Regulation of Three Virulence Strategies of Mycobacterium tuberculosis: A Success Story

 , ,
, ,
Abstract
:1. Introduction
2. Divalent Metals at the Interface of M. tuberculosis Host Interaction
2.1. Iron Homeostasis and Redox Sensing
2.2. Manganese Homeostasis and cAMP Production
2.3. Zinc Homeostasis
2.4. ESX-3 Secretion System
3. Three Main Virulence Strategies of Mtb
3.1. Immune Modulation
3.2. Phagosomal Rupture and Pore Formation
3.3. Dormancy and Modulation of Granuloma Formation
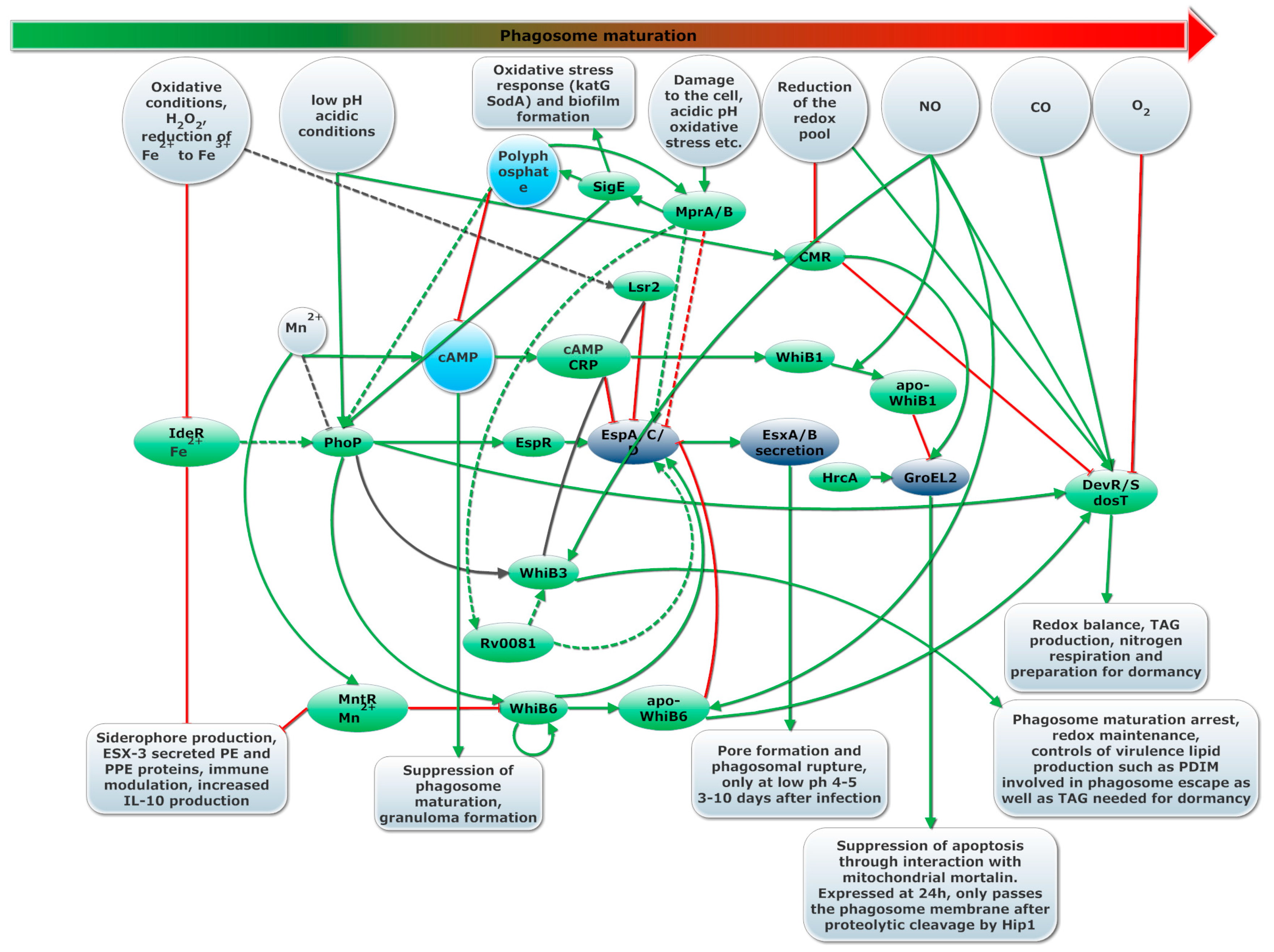
4. Success through Tight Regulation of Virulence Strategies
Supplementary Materials
Acknowledgments
Conflicts of Interest
Abbreviations
| AC | Adenylate cyclase |
| cAMP | Cyclic adenosine monophosphate |
| CMR | Cyclic-AMP and redox responsive transcription factor |
| CRP | Cyclic-AMP dependent regulatory protein |
| DAG | Diacylglycerol |
| DevRST | DevRST is a two component regulator and sensor, which regulate genes coding for proteins that help Mtb prepare for dormancy and subsequent resuscitation |
| EspR | A virulence associated transcriptional regulator upregulated by PhoP |
| IdeR | Iron-dependent regulator |
| Lsr2 | A histone like regulator that binds AT-rich regions virulence islands, acting as a global regulator to aid in the adaptation to extremes in oxygen availability |
| MntR | Manganese-dependent transcriptional repressor |
| MprAB | A two component sensor and regulator that responds to cell envelop stress |
| Mtb | Mycobacterium tuberculosis |
| PDIM | Phthiocerol dimycocerosates |
| SigE | Extracytoplasmic alternative Sigma factor E, involved in response to low pH and cell stress |
| Zur | Zinc uptake regulator |
References
- World Health Organization (WHO). Global Tuberculosis Report 2017; World Health Organization: Geneva, Switzerland, 2017. [Google Scholar]
- Meena, L.S. Rajni Survival mechanisms of pathogenic Mycobacterium tuberculosis H37Rv. FEBS J. 2010, 277, 2416–2427. [Google Scholar] [CrossRef] [PubMed]
- Gengenbacher, M.; Kaufmann, S.H.E. Mycobacterium tuberculosis: Success through dormancy. FEMS Microbiol. Rev. 2012, 36, 514–532. [Google Scholar] [CrossRef] [PubMed]
- Conrad, W.H.; Osman, M.M.; Shanahan, J.K.; Chu, F.; Takaki, K.K.; Cameron, J.; Hopkinson-Woolley, D.; Brosch, R.; Ramakrishnan, L. Mycobacterial ESX-1 secretion system mediates host cell lysis through bacterium contact-dependent gross membrane disruptions. Proc. Natl. Acad. Sci. USA 2017, 114, 1371–1376. [Google Scholar] [CrossRef] [PubMed]
- Dietzold, J.; Gopalakrishnan, A.; Salgame, P. Duality of lipid mediators in host response against Mycobacterium tuberculosis: Good cop, bad cop. F1000Prime Rep. 2015, 7, 1–8. [Google Scholar] [CrossRef] [PubMed]
