Rice Overexpressing OsNUC1-S Reveals Differential Gene Expression Leading to Yield Loss Reduction after Salt Stress at the Booting Stage

 ,
,
Abstract
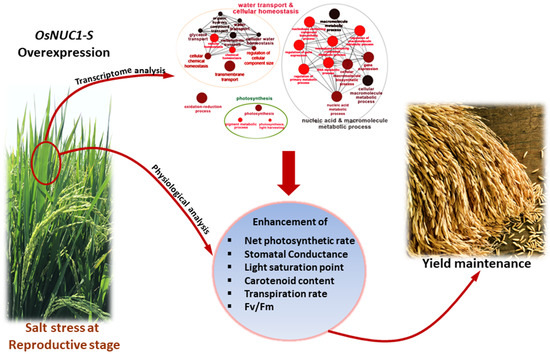
:
1. Introduction
2. Results
2.1. Overexpression of OSNUC1-S Affects Rice Yield
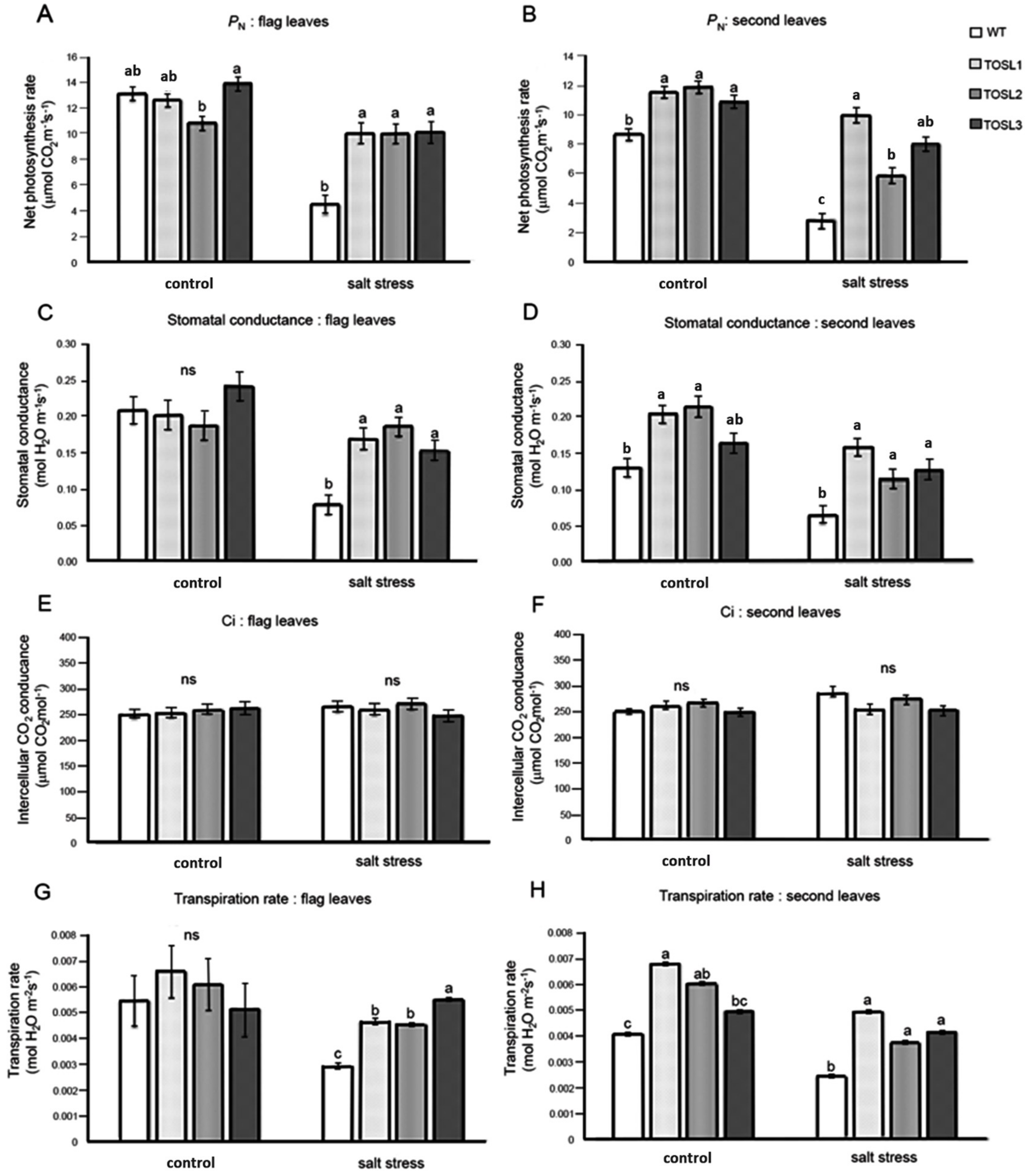
2.2. Overexpression of OSNUC1-S Increased the Photosynthetic Rate, Stomatal Conductance, and Transpiration Rate under Salt-Stress Conditions
2.3. Overexpression of OSNUC1-S Affected Both Light-Response Curves and CO2-Response Curves of Flag and Second Leaves under Salt-Stress Conditions
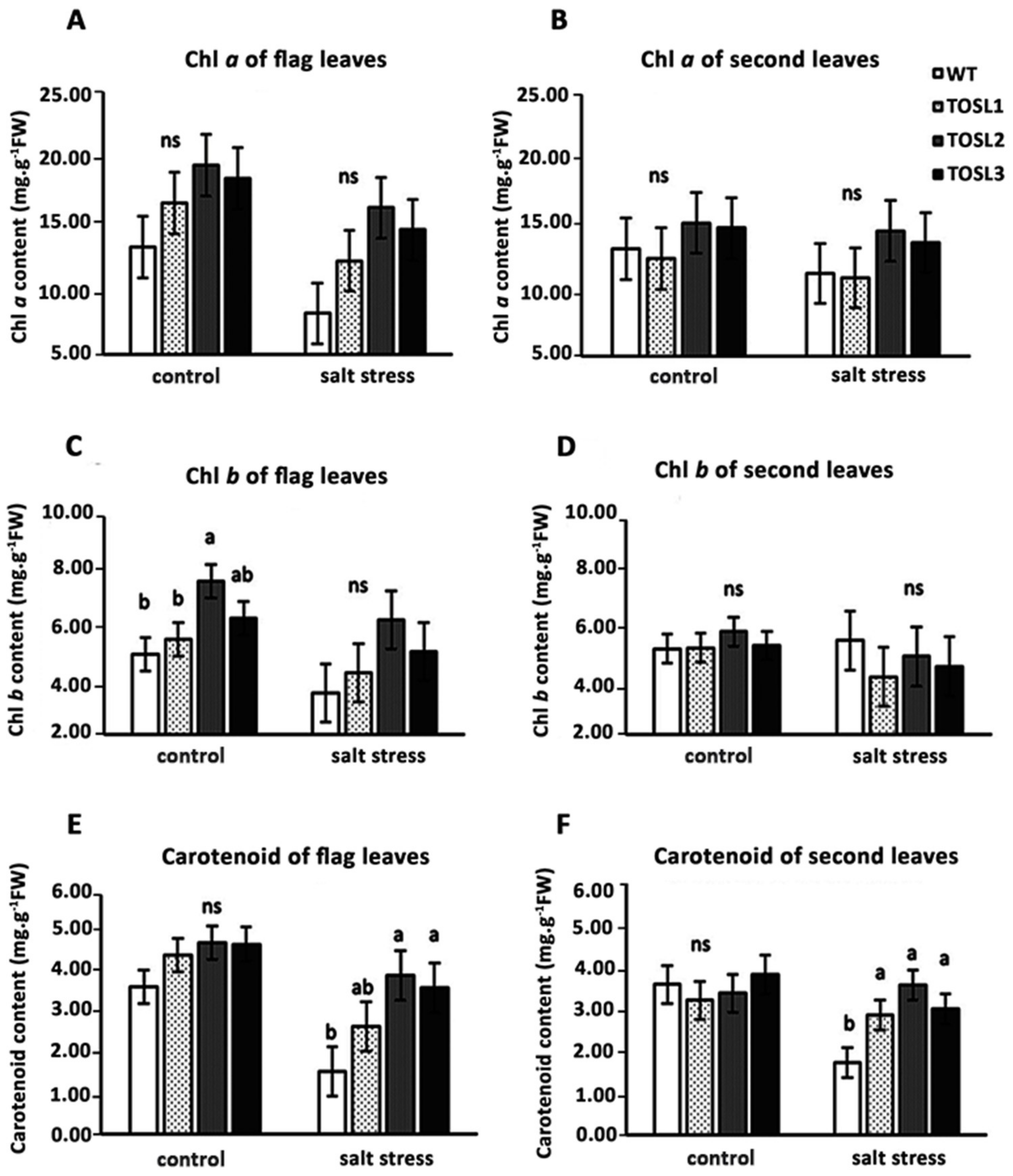
2.4. Salt Stress Affects Photosystem II (PSII) Photochemistry Efficiency and Photosynthetic Pigment Contents in Flag and Second Leaves
2.5. OsNUC1-S’s Overexpression Increased Carbohydrate Metabolism and Sugar Transport in Flag Leaves of Rice Grown Under Control Conditions
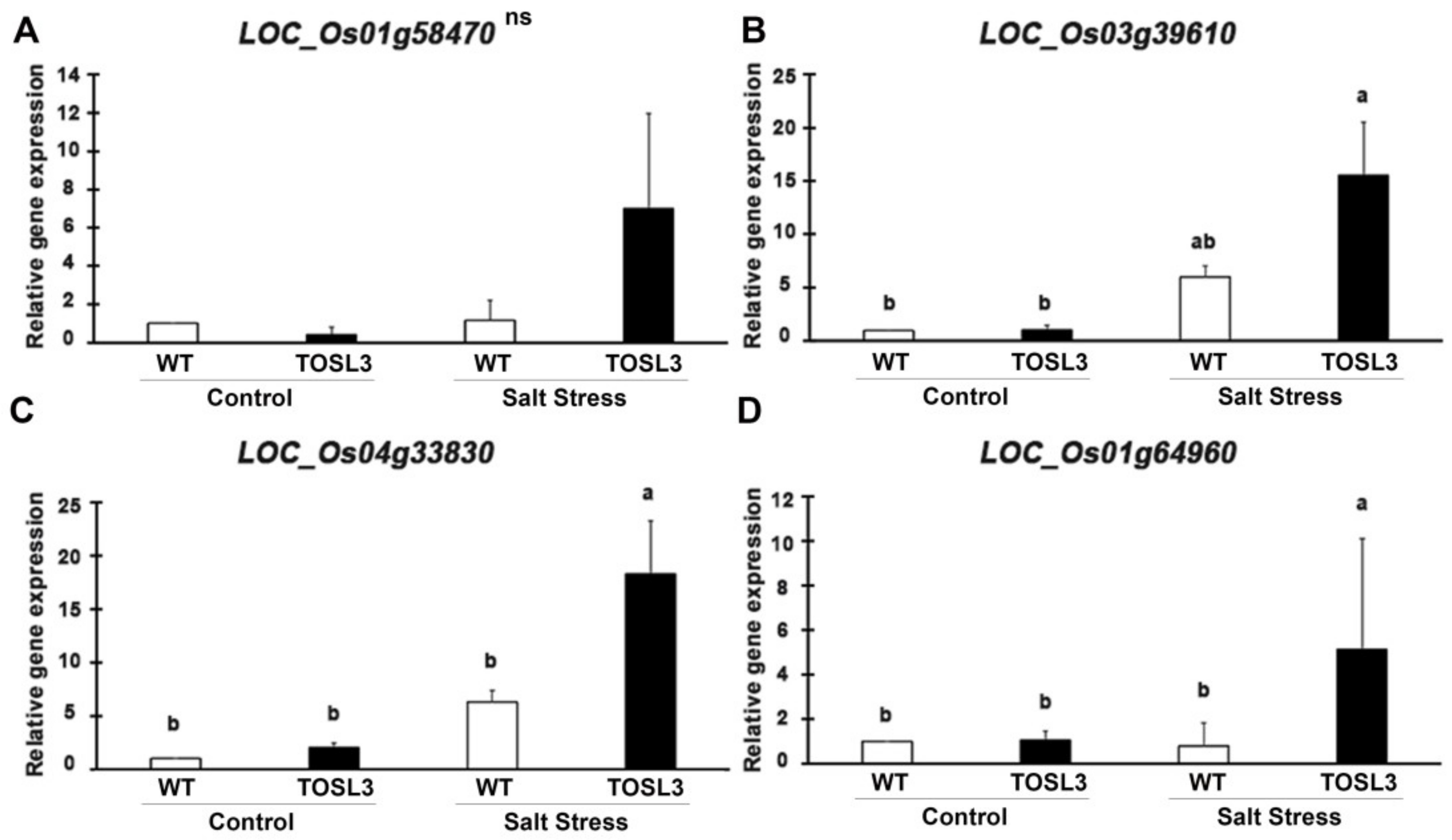
2.6. The Overexpression of OsNUC1-S Increased the Expression of Genes Involved in Water Transport and Cellular Homeostasis, Nucleic Acid and Macromolecule Metabolic Processes, and Photosynthetic Processes
3. Discussion
4. Materials and Methods
4.1. Plant Material and Salt Treatment
4.2. RNA-seq and Data Analysis
4.3. Gene Ontology (GO) Term Analysis
4.4. Pigment Extraction and Quantification
4.5. Yield Collection
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Ci | intercellular CO2 concentration |
| Fv/Fm | intercellular CO2 concentration |
| gs | stomatal conductance |
| OsNUC1 | rice NUCLEOLIN1 |
| PN | net photosynthesis rate |
| WT | wild type |
References
- Fageria, N. Yield physiology of rice. J. Plant Nutr. 2007, 30, 843–879. [Google Scholar] [CrossRef]
