Knockout of SlSBPASE Suppresses Carbon Assimilation and Alters Nitrogen Metabolism in Tomato Plants

College of Forestry, Northwest A&F University, Yangling 712100, Shaanxi, China
*
Author to whom correspondence should be addressed.
Int. J. Mol. Sci. 2018, 19(12), 4046; https://doi.org/10.3390/ijms19124046
Submission received: 23 November 2018
/
Revised: 11 December 2018
/
Accepted: 11 December 2018
/
Published: 14 December 2018
(This article belongs to the Section Molecular Plant Sciences)
Abstract
:Sedoheptulose-1,7-bisphosphatase (SBPase) is an enzyme in the Calvin–Benson cycle and has been documented to be important in carbon assimilation, growth and stress tolerance in plants. However, information on the impact of SBPase on carbon assimilation and nitrogen metabolism in tomato plants (Solanum lycopersicum) is rather limited. In the present study, we investigated the role of SBPase in carbon assimilation and nitrogen metabolism in tomato plants by knocking out SBPase gene SlSBPASE using clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (Cas9) gene editing technology. Compared with wild-type plants, slsbpase mutant plants displayed severe growth retardation. Further analyses showed that knockout of SlSBPASE led to a substantial reduction in SBPase activity and as a consequence, ribulose-1,5-bisphosphate (RuBP) regeneration and carbon assimilation rate were dramatically inhibited in slsbpase mutant plants. It was further observed that much lower levels of sucrose and starch were accumulated in slsbpase mutant plants than their wild-type counterparts during the photoperiod. Intriguingly, mutation in SlSBPASE altered nitrogen metabolism as demonstrated by changes in levels of protein and amino acids and activities of nitrogen metabolic enzymes. Collectively, our data suggest that SlSBPASE is required for optimal growth, carbon assimilation and nitrogen metabolism in tomato plants.
1. Introduction
The Calvin–Benson cycle is the primary pathway of photosynthetic carbon fixation in higher plants. The cycle is central to plant metabolism, providing intermediates for biosynthesis of sucrose, starch, isoprenoid and shikimic acid [1]. Within the cycle, there are a total of 11 different enzymes that catalyze 13 reactions, which are divided into three distinct phases, including carboxylation of ribulose-1,5-bisphosphate (RuBP), reduction of 3-phosphoglycerate and regeneration of the CO2 acceptor RuBP [2]. The enzyme sedoheptulose-1,7-bisphosphatase (SBPase) catalyzes the dephosphorylation of sedoheptulose-1,7-bisphosphate to sedoheptulose-7-phosphate, acting in the regeneration of CO2 acceptor molecule RuBP in the Calvin–Benson cycle. In vitro studies have revealed that the activity of SBPase is regulated by a variety of factors, including pH and Mg2+ [3,4]. In addition, like other enzymes in the Calvin-Benson cycle, SBPase is activated via ferredoxin/thioredoxin system in response to light [5,6].
Multiple lines of evidence support that SBPase is a critical enzyme in the regulation of photosynthetic carbon fixation in the Calvin-Benson cycle and is therefore a target to manipulate to improve photosynthetic capacity in plants. In the early work, expression of cyanobacterial FBPase (fructose-1,6-bisphosatase) /SBPase in tobacco plants led to enhanced photosynthetic carbon fixation and biomass [7]. In another study, activity of SBPase was increased by overexpression of an Arabidopsis cDNA in tobacco plants and higher levels of carbohydrates accumulation and biomass were observed in transgenic plants [8]. Later work has shown that overexpression of SBPase improved photosynthetic carbon gain and yield in tobacco plants grown under field conditions [9]. Additionally, analysis of a loss-of-function mutant illustrated that mutation in SBPASE retarded growth and development through inhibition of carbon assimilation in Arabidopsis [10]. More recently, expression of a Brachypodium distachyon SBPase gene in wheat increased SBPase activity and improved photosynthesis and grain yield [11].
Nitrogen is an essential nutrient element for plant growth and development. It is well known that nitrogen is a major constituent of numerous organic compounds, including nucleic acids, amino acids, chlorophyll and phytohormones. Nitrogen assimilation represents a crucial step in the synthesis of organic compounds that require nitrogen and this step involves several enzymes, including nitrate reductase (NR), glutamine synthetase (GS), glutamate synthase (GOGAT) and glutamate dehydrogenase (GDH) [12,13]. It is recognized that nitrogen assimilation is in close association with carbon metabolism and the balance between nitrogen and carbon metabolism is critical for optimal growth and development in plants [14,15]. Nitrate availability affects the expression of a variety of genes encoding enzymes in carbon metabolism, while carbon depletion decreases protein biosynthesis and alters nitrogen metabolism [14,16]. A previous study has concluded that reduced SBPase activity in transgenic rice leads to decreased nitrogen use efficiency [17], however, no further studies have been conducted to investigate the impact of SBPase activity on nitrogen metabolism. Studies using SBPASE knockout mutant may help to gain further insights into the relationship between SBPase activity and nitrogen metabolism in plants.
Genome editing holds great promise for crop improvement. The clustered regularly interspaced short palindromic repeats (CRISPR)/CRISPR-associated protein 9 (CRISPR/Cas9) system is a powerful tool for genome editing. Since the first report of CRISPR/Cas9 editing in plants in 2013, this system has been widely used for genome editing in model plants and crops, including Arabidopsis thaliana, rice, wheat, maize and tomato [18,19,20,21]. CRISPR/Cas9 has become the tool of choice for gene editing due to several advantages, such as high mutation efficiency, rare off-target mutations, stable inheritance, low cost and great flexibility [22]. Tomato (Solanum lycopersicum) is a high-value vegetable crop, making an important part of agricultural industry. Using antisense transgenic plants, we have previously investigated the importance of tomato SBPase gene SlSBPASE (Solyc05g052600) in the photosynthetic carbon assimilation and growth in tomato plants [23]. However, studies using partial-loss/gain-of-function transgenic plants may yield conflicting results due to the inconsistent expression of target genes within or among generations. In addition, the absence of SlSBPASE knockout mutant hampers us from making a thorough functional analysis of this gene. In the present study, CRISPR/Cas9 system was utilized to generate SlSBPASE knockout mutant plants, which allowed us to better define the functions of SBPase in tomato plants. We were in an attempt to investigate the role for SBPase in the regulation of carbon assimilation and nitrogen metabolism by measuring photosynthetic carbon fixation, carbohydrate accumulation, protein content and enzyme activities related to nitrogen metabolism in slsbpase mutant plants and their wild-type counterparts.
2. Results
2.1. Generation of Loss-of-Function Mutant of SlSBPASE Using CRISPR/Cas9 Gene-Editing System
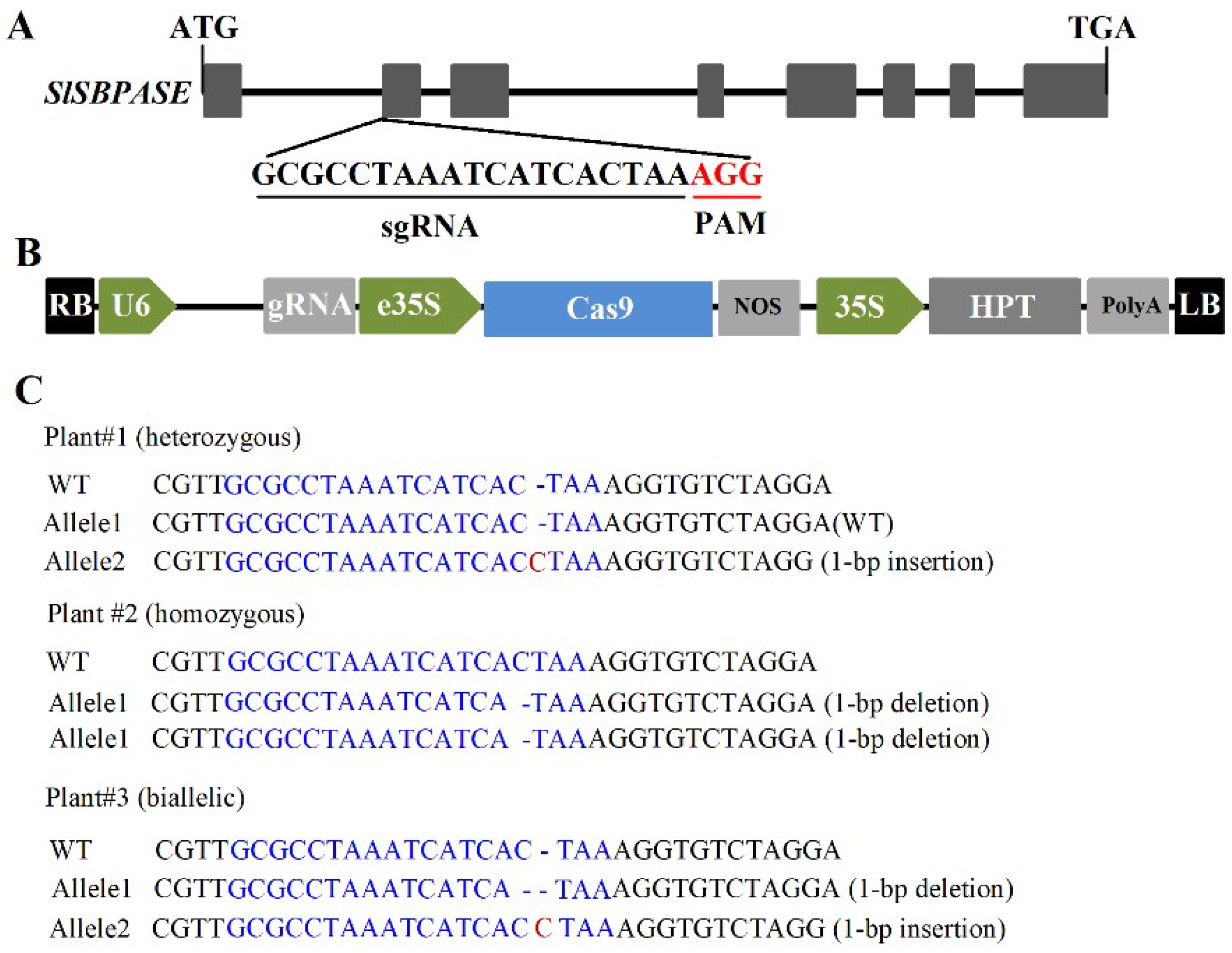
SBPase acts in the regenerative phase of the Calvin–Benson cycle and is important in the control of carbon flux in plants. Our previous studies on SBPase using either SBPase overexpressing or antisense transgenic tomato plants have exhibited a critical role of SBPase in photosynthetic carbon assimilation [23,24]. In tomato plants, SBPase is encoded by one gene SlSBPASE, the transcript of which is most abundant in leaf [23]. In order to provide further evidence for the role of SBPase in the regulation of carbon fixation, we generated stable loss-of-function slspbase mutants using CRISPR/Cas9 gene-editing system. A single guide RNA (sgRNA, 5′-GCGCCTAAATCATCACTAA-3′) was designed to specifically target the second exon of SlSBPASE (Figure 1A). We cloned the sgRNA sequence into a binary vector that contains sgRNA and Cas9 expression cassettes and the resulting construct was transformed into wild-type tomato plants.