- Guirado, E.; Schlesinger, L.S. Modeling the Mycobacterium tuberculosis granuloma—The critical battlefield in host immunity and disease. Front. Immunol. 2013, 4, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Silva Miranda, M.; Breiman, A.; Allain, S.; Deknuydt, F.; Altare, F. The tuberculous granuloma: An unsuccessful host defence mechanism providing a safety shelter for the bacteria? Clin. Dev. Immunol. 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Gomez, J.E.; McKinney, J.D.M. Tuberculosis persistence, latency, and drug tolerance. Tuberculosis 2004, 84, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Kapoor, N.; Pawar, S.; Sirakova, T.D.; Deb, C.; Warren, W.L.; Kolattukudy, P.E. Human granuloma in vitro model, for TB dormancy and resuscitation. PLoS ONE 2013, 8, e53657. [Google Scholar] [CrossRef] [PubMed]
- Paige, C.; Bishai, W.R. Penitentiary or penthouse condo: The tuberculous granuloma from the microbe’s point of view. Cell. Microbiol. 2010, 12, 301–309. [Google Scholar] [CrossRef] [PubMed]
- Shaler, C.R.; Horvath, C.N.; Jeyanathan, M.; Xing, Z. Within the Enemy’s Camp: Contribution of the granuloma to the dissemination, persistence and transmission of Mycobacterium tuberculosis. Front. Immunol. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Russell, D.; Cardona, P.; Kim, M. Foamy macrophages and the progression of the human tuberculosis granuloma. Nat. Immunol. 2009, 10, 943–948. [Google Scholar] [CrossRef] [PubMed]
- Simeone, R.; Bobard, A.; Lippmann, J.; Bitter, W.; Majlessi, L.; Brosch, R.; Enninga, J. Phagosomal rupture by Mycobacterium tuberculosis results in toxicity and host cell death. PLoS Pathog. 2012, 8, e1002507. [Google Scholar] [CrossRef] [PubMed]
- Sani, M.; Houben, E.N.G.; Geurtsen, J.; Pierson, J.; de Punder, K.; van Zon, M.; Wever, B.; Piersma, S.R.; Jiménez, C.R.; Daffé, M.; et al. Direct visualization by Cryo-EM of the mycobacterial capsular layer: A labile structure containing ESX-1-secreted proteins. PLoS Pathog. 2010, 6, e1000794. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lucarelli, D.; Vasil, M.L.; Meyer-Klaucke, W.; Pohl, E. The metal-dependent regulators FurA and FurB from Mycobacterium tuberculosis. Int. J. Mol. Sci. 2008, 9, 1548–1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Juttukonda, L.J.; Skaar, E.P. Manganese homeostasis and utilization in pathogenic bacteria. Mol. Microbiol. 2015, 97, 216–228. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.R.; Gros, P. Iron, manganese, and cobalt transport by Nramp1 (Slc11a1) and Nramp2 (Slc11a2) expressed at the plasma membrane. Blood 2003, 102, 1884–1892. [Google Scholar] [CrossRef] [PubMed]
- Indriate, M.; Skaar, E.P. Nutritional immunity: Transition metals at the pathogen-host interface. Nat. Rev. Microbiol. 2013, 10, 646–656. [Google Scholar] [CrossRef]
- Pandey, R.; Rodriguez, G.M. IdeR is required for iron homeostasis and virulence in Mycobacterium tuberculosis. Mol. Microbiol. 2014, 91, 98–109. [Google Scholar] [CrossRef] [PubMed]