- Hakim, M.; Juraimi, A.; Begum, M.; Hanafi, M.; Ismail, M.R.; Selamat, A. Effect of salt stress on germination and early seedling growth of rice (Oryza sativa L.). Afr. J. Biotechnol. 2010, 9, 1911–1918. [Google Scholar]
- Bethke, P.C.; Drew, M.C. Stomatal and nonstomatal components to inhibition of photosynthesis in leaves of Capsicum annuum during progressive exposure to NaCl salinity. Plant Physiol. 1992, 99, 219–226. [Google Scholar] [CrossRef] [PubMed]
- Reddy, M.; Vora, A. Changes in pigment composition, Hill reaction activity and saccharides metabolism in Bajra (Pennisetum typhoides S & H) leaves under NaCl salinity. Photosynthetica 1986, 20, 50–55. [Google Scholar]
- Petrov, V.; Hille, J.; Mueller-Roeber, B.; Gechev, T.S. ROS-mediated abiotic stress-induced programmed cell death in plants. Front. Plant Sci. 2015, 66, 69. [Google Scholar] [CrossRef] [PubMed]
- Lutts, S.; Kinet, J.; Bouharmont, J. Changes in plant response to NaCl during development of rice (Oryza sativa L.) varieties differing in salinity resistance. J. Exp. Bot. 1995, 46, 1843–1852. [Google Scholar] [CrossRef]
- Touch, S.; Pipatpongsa, T.; Takeda, T.; Takemura, J. The relationships between electrical conductivity of soil and reflectance of canopy, grain, and leaf of rice in northeastern Thailand. Int. J. Remote Sens. 2015, 36, 1136–1166. [Google Scholar]
- Zhang, J.; Lin, Y.J.; Zhu, L.F.; Yu, S.M.; Sanjoy, K.K.; Jin, Q.Y. Effects of 1-methylcyclopropene on function of flag leaf and development of superior and inferior spikelets in rice cultivars differing in panicle types. Field Crops Res. 2015, 177, 64–74. [Google Scholar] [CrossRef]
- Tajrishi, M.M.; Tuteja, R.; Tuteja, N. Nucleolin: The most abundant multifunctional phosphoprotein of nucleolus. Commun. Integr. Biol. 2011, 4, 267–275. [Google Scholar] [CrossRef]
