | Cytoplasm |
| Branched chain amino acid ABC transporter permease protein | Blr2923 | NP_769563 | Cytoplasmic Membrane |
| Sugar ABC transporter permease protein | Bll2675 | NP_769315 | Cytoplasmic Membrane |
| 3-Deoxy-manno-octulosonate cytidylyltransferase | Bll1422 | WP_011084239 | Cytoplasm |
| Exoribonuclease | Blr5112 | NP_771752 | Cytoplasm |
| Δ9 acyl-lipid fatty acid desaturase | Bll4594 | WP_011087365 | Cytoplasmic Membrane |
| DNA glycosylase | Blr6661 | NP_773301 | Unknown |
| Outer membrane channel lipoprotein | Bll4321 | NP_770961 | Outer Membrane |
| Cytochrome P450 | Bll0557 | NP_767197 | Unknown |
| Transcriptional regulatory protein IclR family | Blr3939 | NP_770579 | Cytoplasm |
| Purine-binding chemotaxis protein | Blr2193 | NP_768833 | Cytoplasm |
| Patatin-like phospholipases | Bll7964 | AWO94740 | Cytoplasm |
| Branched-chain amino acid ABC transporter substrate-binding protein | Bll0979 | WP_011083799 | Periplasm |
| Phosphoserine aminotransferase | Bll7402 | NP_774042 | Cytoplasm |
| Rubrerythrin | Blr7895 | KOY06556 | Cytoplasmic Membrane |
Table 5.
The frequency of rare codons in each expression pattern and in the transcription and translation functions. Values are frequency per 1000 codons. The lettered superscripts indicate a statistical difference (p < 0.5) between similar indicated values. The asterisks (*) indicate that the value of 0 is statistically different from each of the other categories.
Table 5.
The frequency of rare codons in each expression pattern and in the transcription and translation functions. Values are frequency per 1000 codons. The lettered superscripts indicate a statistical difference (p < 0.5) between similar indicated values. The asterisks (*) indicate that the value of 0 is statistically different from each of the other categories.
| Expression Pattern/Function | Rare Codon |
|---|
| TTA | TCT | TGT | CTA | ATA | ACT | AGT | AGA | GTA | All |
|---|
| Symbiotic | 1.30ABC | 3.01ABCD | 2.20AB | 1.91AB | 2.66A | 4.11ABCD | 3.41ABC | 3.09ABC | 2.80AB | 2.72ABCDEF |
| Post-Symbiotic | 0.43A | 2.31 | 1.33 | 2.23CD | 2.01 | 3.30E | 2.45D | 2.84DEF | 2.91 | 2.20AGHIJ |
| Reciprocal Symbiotic | 1.07 | 2.76 | 1.46 | 2.18 | 1.34 | 3.00F | 2.30E | 2.16G | 2.28 | 2.06BK |
| Constitutive | 0.35B | 1.06A | 0.97 | 0.80AC | 1.05 | 1.65AEF | 1.62AF | 1.48AH | 1.89 | 1.20CGKLMN |
| Transcription | 0.49C | 1.43B | 0.53A | 1.20 | 0.43A | 2.18B | 2.49G | 1.29BDEI | 1.30A | 1.26DHLO |
| 30S Ribosome Genes | 0 * | 1.72C | 0 * | 0 * | 0 * | 2.36C | 1.07B | 0.78 | 0.57B | 0.72EIM |
| 50S Ribosome Genes | 0 * | 1.82D | 0.43B | 0.12BD | 0 * | 2.69D | 0.29CDEFG | 0.14CFGHI | 1.87 | 0.82FJNO |
Table 6.
Frequency of GC, AG, and nucleotide contents within each expression pattern and the transcription and translation functional groups. Values are frequency per 1000 codons. Superscripts indicate a statistical difference (p < 0.5) between similar indicated values.
Table 6.
Frequency of GC, AG, and nucleotide contents within each expression pattern and the transcription and translation functional groups. Values are frequency per 1000 codons. Superscripts indicate a statistical difference (p < 0.5) between similar indicated values.
| Expression Pattern/Function | %GC | %AG | %A | %T | %G | %C |
|---|
| Symbiotic | 63.4AB | 57.92ABCDEF | 18.59AB | 18.19ABCD | 31.40AB | 32.02A |
| Post-Symbiotic | 63.88C | 49.64AG | 18.00ACD | 18.11EFGH | 31.64A | 32.23 |
| Reciprocal Symbiotic | 64.13 | 49.19BHKL | 17.72EFG | 18.14IJK | 31.46 | 32.66 |
| Constitutive | 65.18ACD | 50.39CIK | 18.09BHIJ | 16.77AEILM | 32.31BC | 32.87A |
| Transcription | 64.94BE | 51.12DL | 18.92EHKL | 16.15BFJN | 32.16 | 32.77 |
| 30S Ribosome Proteins | 63.23E | 51.93EJ | 20.97CDFIK | 15.81CGL | 30.96CD | 32.26 |
| 50S Ribosome Proteins | 63.62D | 53.08FGHIJ | 21.30GJL | 15.00DHKMN | 31.77D | 31.92 |











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}