- Papp-Wallace, K.M.; Maguire, M.E. Manganese transport and the role of manganese in virulence. Annu. Rev. Microbiol. 2006, 60, 187–209. [Google Scholar] [CrossRef] [PubMed]
- Reddy, S.K.; Kamireddi, M.; Dhanireddy, K.; Young, L.; Davis, A.; Reddy, P.T. Eukaryotic-like adenylyl cyclases in Mycobacterium tuberculosis H37Rv: Cloning and characterization. J. Biol. Chem. 2001, 276, 35141–35149. [Google Scholar] [CrossRef] [PubMed]
- Maciag, A.; Dainese, E.; Rodriguez, G.M.; Milano, A.; Provvedi, R.; Pasca, M.R.; Smith, I.; Palù, G.; Riccardi, G.; Manganelli, R. Global analysis of the Mycobacterium tuberculosis Zur (FurB) regulon. J. Bacteriol. 2007, 189, 730–740. [Google Scholar] [CrossRef] [PubMed]
- Kurthkoti, K.; Amin, H.; Marakalala, M.J.; Ghanny, S.; Subbian, S.; Sakatos, A.; Livny, J.; Fortune, S.M.; Berney, M.; Rodriguez, G.M. The capacity of Mycobacterium tuberculosis to survive iron starvation might enable it to persist in iron-deprived microenvironments of human granulomas. mBio 2017, 8, e01092-17. [Google Scholar] [CrossRef] [PubMed]
- Lin, H.; Andersen, G.R.; Yatime, L. Crystal structure of human S100A8 in complex with zinc and calcium. BMC Struct. Biol. 2016, 16, 8. [Google Scholar] [CrossRef] [PubMed]
- Olakanmi, O.; Schlesinger, L.S.; Ahmed, A.; Britigan, B.E. Intraphagosomal Mycobacterium tuberculosis acquires iron from both extracellular transferrin and intracellular iron pools. Impact of interferon-gamma and hemochromatosis. J. Biol. Chem. 2002, 277, 49727–49734. [Google Scholar] [CrossRef] [PubMed]
- Blanchette, C.D.; Woo, Y.-H.; Thomas, C.; Shen, N.; Sulchek, T.A.; Hiddessen, A.L. Decoupling internalization, acidification and phagosomal-endosomal/lysosomal fusion during phagocytosis of InlA coated beads in epithelial cells. PLoS ONE 2009, 4, e6056. [Google Scholar] [CrossRef] [PubMed]
- Forbes, J.R.; Gros, P. Divalent-metal transport by NRAMP proteins at the interface of host-pathogen interactions. Trends Microbiol. 2001, 9, 397–403. [Google Scholar] [CrossRef]
- Olakanmi, O.; Schlesinger, L.S.; Ahmed, A.; Britigan, B.E. The nature of extracellular iron influences iron acquisition by Mycobacterium tuberculosis residing within human macrophages. Infect. Immun. 2004, 72, 2022–2028. [Google Scholar] [CrossRef] [PubMed]
- Jabado, N.; Jankowski, A.; Dougaparsad, S.; Picard, V.; Grinstein, S.; Gros, P. Natural resistance to intracellular infections: Natural resistance-associated macrophage protein 1 (Nramp1) functions as a pH-dependent manganese transporter at the phagosomal membrane. J. Exp. Med. 2000, 192, 1237–1248. [Google Scholar] [CrossRef] [PubMed]
- Supek, F.; Supekova, L.; Nelson, H.; Nelson, N. A yeast manganese transporter related to the macrophage protein involved in conferring resistance to mycobacteria. Proc. Natl. Acad. Sci. USA 1996, 93, 5105–5110. [Google Scholar] [CrossRef] [PubMed]