- Petricka, J.J.; Nelson, T.M. Arabidopsis nucleolin affects plant development and patterning. Plant Physiol. 2007, 144, 173–186. [Google Scholar] [CrossRef]
- Reichler, S.A.; Balk, J.; Brown, M.E.; Woodruff, K.; Clark, G.B.; Roux, S.J. Light differentially regulates cell division and the mRNA abundance of pea nucleolin during de-etiolation. Plant Physiol. 2001, 125, 339–350. [Google Scholar] [CrossRef] [PubMed]
- Sripinyowanich, S.; Chamnanmanoontham, N.; Udomchalothorn, T.; Maneeprasopsuk, S.; Santawee, P.; Buaboocha, T.; Qu, L.-J.; Gu, H.; Chadchawan, S. Overexpression of a partial fragment of the salt-responsive gene OsNUC1 enhances salt adaptation in transgenic Arabidopsis thaliana and rice (Oryza sativa L.) during salt stress. Plant Sci. 2013, 213, 67–78. [Google Scholar] [CrossRef] [PubMed]
- Udomchalothorn, T.; Plaimas, K.; Sripinyowanich, S.; Boonchai, C.; Kojonna, T.; Chutimanukul, P.; Comai, L.; Buaboocha, T.; Chadchawan, S. OsNucleolin1-L expression in Arabidopsis enhances photosynthesis via transcriptome modification under salt stress conditions. Plant Cell Physiol. 2017, 58, 717–734. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.; Jeong, H.; Lee, S.; Lee, J.; Kim, S.-J.; Park, J.-W.; Woo, H.R.; Lim, P.O.; An, G.; Nam, H.G. Molecular bases for differential aging programs between flag and second leaves during grain-filling in rice. Sci. Rep. 2017, 7, 8792. [Google Scholar] [CrossRef] [Green Version]
- Junghans, T.G.; De Almeida-Oliveira, M.G.; Moreira, M.A. Lipoxygenase activities during development of root and nodule of soybean. Pesqui. Agropecu. Bras. 2004, 39, 625–630. [Google Scholar] [CrossRef]
- Lenglet, A.; Jaślan, D.; Toyota, M.; Mueller, M.; Müller, T.; Schönknecht, G.; Marten, I.; Gilroy, S.; Hedrich, R.; Farmer, E.E. Control of basal jasmonate signalling and defence through modulation of intracellular cation flux capacity. New Phytol. 2017, 216, 1161–1169. [Google Scholar] [CrossRef]