T0 transgenic plants were genotyped by sequencing PCR products from genomic DNA flanking the target site and mutant plants were confirmed. In these mutant plants, biallelic, heterozygous and homozygous mutations were found and two of the plants carried the same homozygous 1-bp deletion in the second exon of SlSBPASE (Figure 1C). Homozygous T1 plants were selected for further analyses in this study and no off-target mutations in these plants were found in the three predicted off-target sites.
2.2. Characterization of slsbpase Mutant Plants
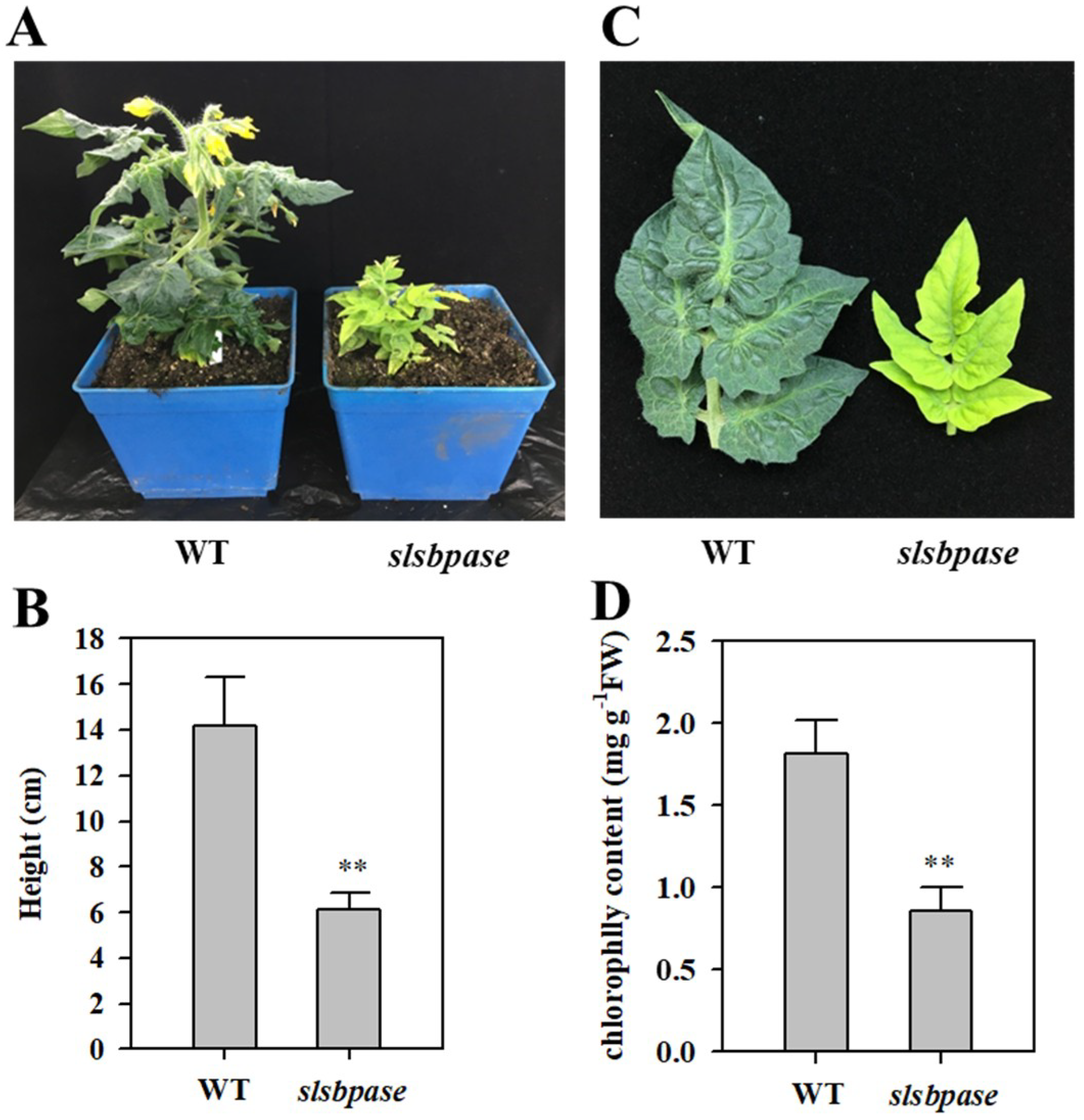
Growth of slsbpase mutant plants was dramatically inhibited compared with that of wild-type plants at both vegetative and reproductive stages. The height and the number of leaves were substantially reduced by mutation of SlSBPASE (Figure 2A,B). The number of flowers in slsbpase mutant plants was also decreased. Moreover, slsbpase mutants exhibited a leaf chlorotic phenotype (Figure 2C), with the content of chlorophyll being substantially reduced (Figure 2D). In spite of severe retardation of growth, slsbpase mutants were still able to flower and set fruits under optimal growth conditions.
2.3. Mutation of SlSBPASE Causes Severe Reductions in SBPase Activity, RuBP Regeneration and CO2 Assimilation Rate
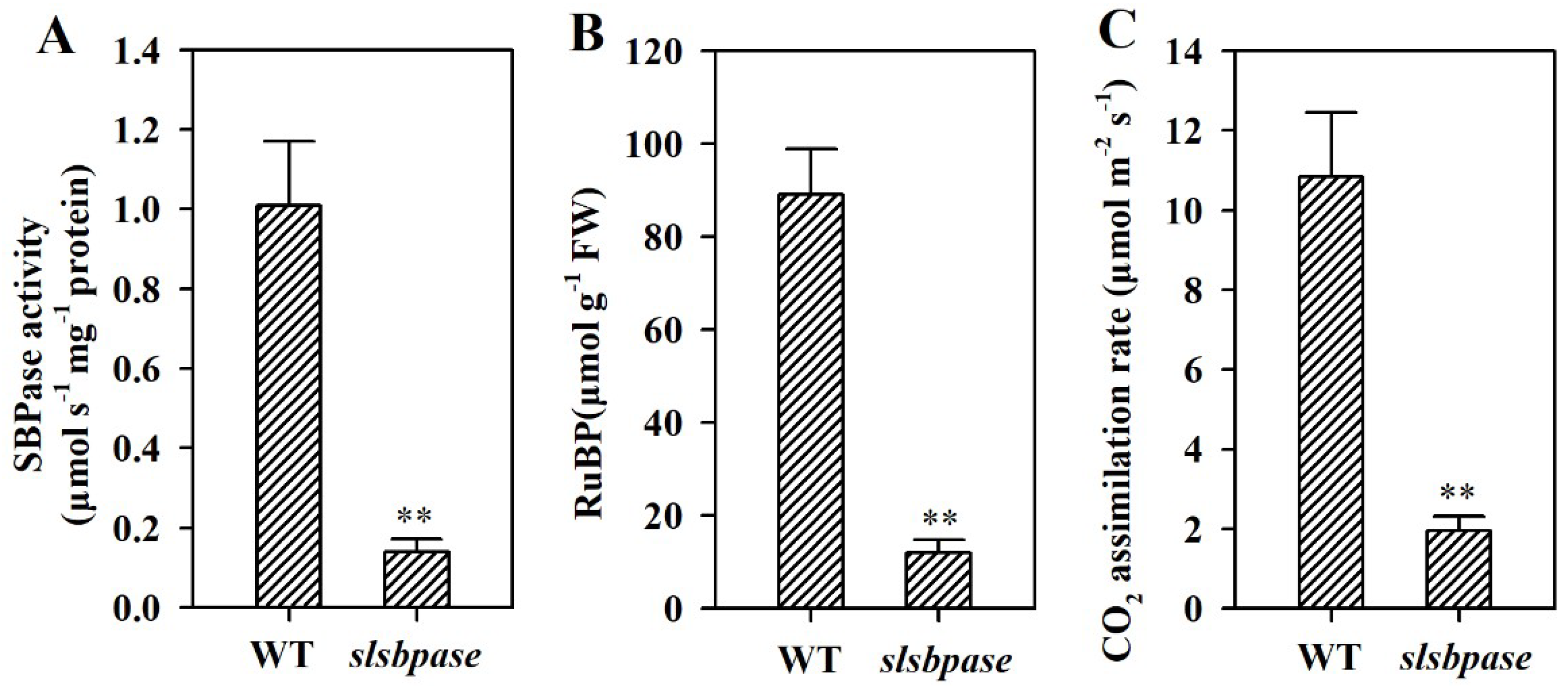
SBPase is thought to be responsible for the regeneration of RuBP and affects the photosynthetic carbon fixation [25]. To validate the relationship between mutation of SlSBPASE and reductions in SBPase activity and CO2 assimilation rate, we first measured SBPase activity of slsbpase mutants. A low level of SBPase activity was detected in the leaves of mutant plants. SBPase activity in slsbpase mutant leaves was reduced to 11% of that in wild-type leaves (Figure 3A). Given that SlSBPASE was mutated, this result came as unexpected. It is likely that this small amount of activity was caused by non-specific activity of other proteins in the measurement of SBPase activity. Similar result of trace SBPase activity was also observed in an Arabidopsis mutant with loss of function of SBPASE [10].
SBPase functions in the regenerative stage and is crucial for the regeneration of CO2 acceptor molecule RuBP in the Calvin–Benson cycle. We therefore determined the accumulation of RuBP in slsbpase mutants to examine the effect of decreased SBPase activity on the regeneration of RuBP. In this study, it was observed that RuBP accumulation in slsbpase mutant plants was dramatically reduced compared with that in wild-type plants (Figure 3B), suggestive of the key role for SBPase in controlling RuBP regeneration.