- Peracino, B.; Buracco, S.; Bozzaro, S. The Nramp (Slc11) proteins regulate development, resistance to pathogenic bacteria and iron homeostasis in Dictyostelium discoideum. J. Cell Sci. 2013, 126, 301–311. [Google Scholar] [CrossRef] [PubMed]
- Wagner, D.; Maser, J.; Lai, B.; Cai, Z.; Barry, C.E., III; Höner zu Bentrup, K.; Russell, D.G.; Bermudez, L.; Iii, C.E.B. Elemental analysis of Mycobacterium avium-, Mycobacterium tuberculosis-, and Mycobacterium smegmatis-containing phagosomes indicates pathogen-induced microenvironments within the host cell’s endosomal system. J. Immunol. 2013, 174, 1491–1500. [Google Scholar]
- Pandey, R.; Russo, R.; Ghanny, S.; Huang, X.; Helmann, J.; Rodriguez, G.M. MntR(Rv2788): A transcriptional regulator that controls manganese homeostasis in Mycobacterium tuberculosis. Mol. Microbiol. 2015, 98, 1168–1183. [Google Scholar] [CrossRef] [PubMed]
- Pohl, E.; Holmes, R.K.; Hol, W.G. Crystal structure of the iron-dependent regulator (IdeR) from Mycobacterium tuberculosis shows both metal binding sites fully occupied. J. Mol. Biol. 1999, 285, 1145–1156. [Google Scholar] [CrossRef] [PubMed]
- Dewitt, M.A.; Kliegman, J.I.; Helmann, J.D.; Brennan, R.G.; David, L.; Glasfeld, A. The conformations of the manganese transport regulator of Bacillus subtilis in its metal-free state. J. Mol. Biol. 2007, 365, 1257–1265. [Google Scholar] [CrossRef] [PubMed]
- Luo, M.; Fadeev, E.A.; Groves, J.T. Mycobactin-mediated iron acquisition within macrophages. Nat. Chem. Biol. 2005, 1, 149–153. [Google Scholar] [CrossRef] [PubMed]
- McMahon, M.D.; Rush, J.S.; Thomas, M.G. Analyses of MbtB, MbtE, and MbtF suggest revisions to the Mycobactin biosynthesis pathway in Mycobacterium tuberculosis. J. Bacteriol. 2012, 194, 2809–2818. [Google Scholar] [CrossRef] [PubMed]
- Boradia, V.M.; Malhotra, H.; Thakkar, J.S.; Tillu, V.A.; Vuppala, B.; Patil, P.; Sheokand, N.; Sharma, P.; Chauhan, A.S.; Raje, M.; et al. Mycobacterium tuberculosis acquires iron by cell-surface sequestration and internalization of human holo-transferrin. Nat. Commun. 2014, 5, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madigan, C.A.; Cheng, T.-Y.; Layre, E.; Young, D.C.; McConnell, M.J.; Debono, C.A.; Murry, J.P.; Wei, J.-R.; Barry, C.E.; Rodriguez, G.M.; et al. Lipidomic discovery of deoxysiderophores reveals a revised mycobactin biosynthesis pathway in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2012, 109, 1257–1262. [Google Scholar] [CrossRef] [PubMed]
- Rohde, K.H.; Veiga, D.F.T.; Caldwell, S.; Balázsi, G.; Russell, D.G. Linking the transcriptional profiles and the physiological states of Mycobacterium tuberculosis during an extended intracellular infection. PLoS Pathog. 2012, 8. [Google Scholar] [CrossRef] [PubMed]