- Brodhun, F.; Cristobal-Sarramian, A.; Zabel, S.; Newie, J.; Hamberg, M.; Feussner, I. An iron 13S-lipoxygenase with an α-linolenic acid specific hydroperoxidase activity from Fusarium oxysporum. PLoS ONE 2013, 8, e64919. [Google Scholar] [CrossRef]
- Kholupenko, I.; Voronkova, N.; Burundukova, O.; Zhemchugova, V. Demand for assimilates determines the productivity of intensive and extensive rice crops in Primorskii krai. Russ. J. Plant Physiol. 2003, 50, 112–118. [Google Scholar] [CrossRef]
- Liu, L.; Li, H.; Zeng, H.; Cai, Q.; Zhou, X.; Yin, C. Exogenous jasmonic acid and cytokinin antagonistically regulate rice flag leaf senescence by mediating chlorophyll degradation, membrane deterioration, and senescence-associated genes expression. J. Plant Growth Regul. 2016, 35, 366–376. [Google Scholar] [CrossRef]
- Hakata, M.; Muramatsu, M.; Nakamura, H.; Hara, N.; Kishimoto, M.; Iida-Okada, K.; Kajikawa, M.; Imai-Toki, N.; Toki, S.; Nagamura, Y. Overexpression of TIFY genes promotes plant growth in rice through jasmonate signaling. Biosci. Biotechnol. Biochem. 2017, 81, 906–913. [Google Scholar] [CrossRef]
- Li, Z.; Pinson, S.R.; Stansel, J.W.; Paterson, A.H. Genetic dissection of the source-sink relationship affecting fecundity and yield in rice shape (Oryza sativa L.). Mol. Breed. 1998, 4, 419–426. [Google Scholar] [CrossRef]
- Kobayashi, N.I.; Yamaji, N.; Yamamoto, H.; Okubo, K.; Ueno, H.; Costa, A.; Tanoi, K.; Matsumura, H.; Fujii-Kashino, M.; Horiuchi, T.; et al. OsHKT1;5 mediates Na+ exclusion in the vasculature to protect leaf blades and reproductive tissues from salt toxicity in rice. Plant J. 2017, 91, 657–670. [Google Scholar] [CrossRef] [PubMed]