To investigate the role of SBPase in carbon assimilation, we then measured CO2 assimilation rate of slsbpase mutants. It was found that CO2 assimilation was substantially inhibited, with carbon assimilation rate in slsbpase mutant being reduced to 18% of that in their wild-type counterparts (Figure 3C). This observation is consistent with a previous report that decreases in SBPase activity result in significant reductions in photosynthesis in transgenic tobacco plants [26,27].
2.4. Mutation of SlSBPASE Dramatically Inhibits Biosynthesis of Sucrose and Starch
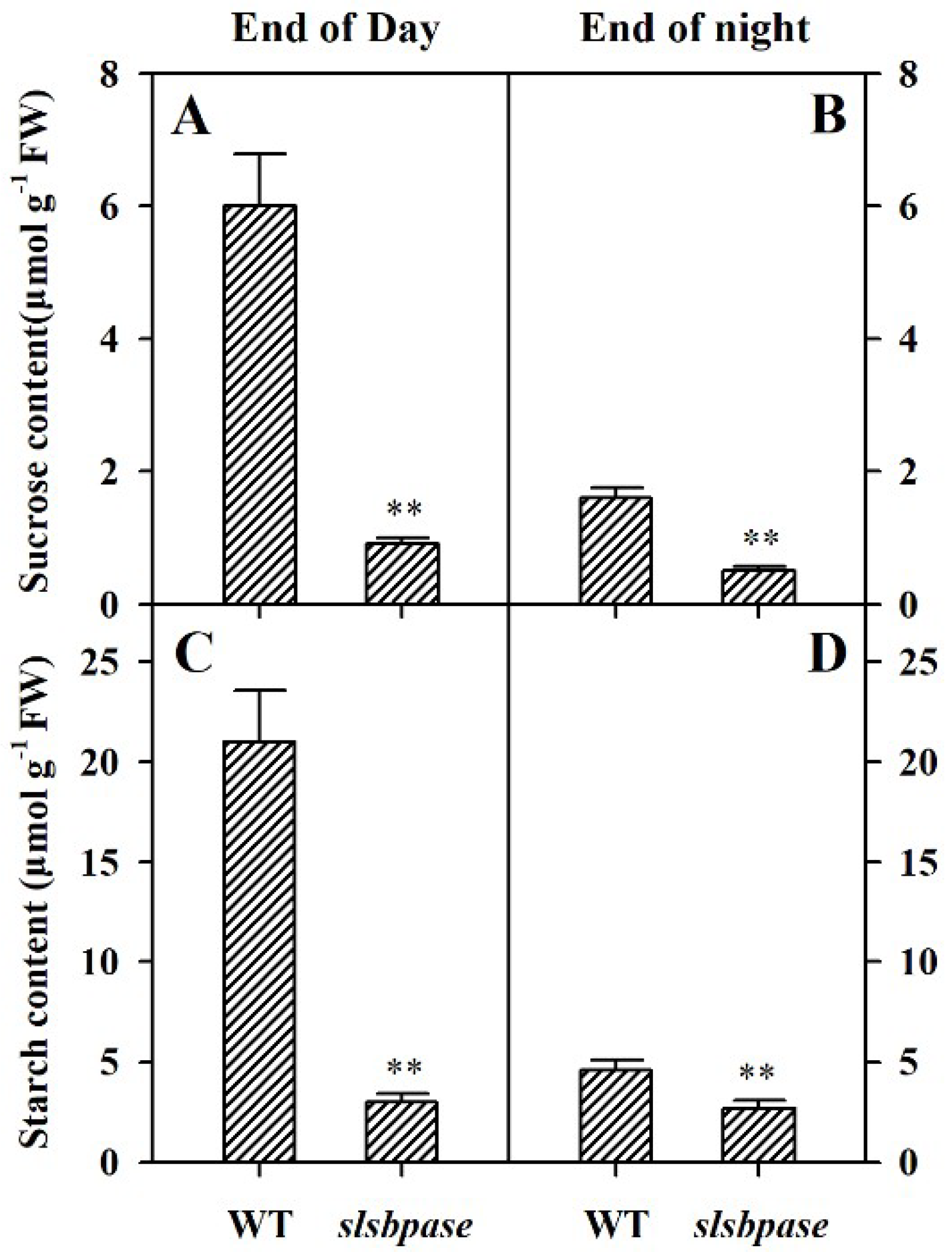
To examine the effect of the SlSBPASE mutation on the accumulation of carbohydrates in slsbpase mutant plants, in parallel with photosynthetic analysis, we measured the contents of sucrose and starch in the mutant and wild-type plants at the end of the light period and at the end of dark period (16-h light/8-h dark). In both the mutant plants and their wild-type counterparts, sucrose and starch accumulated during the light period. However, the mutant plants accumulated substantially less sucrose and starch than wild-type plants (Figure 4A,C), in agreement with the role of SBPase in photosynthetic carbon fixation. At the end of dark period, levels of sucrose and starch were low in both slsbpase mutant and wild-type plants (Figure 4B,D), demonstrating that the carbohydrates accumulated in the light were being remobilized at night.
2.5. Mutation of SlSBPASE Reduces Night Respiration Rate
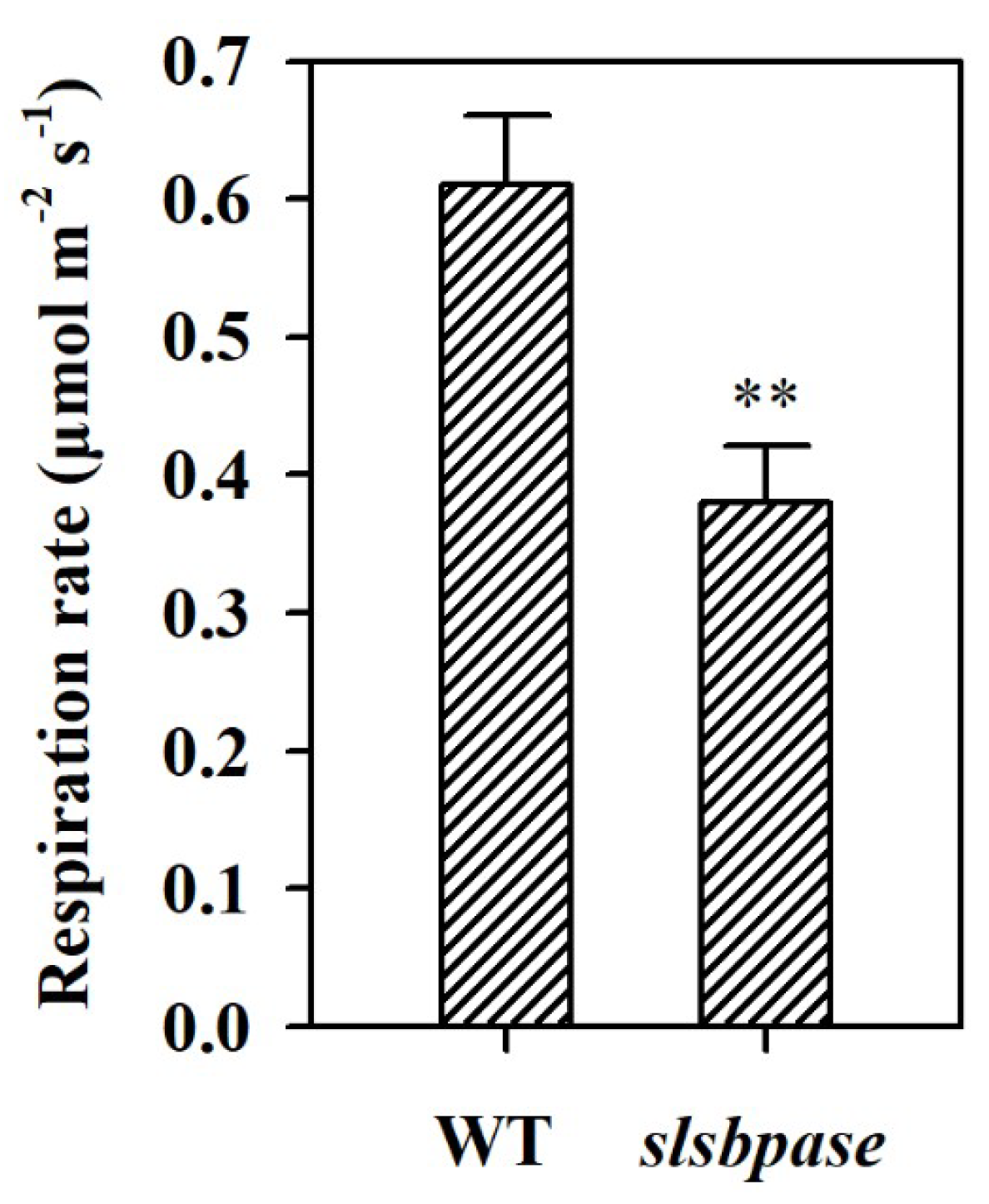
In order to further understand the difference in the accumulation of starch between wild-type and slsbpase mutant plants, we measured the night respiration rate. It was observed that respiration rate was decreased in slsbpase mutant plants at night compared with that in wild-type plants (Figure 5). This result was consistent with our observation that starch was reduced by ~10% in slsbpase mutant plants, while starch was decreased by ~78% in wild-type plants at the end of night.