- Tyagi, P.; Dharmaraja, A.T.; Bhaskar, A.; Chakrapani, H.; Singh, A. Mycobacterium tuberculosis has diminished capacity to counteract redox stress induced by elevated levels of endogenous superoxide. Free Radic. Biol. Med. 2015, 84, 344–354. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, G.M.; Voskuil, M.I.; Gold, B.; Schoolnik, G.K.; Smith, I. ideR, an essential gene in Mycobacterium tuberculosis: Role of IdeR in iron-dependent gene expression, iron metabolism, and oxidative stress response. Infect. Immun. 2002, 70, 3371–3381. [Google Scholar] [CrossRef] [PubMed]
- Vilchèze, C.; Hartman, T.; Weinrick, B.; William, R.J., Jr. Mycobacterium tuberculosis is extraordinarily sensitive to killing by a vitamin C-induced Fenton reaction. Nat. Commun. 2013, 4. [Google Scholar] [CrossRef] [PubMed]
- Litwin, C.M.; Calderwood, S.B. Role of iron in regulation of virulence genes. Clin. Microbiol. Rev. 1993, 6, 137–149. [Google Scholar] [CrossRef] [PubMed]
- Schaible, U.E.; Kaufmann, S.H.E. Iron and microbial infection. Nat. Rev. Microbiol. 2004, 2, 946–953. [Google Scholar] [CrossRef] [PubMed]
- Outten, F.W.; Theil, E.C. Iron-based redox switches in biology. Antioxid. Redox Signal. 2009, 11, 1029–1046. [Google Scholar] [CrossRef] [PubMed]
- Siegrist, M.S.; Unnikrishnan, M.; Mcconnell, M.J.; Borowsky, M.; Cheng, T.; Siddiqi, N.; Fortune, S.M.; Moody, D.B.; Rubin, E.J. Mycobacterial Esx-3 is required for mycobactin-mediated iron acquisition. Proc. Natl. Acad. Sci. USA 2009, 106, 18792–18797. [Google Scholar] [CrossRef] [PubMed]
- Serafini, A.; Boldrin, F.; Palù, G.; Manganelli, R.; Palu, G. Characterization of a Mycobacterium tuberculosis ESX-3 conditional mutant: Essentiality and rescue by iron and zinc. J. Bacteriol. 2009, 191, 6340–6344. [Google Scholar] [CrossRef] [PubMed]
- Farhana, A.; Kumar, S.; Rathore, S.S.; Ghosh, P.C.; Ehtesham, N.Z.; Tyagi, A.K.; Hasnain, S.E. Mechanistic insights into a novel exporter-importer system of Mycobacterium tuberculosis unravel its role in trafficking of iron. PLoS ONE 2008, 3, e2087. [Google Scholar] [CrossRef] [PubMed]
- Gold, B.; Rodriguez, G.M.; Marras, S.A.; Pentecost, M.; Smith, I. The Mycobacterium tuberculosis IdeR is a dual functional regulator that controls transcription of genes involved in iron acquisition, iron storage and survival in macrophages. Mol. Microbiol. 2001, 42, 851–865. [Google Scholar] [CrossRef] [PubMed]
- Rodriguez, G.M.; Smith, I. Mechanisms of iron regulation in mycobacteria: Role in physiology and virulence. Mol. Microbiol. 2003, 47, 1485–1494. [Google Scholar] [CrossRef] [PubMed]
- Oldridge, D.A.; Wood, A.C.; Weichert-leahey, N.; Crimmins, I.; Winter, C.; Mcdaniel, L.D.; Diamond, M.; Hart, L.S.; Durbin, A.D.; Abraham, B.J.; et al. The mycobacterial iron dependent regulator IdeR induces ferritin (bfrB) by alleviating Lsr2 repression. Mol. Microbiol. 2016, 528, 418–421. [Google Scholar] [CrossRef]