- Missirian, V.; Henry, I.; Comai, L.; Filkov, V. POPE: Pipeline of parentally-biased expression. In Proceedings of the ISBRA: 2012 Bioinformatics Research and Applications, Dallas, TX, USA, 21–23 May 2012; pp. 177–188. [Google Scholar]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [PubMed]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef] [PubMed]
- Bindea, G.; Mlecnik, B.; Hackl, H.; Charoentong, P.; Tosolini, M.; Kirilovsky, A.; Fridman, W.-H.; Pagès, F.; Trajanoski, Z.; Galon, J. ClueGO: A Cytoscape plug-in to decipher functionally grouped gene ontology and pathway annotation networks. Bioinformatics 2009, 25, 1091–1093. [Google Scholar] [CrossRef] [PubMed]
- Kanehisa, M.; Goto, S. KEGG: Kyoto encyclopedia of genes and genomes. Nucleic Acids Res. 2000, 28, 27–30. [Google Scholar] [CrossRef] [PubMed]
- Lichtenthaler, H.K. Chlorophylls and carotenoids: Pigments of photosynthetic biomembranes. Methods Enzymol. 1987, 148, 350–380. [Google Scholar]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Reproductive Characters | Control * | Salt Stress * | ||||||
|---|---|---|---|---|---|---|---|---|
| WT | TOSL1 | TOSL2 | TOSL3 | WT | TOSL1 | TOSL2 | TOSL3 | |
| Tiller number/pl | 5.75 ± 0.55 ab | 5.25 ± 0.55 abc | 7.00 ± 0.55 a | 5.50 ± 0.55 abc | 3.00 ± 0.55 c | 3.75 ± 0.55 bc | 4.50 ± 0.55 abc | 4.00 ± 0.55 bc |
| Panicle number/pl | 5.25 ± 0.33 a | 3.50 ± 0.33 b | 3.50 ± 0.33 b | 3.00 ± 0.33 bc | 1.75 ± 0.33 c | 2.50 ± 0.33 bc | 2.75 ± 0.33 bc | 2.00 ± 0.33 bc |
| Panicle length | 10.15 ± 0.71 | 9.28 ± 0.71 | 8.87 ± 0.71 | 8.87 ± 0.71 | 7.38 ± 0.71 | 8.27 ± 0.71 | 9.00 ± 0.71 | 9.73 ± 0.71 |
| % Fertility | 65.36 ± 9.52 a | 45.18 ± 9.52 ab | 48.39 ± 9.52 ab | 44.34 ± 9.52 ab | 15.27 ± 9.52 b | 47.03 ± 9.52 ab | 14.17 ± 9.52 b | 36.84 ± 9.52 ab |