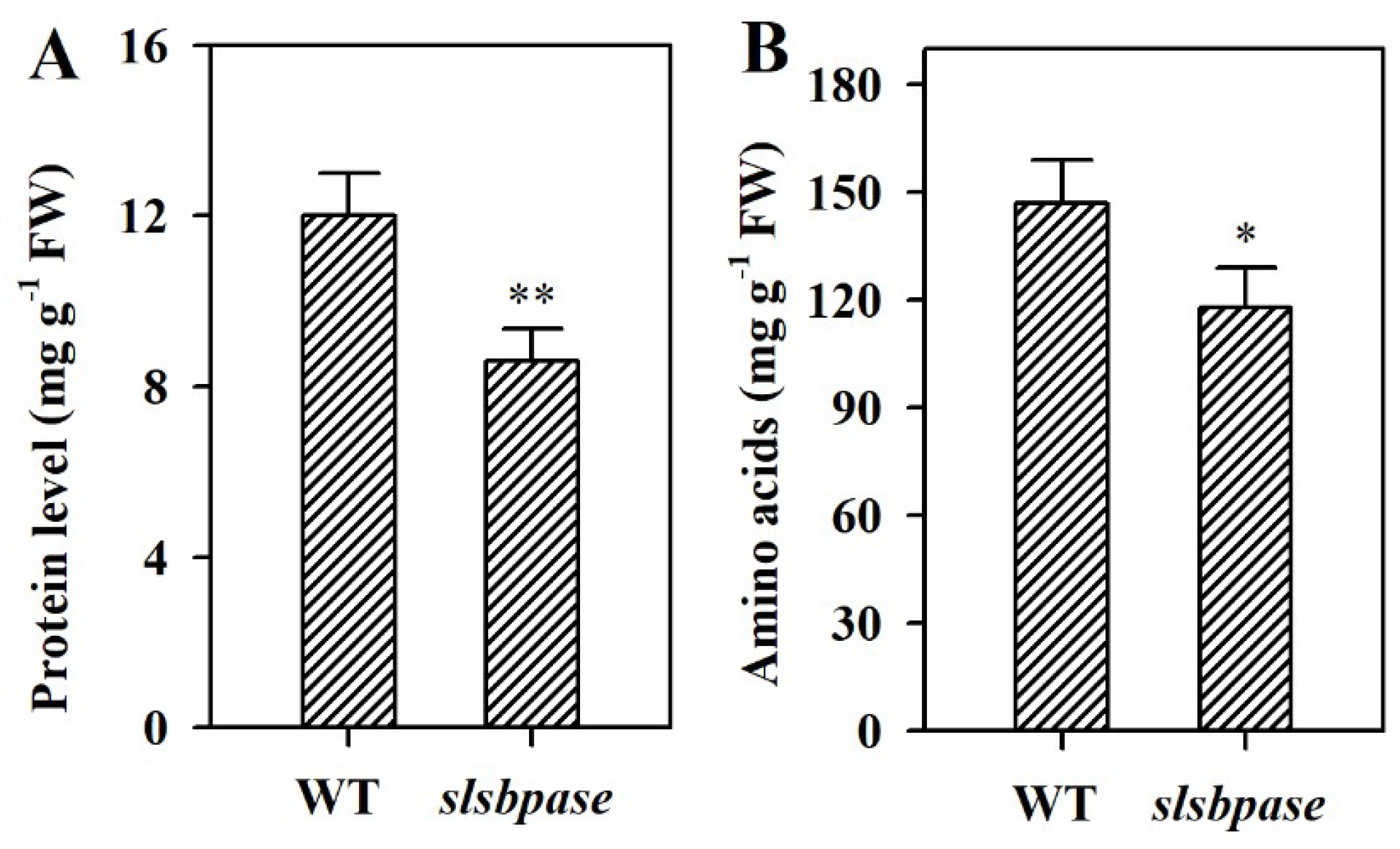
2.6. Mutation of SlSBPASE Decreases Levels of Protein and Amino Acids in Tomato Leaves
To examine the impact of SlSBPASE mutation on nitrogen metabolism, we measured the contents of protein and amino acids in slsbpase mutant plants and their wild-type counterparts. In slsbpase mutant leaves, total protein was decreased by ~30% compared with wild-type leaves (Figure 6A). Similar to protein level, level of amino acids was reduced as a consequence of SlSBPASE mutation. Level of amino acids was reduced by ~14% in slsbpase mutant plants compared with their wild-type counterparts (Figure 6B). The decreased levels of protein and amino acids suggest that mutation in SlSBPASE leads to alterations in nitrogen metabolism.
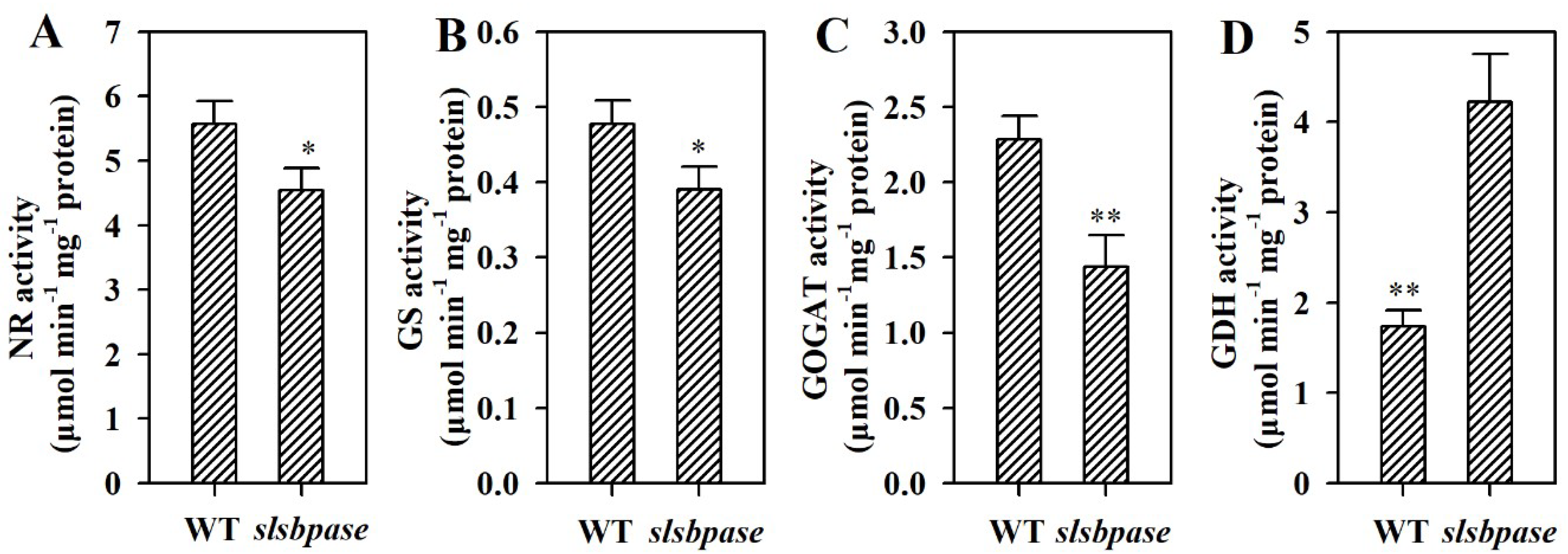
2.7. Mutation of SlSBPASE Alters Activities of Enzymes Involved in Nitrogen Metabolism
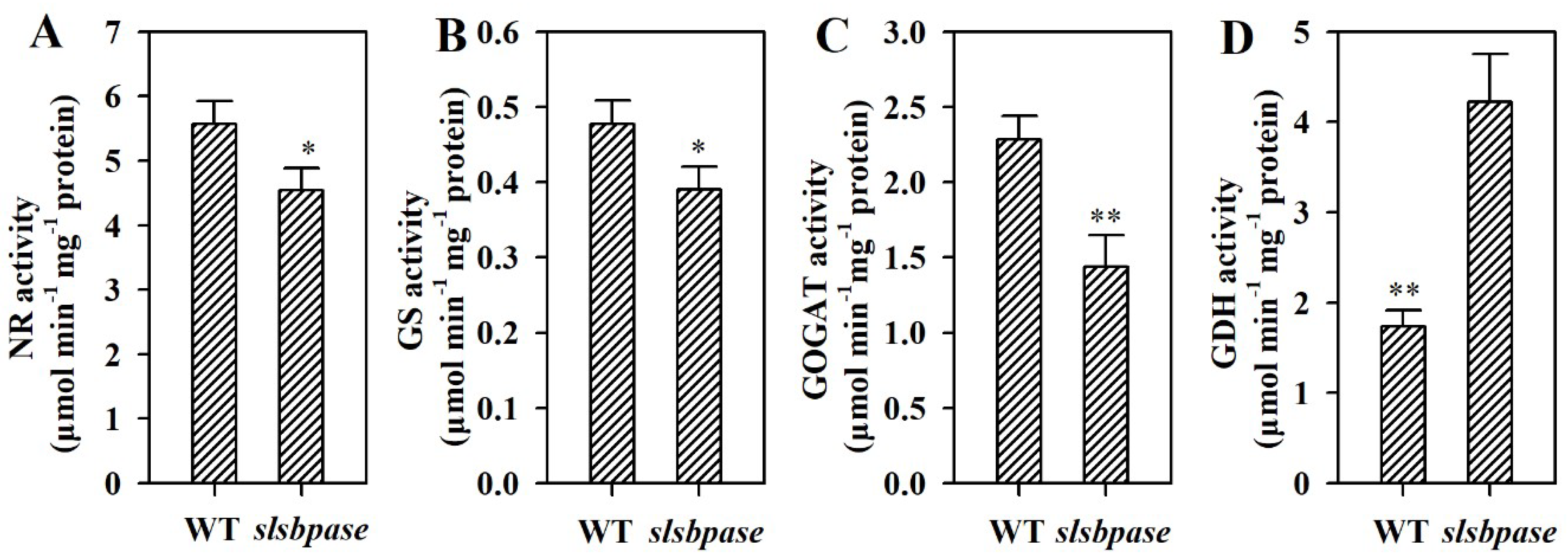
To gain further insights into the impact of mutagenesis of SBPase gene on nitrogen metabolism, we measured activities of nitrate reductase (NR), glutamine synthetase (GS), glutamate synthase (GOGAT) and glutamate dehydrogenase (GDH), which are key enzymes in the nitrogen metabolism. Mutation in SlSBPASE led to reductions in the activities of NR, GS and GOGAT in tomato leaves (Figure 7A–C). Contrary to NR, GS and GOGAT, GDH activity was significantly enhanced in slsbpase mutant leaves compared with wild-type leaves (Figure 7D).
3. Discussion
SBPase has been demonstrated to be an important enzyme mediating the carbon flux in the Calvin–Benson cycle. Previous studies have produced transgenic plants displaying a range of SBPase activities to investigate the role of SBPase in photosynthetic capacity, growth and tolerance to abiotic stresses [8,11,23,24,26,28,29,30,31,32]. In this study, we generated loss-of-function mutants of SlSBPASE using CRISPR/Cas9 gene-editing system in tomato plants to confirm the function of SBPase in photosynthetic carbon assimilation and growth and to examine the impact of SlSBPASE mutation on nitrogen metabolism. We concluded that SBPase plays a critical role in growth and carbon assimilation and suppression of SBPase activity alters nitrogen metabolism in tomato plants. The evidence supporting this conclusion includes that (1) CRISPR/Cas9-mediated mutagenesis of SlSBPASE led to the phenotype of dramatic growth retardation; (2) slsbpase mutant plants exhibited a substantial reduction in SBPase activity, RuBP regeneration and carbon assimilation rate; (3) mutation of SlSBPASE decreased the accumulation of carbohydrates; (4) mutation of SlSBPASE altered levels of protein and amino acids and activities of key metabolic enzymes in nitrogen metabolism.