- Fu, G.; Lees, R.S.; Aw, D.; Jin, L.; Gray, P.; Berendonk, T.U.; White-cooper, H.; Scaife, S.; Phuc, H.K.; Jasinskiene, N.; et al. Lsr2 is a nucleoid-associated protein that targets AT-rich sequences and virulence genes in Mycobacterium tuberculosis. Proc. Natl. Acad. Sci. USA 2010, 107, 18741. [Google Scholar] [CrossRef]
- Colangeli, R.; Haq, A.; Arcus, V.L.; Summers, E.; Magliozzo, R.S.; McBride, A.; Mitra, A.K.; Radjainia, M.; Khajo, A.; Jacobs, W.R.; et al. The multifunctional histone-like protein Lsr2 protects mycobacteria against reactive oxygen intermediates. Proc. Natl. Acad. Sci. USA 2009, 106, 4414–4418. [Google Scholar] [CrossRef] [PubMed]
- Colangeli, R.; Helb, D.; Vilchèze, C.; Hazbón, M.H.; Lee, C.G.; Safi, H.; Sayers, B.; Sardone, I.; Jones, M.B.; Fleischmann, R.D.; et al. Transcriptional regulation of multi-drug tolerance and antibiotic-induced responses by the histone-like protein Lsr2 in M. tuberculosis. PLoS Pathog. 2007, 3, 780–793. [Google Scholar] [CrossRef] [PubMed]
- Khare, G.; Nangpal, P.; Tyagi, A.K. Differential roles of iron storage proteins in maintaining the iron homeostasis in Mycobacterium tuberculosis. PLoS ONE 2017, 12, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Jones, C.M.; Wells, R.M.; Madduri, A.V.R.; Renfrow, M.B.; Ratledge, C.; Moody, D.B.; Niederweis, M. Self-poisoning of Mycobacterium tuberculosis by interrupting siderophore recycling. Proc. Natl. Acad. Sci. USA 2014, 111, 1945–1950. [Google Scholar] [CrossRef] [PubMed]
- Pandey, S.D.; Choudhury, M.; Yousuf, S.; Wheeler, P.R.; Gordon, S.V.; Ranjan, A.; Sritharan, M. Iron-regulated protein HupB of Mycobacterium tuberculosis positively regulates siderophore biosynthesis and is essential for growth in macrophages. J. Bacteriol. 2014, 196, 1853–1865. [Google Scholar] [CrossRef] [PubMed]
- Sritharan, M. Iron homeostasis in Mycobacterium tuberculosis: Mechanistic insights into siderophore-mediated iron uptake. Bacteriology 2016, 198, 2399–2409. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Asensio, J.; Mostowy, S.; Harders-Westerveen, J.; Huygen, K.; Hernández-Pando, R.; Thole, J.; Behr, M.; Gicquel, B.; Martín, C. PhoP: A missing piece in the intricate puzzle of Mycobacterium tuberculosis virulence. PLoS ONE 2008, 3, e3496. [Google Scholar] [CrossRef] [PubMed]
- Gupta, S.; Sinha, A.; Sarkar, D. Transcriptional autoregulation by Mycobacterium tuberculosis PhoP involves recognition of novel direct repeat sequences in the regulatory region of the promoter. FEBS Lett. 2006, 580, 5328–5338. [Google Scholar] [CrossRef] [PubMed]
- Gonzalo-Asensio, J.; Soto, C.Y.; Arbués, A.; Sancho, J.; del Carmen Menéndez, M.; García, M.J.; Gicquel, B.; Martín, C. The Mycobacterium tuberculosis phoPR operon is positively autoregulated in the virulent strain H37Rv. J. Bacteriol. 2008, 190, 7068–7078. [Google Scholar] [CrossRef] [PubMed]