| Seeds per plant | 193.50 ± 11.15 a | 108.75 ± 11.15 bc | 130.00 ± 11.15 b | 115.75 ± 11.15 bc | 70.25 ± 11.15 c | 89.75 ± 11.15 bc | 68.50 ± 11.15 c | 84.50 ± 11.15 bc |
| Gene | Primer | Sequence (5′–3′) | Tm (°C) |
|---|---|---|---|
| OsEF-1∝ | EF-1∝-F | ATGGTTGTGGAGACCTTC | 53.7 |
| EF-1∝-R | ATGGTTGTGGAGACCTTC | 58.2 | |
| LOC_Os01g58470 | Os01g58470-F | AGGCATTGATCCTGAGACAG | 54.3 |
| Os01g58470-R | AGAGCAGAATATCCCACTGC | 54.4 | |
| LOC_Os01g64960 | Os01g64960-F | GCATCGCCTTCTCCATCA | 57.1 |
| Os01g64960-R | GAAGACGACGTTGAAGAGGA | 57.3 | |
| LOC_Os03g39610 | Os03g39610-F | GGAGGCGGTGTGGTTCAAGG | 61.0 |
| Os03g39610-R | GCGGTAGCCCTCGACGAATC | 60.3 | |
| LOC_Os04g3383 | Os04g33830-F | CCGTTCTGGCTGTGGTT | 55.4 |
| Os04g33830-R | CGTCCGTACAGTCAAGCTAA | 54.3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Boonchai, C.; Udomchalothorn, T.; Sripinyowanich, S.; Comai, L.; Buaboocha, T.; Chadchawan, S. Rice Overexpressing OsNUC1-S Reveals Differential Gene Expression Leading to Yield Loss Reduction after Salt Stress at the Booting Stage. Int. J. Mol. Sci. 2018, 19, 3936. https://doi.org/10.3390/ijms19123936
Boonchai C, Udomchalothorn T, Sripinyowanich S, Comai L, Buaboocha T, Chadchawan S. Rice Overexpressing OsNUC1-S Reveals Differential Gene Expression Leading to Yield Loss Reduction after Salt Stress at the Booting Stage. International Journal of Molecular Sciences. 2018; 19(12):3936. https://doi.org/10.3390/ijms19123936
Chicago/Turabian StyleBoonchai, Chuthamas, Thanikarn Udomchalothorn, Siriporn Sripinyowanich, Luca Comai, Teerapong Buaboocha, and Supachitra Chadchawan. 2018. "Rice Overexpressing OsNUC1-S Reveals Differential Gene Expression Leading to Yield Loss Reduction after Salt Stress at the Booting Stage" International Journal of Molecular Sciences 19, no. 12: 3936. https://doi.org/10.3390/ijms19123936