The Calvin–Benson cycle is the primary photosynthetic carbon assimilation pathway and can be limited by several enzymes, including Rubisco, fructose-1,6-bisphosatase (FBPase), SBPase and phosphoribulokinase (PRKase). SBPase functions in the regenerative phase of the Calvin–Benson Cycle, catalyzing the irreversible dephosphorylation of sedoheptulose-1,7-biphosphate to sedoheptulose-7-phosphate. SBPase has been proved to exert control over flux of carbon through the cycle. Given the importance of SBPase, it has been extensively studied using transgenic lines of both model and crop plants. In tobacco plants with decreased SBPase activity, photosynthetic capacity and growth were reduced [25,28]. On the contrary, elevated levels of SBPase resulted in increased carbon assimilation rates and biomass in tobacco plants [8,9]. These results were also observed in transgenic crops with different levels of SBPase, such as tomato, rice and wheat [11,17,23,30]. To provide additional and direct evidence for the role of SBPase, in this work, we generated loss-of-function mutants of SlSBPASE using CRISPR/Cas9 gene-editing system, which has proved powerful and efficient in knocking out genes of interest. The slsbpase mutant plants displayed the phenotype of severe growth retardation, with plant size being reduced and the number of leaves decreased, supporting the notion that SlSBPASE is required for normal growth in tomato plants.
It was observed that slsbpase mutant plants had a chlorotic leaf phenotype. Similar phenotype was also observed in sbpase mutant plants of Arabidopsis [10]. One possible explanation of leaf chlorosis is that SlSBPASE mutation led to the inhibition of chloroplast biogenesis, which may result from the limited carbon reserve such as starch. The other explanation is that mutation in SlSBPASE might accelerate the degradation of chlorophyll as loss of SlSBPASE was previously demonstrated to trigger premature leaf senescence [33]. Typically, degradation of chlorophyll was closely associated with leaf senescence.
SBPase plays an essential role in the regeneration of RuBP, which is the CO2 acceptor and, in association with Rubisco activase, is crucial for carbon assimilation by affecting the activation state of Rubisco [34]. The regenerative capacity for RuBP in the Calvin-Benson cycle is largely dependent on SBPase activity. It has been illustrated that RuBP regeneration decreases linearly with reduced SBPase activity in tobacco plants [25]. Consistently, in this study, we observed that mutation of SlSBPASE led to severe reduction in SBPase activity, which consequently decreased RuBP accumulation and carbon assimilation rate. This observation supports the critical of SBPase in retaining photosynthetic carbon assimilation in tomato plants.
Though growth was severely retarded, slsbpase mutant plants were still able to survive and produce seeds. Given that RuBP regeneration is indispensable for carbon fixation, there may be an enzyme that possesses catalytic activity of SBPase, catalyzing the dephosphorylation of sedoheptulose-1,7-biphosphate to sedoheptulose-7-phosphate at a very low efficiency in slsbpase mutant plants. Chloroplast FBPase might be a candidate enzyme because FBPase and SBPase have similar architecture and their regulatory and catalytic properties resemble. In addition, there are also some amino acids that are conserved in the substrate binding domain of both enzymes [35]. However, it must be noted that the conclusion from a recent study refutes FBPase as a possible substitute. Gütle et al. claimed that SBPase has a larger substrate binding site than FBPase, accommodating both seven- and six-carbon sugar phosphate substrates, while FBPase is active only with six-carbon substrates [36]. In this scenario, there may exist a third enzyme for the dephosphorylation of sedoheptulose-1,7-biphosphate to sedoheptulose-7-phosphate operating inefficiently. It is also possible that an unknown mechanism is present in plants, which might bypass the pathway of sedoheptulose-1,7-biphosphate and aids in the minimum regeneration of RuBP to guarantee the survival of plants in the absence of enough SBPase.
The rate of plant growth partly depends on the efficiency of photosynthetic carbon assimilation. In the present work, the retarded growth was observed concurrent with suppressed carbon assimilation rate in SlSBPASE-knockout plants. Analysis of carbohydrate accumulation during the light period showed that levels of sucrose and starch were much reduced in slsbpase mutant plants in comparison with those in wild-type plants. Thus, there was evident correlation between retarded growth, reduced SBPase activity, suppressed photosynthesis and decreased accumulation of sucrose and starch. However, at the end of dark period, accumulation of carbohydrate was reduced to a low level in wild-type plants, suggesting carbohydrates produced in the light period were being used possibly for growth in the dark period. As such, the difference in the consumption of carbohydrates in the dark between mutant and wild-type plants may explain, in part, the growth difference between them.
It has been established that carbon metabolism and nitrogen metabolism are interrelated in plants [14,15,16]. In this study, we found that mutation in SlSBPASE severely reduced SBPase activity, leading to suppressed carbon assimilation, which further altered nitrogen metabolism, as demonstrated by reduced levels of proteins and amino acids and altered activities of nitrogen metabolic enzymes in slsbpase mutant plants. The enzymes NR, GS and GOGAT are required for the assimilation of inorganic nitrogen. It was observed that the activities of these enzymes were markedly decreased by mutation of SlSBPASE, implying that nitrogen assimilation was reduced as a consequence of suppressed SBPase. Nitrogen assimilation is among the most energy-intensive reactions. It is estimated that the energy needed for assimilating each NO3− is equivalent to 12 ATP (adenosine triphosphate), while most biochemical reactions consume the energy of one or two ATP [37]. We thus reasoned that downregulation of nitrogen assimilation may be a general response to reduced energy reserve in slsbpase mutant plants. GDH (glutamate dehydrogenase) has been considered as a metabolic indicator of carbon deficiency [16]. During carbon shortage, GDH acts in the direction of releasing amino nitrogen and recycles glutamate to 2-oxoglutarate for the tricarboxylic acid (TCA) cycle [38,39]. Consistently, we observed that in contrast to NR, GS and GOGAT, GDH activity was significantly increased by knockout of SlSBPASE, demonstrating the increased nitrogen catabolism in slsbpase mutant plants. These results support that slsbpase mutant plants are able to adjust central nitrogen metabolism in response to carbon deficiency caused by loss of SBPase activity.
In summary, we generated a loss-of-function mutant of SlSBPASE using CRISPR/Cas9 mediated gene-editing system. We have demonstrated that SlSBPASE is required for optimal growth, carbon assimilation and nitrogen metabolism in tomato plants. Mutation in SlSBPASE leads to reduced photosynthesis, decreased carbohydrate accumulation, altered nitrogen metabolism and retarded growth in slsbpase mutant plants. Our study provides evidence that SBPase, as an individual enzyme in the Calvin–Benson cycle, not only exerts control over carbon assimilation but also impacts nitrogen metabolism in plants.
4. Materials and Methods
4.1. Plant Materials
Tomato (Solanum lycopersicum cv. Micro-Tom) slsbpase mutant and wild-type seeds were sterilized and germinated at 25 °C in the dark on filter paper in petri dishes. Germinated seeds were then planted individually in 12 cm × 12 cm × 10 cm plastic pots filled with peat and vermiculite (3/1 v/v). Plants were grown under following conditions: 380 μmol mol−1 of CO2, photon flux density of 300 μmol m−2s−1, day/night temperature of 25/20 °C, relative humidity of 60% and a photoperiod of 16 h.
4.2. Selection of Target Sequence and Plasmid Construction
A target sequence (5′-TGCGCCTAAATCATCACTAAAGG-3′) in the second exon of SLSBPASE (Solyc05g052600) was selected using CRISPR online tool (available online: http://skl.scau.edu.cn/targetdesign/) [40]. The CRISPR/Cas9-SlSBPASE vector was constructed using a kit (BGK01) according to the manufacturer’s instructions (BIOGLE, Changzhou, Jiangsu, China). A 19-bp sgRNA oligo (5′-GCGCCTAAATCATCACTAA-3′) was introduced into the expression cassette, including chimeric RNA driven by the AtU6 promoter and optimized Cas9 driven by the enhanced CaMV 35S promoter.
4.3. Agrobacterium Tumefaciens-Mediated Transformation
The CRISPR/Cas9-SlSBPASE-expressing plasmid was introduced into Agrobacterium tumefaciens strain EHA105 (Waryong, Beijing, China). Transgenic tomato plants were produced using the Agrobacterium tumefaciens-mediated transformation method [41]. Transgenic plants were selected by hygromycin resistance and were maintained in a growth chamber at 25 °C with a photoperiod of 16 h light and 8 h dark.
4.4. Mutation Analysis of Transgenic Lines
The genomic DNA was extracted from leaves of transgenic plants using the DNeasy® Plant Mini Kit (TIANGEN Biotech Co. Ltd., Beijing, China) and was used as template to amplify SlSBPASE fragment using primers ATGGATTTGGCATAGCCTAGTAGCT (Forward) and ATCATTAACCTGATTAACCCCTTAT (Reverse) by PCR. The PCR products were sequenced and the sequencing chromatograms were analyzed using a web-based tool DSDecode (available online: http://skl.scau.edu.cn/dsdecode/) [40].
4.5. Off-Target Analysis
Potential off-target sites were predicted using a web-based tool (available online: http://skl.scau.edu.cn/offtarget/) [40]. Three potential off-target sites were selected (Table 1). The genomic DNA was extracted from leaves of T1 transgenic tomato plants and was used to amplify fragments surrounding the potential off-target sites using specific primers (Table 1). The PCR products were sequenced and analyzed.
4.6. Activities of SBPase, NR, GS, GOGAT and GDH
SBPase activity was measured according to the protocol of Harrison et al. [26]. Leaf samples of mutant and wild-type plants were thoroughly ground in liquid nitrogen and then transferred to 1 mL extraction buffer containing 50 mM Hepes, pH 8.2; 5 mM MgCl2; 1 mM EDTA (ethylenediaminetetraacetic acid); 1 mM EGTA (ethylene glycol-bis(β-aminoethyl ether)-N,N,N′,N′-tetraacetic acid); 10% glycerol; 2 mM benzamidine; 2 mM amino caproic acid; 0.5 mM phenylmethylsulfonyluoride (PMSF); 10 mM dithiothreitol (DTT). The supernatant after centrifuging was collected and passed through a NAP-10 column (GE Healthcare Life Sciences, Pittsburgh, PA, USA) equilibrated with desalting buffer. For the assay, 20 μL of protein samples was added to 80 μL of assay buffer (50 mM Tris, pH 8.2; 15 mM MgCl2; 1.5 mM EDTA; 10 mM DTT; 2 mM sedoheptulose-7-phosphate (SBP) and incubated at 25 °C for 5 min. The reaction was stopped by adding 50 μL of 1 M perchloric acid. The samples were then centrifuged for 5 min and the supernatant assayed for phosphate. Fifty microliter samples and phosphate standards (0–0.5 mM NaH2PO4) were incubated with 850 μL molybdate solution (0.3% ammonium molybdate in 0.55 M H2SO4) for 10 min at room temperature. Malachite green (0.035% malachite green, 0.35% polyvinyl alcohol) was added (150 μL) and the samples incubated for a further 45 min at room temperature. Absorbance at 620 nm was measured and used for calculation of SBPase activity.
Nitrate reductase (NR) was extracted and measured using a Nitrate Reductase (NR) Assay Kit (BC0080, Solarbio, Beijing, China). Briefly, 0.1 g leaf samples were extracted in 1 ml extraction solution and the mixture was centrifuged at 4000 g for 10 min. The resulting supernatant was collected for further analysis. The absorbance at 520 nm was used for the calculation of NR activity.
Glutamine synthetase (GS) was extracted and measured using a Micro Glutamine Synthetase (GS) Assay Kit (BC0915, Solarbio, Beijing, China). Briefly, 0.1 g leaf samples were thoroughly ground in liquid nitrogen and extracted with 1 mL extraction buffer. The mixture was centrifuged at 8000 g at 4 °C for 10 min. The supernatant after centrifuging was collected for activity measurement. The absorbance at 520 nm was used for the calculation of GS activity.
Glutamate synthase was extracted and measured using a Glutamate Synthase (GOGAT) Assay Kit (BC0070, Solarbio, Beijing, China). Briefly, 0.1 g leaf samples were extracted in 1 mL extraction buffer. The extraction mixture was centrifuged at 10,000 g at 4 °C for 10 min. The resulting supernatant was harvested and the absorbance at 340 nm was measured for the calculation of GOGAT activity.
Glutamate dehydrogenase Glutamate Dehydrogenase (GDH) Assay Kit (BC1460, Solarbio, Beijing, China). In brief, 0.1 g leaf samples were extracted in 1 mL GDH extraction buffer. The extraction mixture was centrifuged at 8000 g at 4 °C for 10 min. The resulting supernatant was collected for further analysis. The absorbance at 340 nm was measured for the calculation of GDH activity.
4.7. Measurement of Carbon Assimilation Rate and Night Respiration Rate
Carbon assimilation rate and night respiration rate were measured with a photosynthesis system LI-6400XT (LI-COR Biosciences, Lincoln, NE, USA). The measurements were made on young fully expanded leaves of slsbpase and wild-type plants.
4.8. Determination of RuBP Level
Fresh leaf samples (0.1 g) were ground with 1 ml of 5% HClO4 and the level of phosphorylated RuBP was determined enzymatically as described by Sicher et al. [42].
4.9. Determination of Carbohydrate Level
4.10. Measurements of Protein and Amino Acids
Leaf samples were collected from slsbpase mutant plants and wild-type plants. Total protein was extracted using a Plant Total Protein Extraction Kit (Solarbio, Beijing, China) and quantified by Bradford assay [45]. Amino acids were extracted and measured using a kit (BC1575, Solarbio, Beijing, China)
4.11. Statistical Analysis
Experiments in this study were repeated three times and the values presented are the means ± SDs. Student’s t test was performed to compare the difference between wild-type plants and mutant plants. Asterisks indicate that mean values are significantly different at p < 0.05 or p < 0.01 between wild-type and slsbpase mutant plants.
Author Contributions
F.D. and S.Z. designed the study. F.D., Q.H. and M.W. conducted the experiments and analyzed the data. F.D. wrote the manuscript. S.Z. supervised the project and edited the manuscript.
Funding
This research was funded by grants from the Ministry of Science and Technology of China (2015BAD07B05) and the China Postdoctoral Science Foundation (2018M633593).
Conflicts of Interest
The authors declare no conflict of interest.
References
- Geiger, D.R.; Servaites, J.C. Diurnal regulation of photosynthetic carbon metabolism in C3 plants. Annu. Rev. Plant Biol. 1994, 45, 235–256. [Google Scholar] [CrossRef]
- Raines, C.A.; Lloyd, J.C.; Dyer, T.A. New insights into the structure and function of sedoheptulose-1,7-bisphosphatase; an important but neglected Calvin cycle enzyme. J. Exp. Bot. 1999, 50, 1–8. [Google Scholar] [Green Version]
- Laing, W.A.; Stitt, M.; Heldt, H.W. Control of CO2 fixation. Changes in the activity of ribulosephosphate kinase and fructose- and sedoheptulose-bisphosphatase in chloroplasts. BBA Bioenerg. 1981, 637, 348–359. [Google Scholar] [CrossRef]
- Woodrow, I.E.; Murphy, D.J.; Latzko, E. Regulation of stromal sedoheptulose 1,7-bisphosphatase activity by pH and Mg2+ concentration. J. Biol. Chem. 1984, 259, 3791–3795. [Google Scholar]
- Breazeale, V.D.; Buchanan, B.B.; Wolosiuk, R.A. Chloroplast sedoheptulose-1,7-bisphosphatase: Evidence for regulation by the ferredoxin/thioredoxin system. Z. Naturforsch. C 1978, 33, 521–528. [Google Scholar] [CrossRef]
- Wirtz, W.; Stitt, M.; Heldt, H.W. Light activation of calvin cycle enzymes as measured in pea leaves. FEBS Lett. 1982, 142, 223–226. [Google Scholar] [CrossRef] [Green Version]
- Miyagawa, Y.; Tamoi, M.; Shigeoka, S. Overexpression of a cyanobacterial fructose-1,6-/sedoheptulose-1,7-bisphosphatase in tobacco enhances photosynthesis and growth. Nat. Biotechnol. 2001, 19, 965–969. [Google Scholar] [CrossRef]
- Lefebvre, S.; Lawson, T.; Fryer, M.; Zakhleniuk, O.V.; Lloyd, J.C.; Raines, C. Increased sedoheptulose-1, 7-bisphosphatase activity in transgenic tobacco plants stimulates photosynthesis and growth from an early stage in development. Plant Physiol. 2005, 138, 451–460. [Google Scholar] [CrossRef]
- Rosenthal, D.M.; Locke, A.M.; Khozaei, M.; Raines, C.A.; Long, S.P.; Ort, D.R. Over-expressing the C3 photosynthesis cycle enzyme sedoheptulose-1-7 bisphosphatase improves photosynthetic carbon gain and yield under fully open air CO2 fumigation (FACE). BMC Plant Biol. 2011, 11, 123. [Google Scholar] [CrossRef]
- Liu, X.L.; Yu, H.D.; Guan, Y.; Li, J.K.; Guo, F.Q. Carbonylation and loss-of-function analyses of SBPase reveal its metabolic interface role in oxidative stress, carbon assimilation and multiple aspects of growth and development in Arabidopsis. Mol. Plant 2012, 5, 1082–1099. [Google Scholar] [CrossRef]
- Driever, S.M.; Simkin, A.J.; Alotaibi, S.; Fisk, S.J.; Madgwick, P.J.; Sparks, C.A.; Jones, H.D.; Lawson, T.; Parry, M.A.J.; Raines, C.A. Increased SBPase activity improves photosynthesis and grain yield in wheat grown in greenhouse conditions. Philos. Trans. R. Soc. B 2017, 372, 20160384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stitt, M.; Matt, P.; Gibon, Y. Steps towards an integrated view of nitrogen metabolism. J. Exp. Bot. 2002, 53, 959–970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabuchi, M.; Abiko, T.; Yamaya, T. Assimilation of ammonium ions and reutilization of nitrogen in rice (Oryza sativa L.). J. Exp. Bot. 2007, 58, 2319–2327. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M. Nitrate regulation of metabolism and growth. Curr. Opin. Plant Biol. 1999, 2, 178–186. [Google Scholar] [CrossRef]
- Gao, L.; Lu, Z.; Ding, L.; Guo, J.; Wang, M.; Ling, N.; Guo, S.; Shen, Q. Role of aquaporins in determining carbon and nitrogen status in higher plants. Int. J. Mol. Sci. 2018, 19, 35. [Google Scholar] [CrossRef] [PubMed]
- Gibon, Y.; Pyl, E.T.; Sulpice, R.; Lunn, J.E.; HÖhne, M.; GÜnther, M.; Stitt, M. Adjustment of growth, starch turnover, protein content and central metabolism to a decrease of the carbon supply when Arabidopsis is grown in very short photoperiods. Plant Cell Environ. 2009, 32, 859–874. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Li, H.; Jiao, J.; Li, D.; Zhou, L.; Wan, J.; Li, Y. Reduction in SBPase activity by antisense RNA in transgenic rice plants: Effect on photosynthesis, growth and biomass allocation at different nitrogen levels. J. Plant Biol. 2009, 52, 382–394. [Google Scholar] [CrossRef]
- Alagoz, Y.; Gurkok, T.; Zhang, B.; Unver, T. Manipulating the biosynthesis of bioactive compound alkaloids for next-generation metabolic engineering in opium poppy using CRISPR-Cas 9 genome editing technology. Sci. Rep. 2016, 6, 1–9. [Google Scholar] [CrossRef]
- Liu, X.; Wu, S.; Xu, J.; Sui, C.; Wei, J. Application of CRISPR/Cas9 in plant biology. Acta Pharm. Sin. B 2017, 7, 292–302. [Google Scholar] [CrossRef] [Green Version]
- Wang, L.; Chen, L.; Li, R.; Zhao, R.; Yang, M.; Sheng, J.; Shen, L. Reduced drought tolerance by CRISPR/Cas9-mediated SlMAPK3 mutagenesis in tomato plants. J. Agric. Food Chem. 2017, 65, 8674–8682. [Google Scholar] [CrossRef]
- Li, T.; Yang, X.; Yu, Y.; Si, X.; Zhai, X.; Zhang, H.; Dong, W.; Gao, C.; Xu, C. Domestication of wild tomato is accelerated by genome editing. Nat. Biotechnol. 2018. [Google Scholar] [CrossRef] [PubMed]
- Belhaj, K.; Chaparro-Garcia, A.; Kamoun, S.; Patron, N.J.; Nekrasov, V. Editing plant genomes with CRISPR/Cas9. Curr. Opin. Biotechnol. 2015, 32, 76–84. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Wang, M.; Zhang, S.; Ai, X. Changes in SBPase activity influence photosynthetic capacity, growth and tolerance to chilling stress in transgenic tomato plants. Sci. Rep. 2016, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Ding, F.; Wang, M.; Zhang, S. Overexpression of a Calvin cycle enzyme SBPase improves tolerance to chilling-induced oxidative stress in tomato plants. Sci. Hortic. 2017, 214, 27–33. [Google Scholar] [CrossRef]
- Harrison, E.P.; Olcer, H.; Lloyd, J.C.; Long, S.P.; Raines, C.A. Small decreases in SBPase cause a linear decline in the apparent RuBP regeneration rate but do not affect Rubisco carboxylation capacity. J. Exp. Bot. 2001, 52, 1779–1784. [Google Scholar] [CrossRef] [PubMed]
- Harrison, E.P.; Willingham, N.M.; Lloyd, J.C.; Raines, C.A. Reduced sedoheptulose-1,7-bisphosphatase levels in transgenic tobacco lead to decreased photosynthetic capacity and altered carbohydrate accumulation. Planta 1998, 204, 27–36. [Google Scholar] [CrossRef]
- Olçer, H.; Lloyd, J.C.; Raines, C.A. Photosynthetic capacity is differentially affected by reductions in sedoheptulose-1,7-bisphosphatase activity during leaf development in transgenic tobacco plants. Plant Physiol. 2001, 125, 982–989. [Google Scholar] [CrossRef]
- Lawson, T.; Bryant, B.; Lefebvre, S.; Lloyd, J.C.; Raines, C.A. Decreased SBPase activity alters growth and development in transgenic tobacco plants. Plant Cell Environ. 2006, 29, 48–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raines, C.A.; Harrison, E.P.; Lloyd, J.C. Investigating the role of the thiol-regulated enzyme sedoheptulose-1, 7-bisphosphatase in the control of photosynthesis. Physiol. Plant. 2000, 110, 303–308. [Google Scholar] [CrossRef]
- Simkin, A.J.; Lopez-Calcagno, P.E.; Davey, P.A.; Headland, L.R.; Lawson, T.; Timm, S.; Bauwe, H.; Raines, C.A. Simultaneous stimulation of sedoheptulose 1,7-bisphosphatase, fructose 1,6-bisphophate aldolase and the photorespiratory glycine decarboxylase-H protein increases CO2assimilation, vegetative biomass and seed yield in Arabidopsis. Plant Biotechnol. J. 2017, 15, 805–816. [Google Scholar] [CrossRef] [PubMed]
- Feng, L.; Han, Y.; Liu, G.; An, B.; Yang, J.; Yang, G.; Li, Y.; Zhu, Y. Overexpression of sedoheptulose-1, 7-bisphosphatase enhances photosynthesis and growth under salt stress in transgenic rice plants. Funct. Plant Biol. 2007, 34, 822–834. [Google Scholar] [CrossRef]
- Feng, L.; Wang, K.; Li, Y.; Tan, Y.; Kong, J.; Li, H.; Li, Y.; Zhu, Y. Overexpression of SBPase enhances photosynthesis against high temperature stress in transgenic rice plants. Plant Cell Rep. 2007, 26, 1635–1646. [Google Scholar] [CrossRef]
- Ding, F.; Wang, M.; Zhang, S. Sedoheptulose-1,7-bisphosphatase is involved in methyl jasmonate- and dark-induced leaf senescence in tomato plants. Int. J. Mol. Sci. 2018, 19, 3673. [Google Scholar] [CrossRef] [PubMed]
- Salvucci, M.E.; Ogren, W.L. The mechanism of Rubisco activase: Insights from studies of the properties and structure of the enzyme. Photosynth. Res. 1996, 47, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Raines, C.A.; Lloyd, J.C.; Willingham, N.M.; Potts, S.; Dyer, T.A. cDNA and gene sequences of wheat chloroplast sedoheptulose-1,7-bisphosphatase reveal homology with fructose-1,6-bisphosphatases. Eur. J. Biochem. 1992, 205, 1053–1059. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gütle, D.D.; Roret, T.; Müller, S.J.; Couturier, J.; Lemaire, S.D.; Hecker, A.; Dhalleine, T.; Buchanan, B.B.; Reski, R.; Einsle, O.; Jacquot, J.-P. Chloroplast FBPase and SBPase are thioredoxin-linked enzymes with similar architecture but different evolutionary histories. Proc. Natl. Acad. Sci. USA 2016, 113, 6779–6784. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloom, A.J. Photorespiration and nitrate assimilation: A major intersection between plant carbon and nitrogen. Photosynth. Res. 2015, 123, 117–128. [Google Scholar] [CrossRef]
- Miflin, B.J.; Habash, D.Z. The role of glutamine synthetase and glutamate dehydrogenase in nitrogen assimilation and possibilities. J. Exp. Bot. 2002, 53, 979–987. [Google Scholar] [CrossRef]
- Guo, Q.; Yoshida, Y.; Major, I.T.; Wang, K.; Sugimoto, K.; Kapali, G.; Havko, N.E.; Benning, C.; Howe, G.A. JAZ repressors of metabolic defense promote growth and reproductive fitness in Arabidopsis. Proc. Natl. Acad. Sci. USA 2018, 115, E10768–E10777. [Google Scholar] [CrossRef]
- Xie, X.; Ma, X.; Zhu, Q.; Zeng, D.; Li, G.; Liu, Y.G. CRISPR-GE: A convenient software toolkit for CRISPR-based genome editing. Mol. Plant 2017, 10, 1246–1249. [Google Scholar] [CrossRef]
- Dan, Y.; Yan, H.; Munyikwa, T.; Dong, J.; Zhang, Y.; Armstrong, C.L. MicroTom—A high-throughput model transformation system for functional genomics. Plant Cell Rep. 2006, 25, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Sicher, R.C.; Bahr, J.T.; Jensen, R.G. Measurement of ribulose 1, 5-bisphosphate from spinach chloroplasts. Plant Physiol. 1979, 64, 876–879. [Google Scholar] [CrossRef] [PubMed]
- Maeda, H.; Song, W.; Sage, T.L.; DellaPenna, D. Tocopherols play a crucial role in low-temperature adaptation and phloem loading in Arabidopsis. Plant Cell 2006, 18, 2710–2732. [Google Scholar] [CrossRef] [PubMed]
- Stitt, M.; Lilley, R.; Gerhardt, R.; Heldt, H.W. Metabolite levels in specific cells and subcellular compartments of plant leaves. In Methods in Enzymology; Fleischer, S., Fleischer, B., Eds.; Academic Press: Cambridge, MA, USA, 1989; Volume 174, pp. 518–552. [Google Scholar]
- Bradford, M.M. A rapid and sensitive method for the quantification of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
Figure 1.
Schematic diagram of SlSBPASE gene structure, vector structure and CRSIPR/Cas9-induced mutagenesis. (A) Schematic diagram of SlSBPASE gene structure and target sequence. ATG, start codon; TGA, stop codon; sgRNA, single guide RNA; PAM, protospacer adjacent motif; AGG in red, PAM sequence. (B) Schematic diagram of the vector used in this study. LB, left border of T-DNA (Transfer DNA); RB, right border of T-DNA; U6, Arobidopsis U6 promoter; gRNA, guide RNA; e35S, enhanced 35S promoter; Cas9, optimized Cas9; NOS (nopaline synthase) Ter, NOS terminator; 35S, CaMV (Cauliflower mosaic virus) 35S promoter; HPT (hygromycin phosphotransferase), hygromycin selection marker; PolyA Ter, PolyA terminator. (C) Representative genotypes of slsbpase mutants. WT, wild type. Target sequences are in blue.
Figure 1.
Schematic diagram of SlSBPASE gene structure, vector structure and CRSIPR/Cas9-induced mutagenesis. (A) Schematic diagram of SlSBPASE gene structure and target sequence. ATG, start codon; TGA, stop codon; sgRNA, single guide RNA; PAM, protospacer adjacent motif; AGG in red, PAM sequence. (B) Schematic diagram of the vector used in this study. LB, left border of T-DNA (Transfer DNA); RB, right border of T-DNA; U6, Arobidopsis U6 promoter; gRNA, guide RNA; e35S, enhanced 35S promoter; Cas9, optimized Cas9; NOS (nopaline synthase) Ter, NOS terminator; 35S, CaMV (Cauliflower mosaic virus) 35S promoter; HPT (hygromycin phosphotransferase), hygromycin selection marker; PolyA Ter, PolyA terminator. (C) Representative genotypes of slsbpase mutants. WT, wild type. Target sequences are in blue.
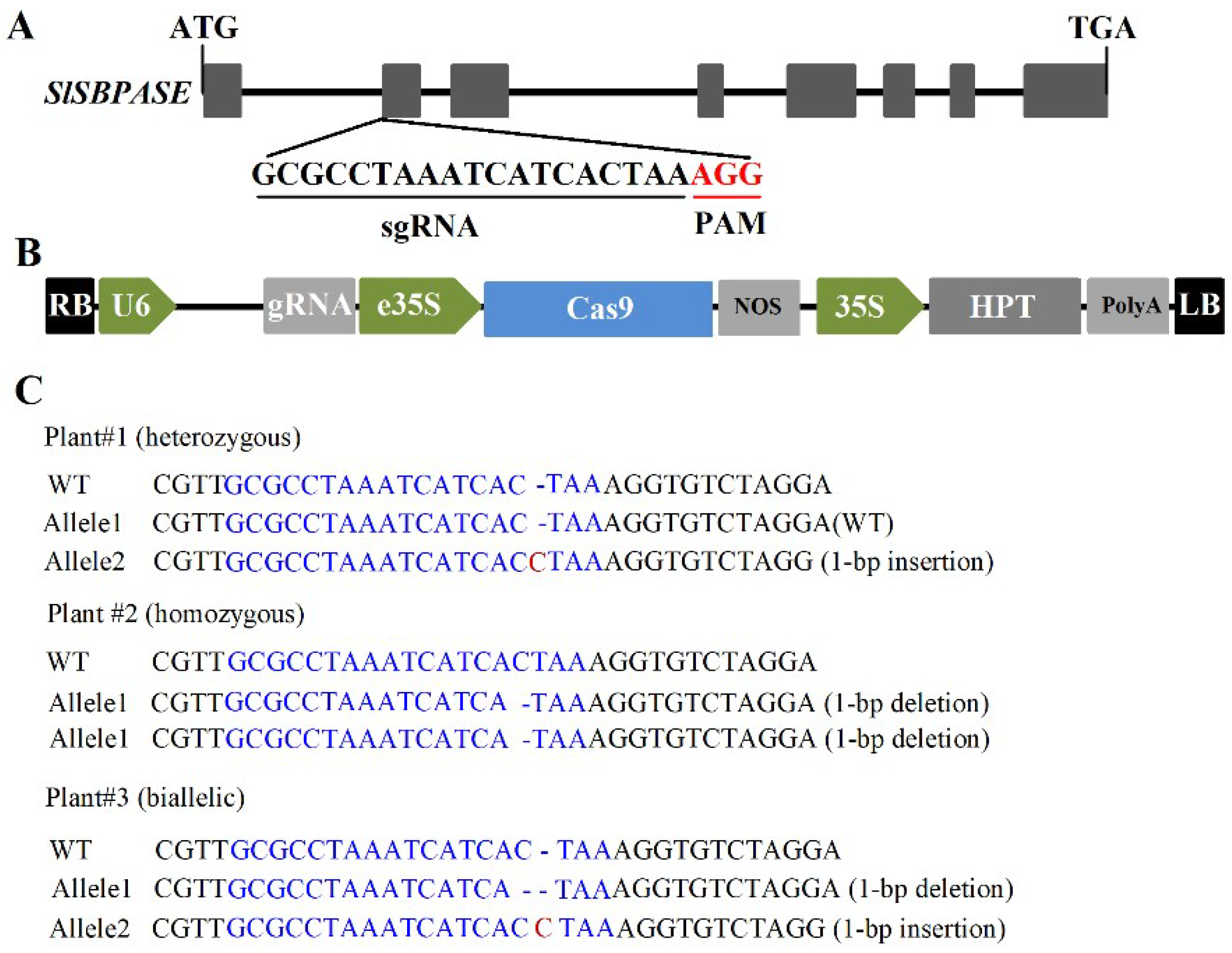
Figure 2.
Characterization of slsbpase mutant. (A) Phenotypes of slsbpase mutant plants (slsbpase) and wild-type plants (WT). (B) Plant height of wild-type and mutant plants. (C) Leaf chlorosis phenotype of slsbpase mutant. (D) Leaf chlorophyll content of wild-type and mutant plants. The values presented are means ± SDs (standard deviation, n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.
Figure 2.
Characterization of slsbpase mutant. (A) Phenotypes of slsbpase mutant plants (slsbpase) and wild-type plants (WT). (B) Plant height of wild-type and mutant plants. (C) Leaf chlorosis phenotype of slsbpase mutant. (D) Leaf chlorophyll content of wild-type and mutant plants. The values presented are means ± SDs (standard deviation, n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.

Figure 3.
Changes of SBPase activity (A), RuBP regeneration (B) and CO2 assimilation rates (C) in slsbpase mutant plants. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.
Figure 3.
Changes of SBPase activity (A), RuBP regeneration (B) and CO2 assimilation rates (C) in slsbpase mutant plants. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.

Figure 4.
Diurnal carbohydrate accumulation in wild-type and slsbpase mutant plants. Levels of sucrose (A,B) and starch (C,D) were determined in leaves of wild-type and slsbpase mutant plants in the light period (end of day) and at the end of the night. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.
Figure 4.
Diurnal carbohydrate accumulation in wild-type and slsbpase mutant plants. Levels of sucrose (A,B) and starch (C,D) were determined in leaves of wild-type and slsbpase mutant plants in the light period (end of day) and at the end of the night. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.
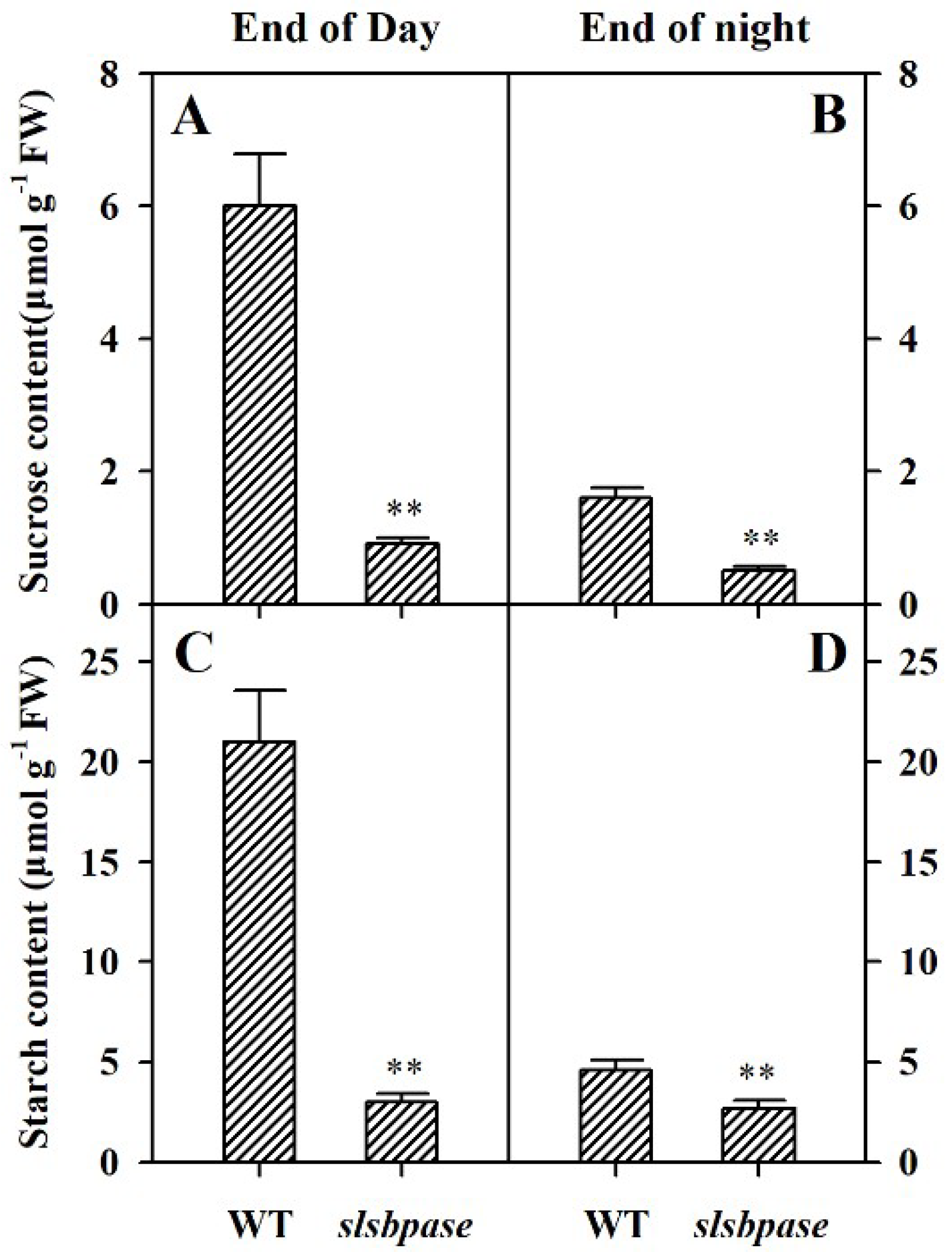
Figure 5.
Night respiration rates in wild-type tomato plants and slsbpase mutant plants. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.
Figure 5.
Night respiration rates in wild-type tomato plants and slsbpase mutant plants. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 between slsbpase mutant plants and wild-type plants.

Figure 6.
Changes in levels of total protein and amino acids as affected by mutation of SlSBPASE in tomato plants. (A) Protein level. (B) Amino acids level. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 and * p < 0.05 between slsbpase mutant plants and wild-type plants.
Figure 6.
Changes in levels of total protein and amino acids as affected by mutation of SlSBPASE in tomato plants. (A) Protein level. (B) Amino acids level. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 and * p < 0.05 between slsbpase mutant plants and wild-type plants.

Figure 7.
Activities of metabolic enzymes involved in nitrogen metabolism in wild-type plants and slsbpase mutant plants. (A) NR. (B) GS. (C) GOGAT. (D) GDH. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 and * p < 0.05 between slsbpase mutant plants and wild-type plants.
Figure 7.
Activities of metabolic enzymes involved in nitrogen metabolism in wild-type plants and slsbpase mutant plants. (A) NR. (B) GS. (C) GOGAT. (D) GDH. The values presented are means ± SDs (n = 3). Asterisks indicate significant difference at ** p < 0.01 and * p < 0.05 between slsbpase mutant plants and wild-type plants.

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Potential off-target sites and primers used in this study.
| Potential off-Target Sites | Primer Sequence (5′–3′) |
|---|---|
| Site 1 | F1: TGGGTCTGAATCATCACTAG |
| TGGGTCTGAATCATCACTAGAGG | R1: CTAGTGATGATTCAGACCCA |
| Site 2 | F2: AGTGCCTAAATCCTCAATAA |
| AGTGCCTAAATCCTCAATAAAGG | R2: TTATTGAGGATTTAGGCACT |
| Site 3 | F3: TGTGTCTAAATCATCATTCA |
| TGTGTCTAAATCATCATTCAAGG | R3: TGAATGATGATTTAGACACA |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ding, F.; Hu, Q.; Wang, M.; Zhang, S. Knockout of SlSBPASE Suppresses Carbon Assimilation and Alters Nitrogen Metabolism in Tomato Plants. Int. J. Mol. Sci. 2018, 19, 4046. https://doi.org/10.3390/ijms19124046
AMA Style
Ding F, Hu Q, Wang M, Zhang S. Knockout of SlSBPASE Suppresses Carbon Assimilation and Alters Nitrogen Metabolism in Tomato Plants. International Journal of Molecular Sciences. 2018; 19(12):4046. https://doi.org/10.3390/ijms19124046
Chicago/Turabian StyleDing, Fei, Qiannan Hu, Meiling Wang, and Shuoxin Zhang. 2018. "Knockout of SlSBPASE Suppresses Carbon Assimilation and Alters Nitrogen Metabolism in Tomato Plants" International Journal of Molecular Sciences 19, no. 12: 4046. https://doi.org/10.3390/ijms19124046
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.