- Manabe, Y.C.; Saviola, B.J.; Sun, L.; Murphy, J.R.; Bishai, W.R. Attenuation of virulence in Mycobacterium tuberculosis expressing a constitutively active iron repressor. Proc. Natl. Acad. Sci. USA 1999, 96, 12844–12848. [Google Scholar] [CrossRef] [PubMed]
- Banerjee, S.; Nandyala, A.K.; Raviprasad, P.; Ahmed, N.; Hasnain, S.E. Iron-dependent RNA-binding activity of Mycobacterium tuberculosis aconitase. J. Bacteriol. 2007, 189, 4046–4052. [Google Scholar] [CrossRef] [PubMed]
- Pechter, K.B.; Meyer, F.M.; Serio, A.W.; Stülke, J.; Sonenshein, A.L. Two roles for aconitase in the regulation of tricarboxylic acid branch gene expression in Bacillus subtilis. J. Bacteriol. 2013, 195, 1525–1537. [Google Scholar] [CrossRef] [PubMed]
- Rohde, K.H.; Abramovitch, R.B.; Russell, D.G. Mycobacterium tuberculosis invasion of macrophages: Linking bacterial gene expression to environmental cues. Cell Host Microbe 2007, 2, 352–364. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Wang, Z.; Feng, L.; Chen, Z.; Mao, C.; Zhu, Y.; Chen, S. σE-dependent activation of RbpA controls transcription of the furA-katG operon in response to oxidative stress in mycobacteria. Mol. Microbiol. 2016, 102, 107–120. [Google Scholar] [CrossRef] [PubMed]
- Zheng, F.; Long, Q.; Xie, J. The function and regulatory network of WhiB and WhiB-like protein from comparative genomics and systems biology perspectives. Cell Biochem. Biophys. 2012, 63, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Reeves, A.Z.; Campbell, P.J.; Sultana, R.; Malik, S.; Murray, M.; Plikaytis, B.B.; Shinnick, T.M.; Posey, J.E. Aminoglycoside cross-resistance in Mycobacterium tuberculosis due to mutations in the 5′ untranslated region of whiB7. Antimicrob. Agents Chemother. 2013, 57, 1857–1865. [Google Scholar] [CrossRef] [PubMed]
- Samuel, L.P.; Song, C.-H.; Wei, J.; Roberts, E.A.; Dahl, J.L.; Barry, C.E.; Jo, E.-K.; Friedman, R.L. Expression, production and release of the Eis protein by Mycobacterium tuberculosis during infection of macrophages and its effect on cytokine secretion. Microbiology 2007, 153, 529–540. [Google Scholar] [CrossRef] [PubMed]
- Farina, M.; Avila, D.S.; Da Rocha, J.B.T.; Aschner, M. Metals, oxidative stress and neurodegeneration: A focus on iron, manganese and mercury. Neurochem. Int. 2013, 62, 575–594. [Google Scholar] [CrossRef] [PubMed]
- Champion, O.L.; Karlyshev, A.; Cooper, I.A.M.; Ford, D.C.; Wren, B.W.; Duffield, M.; Oyston, P.C.F.; Titball, R.W. Yersinia pseudotuberculosis mntH functions in intracellular manganese accumulation, which is essential for virulence and survival in cells expressing functional Nramp1. Microbiology 2011, 157, 1115–1122. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Farhana, A.; Guidry, L.; Saini, V.; Hondalus, M.; Steyn, A.J.C. Redox homeostasis in mycobacteria: The key to tuberculosis control? Expert Rev. Mol. Med. 2011, 13, e39. [Google Scholar] [CrossRef] [PubMed]
- Akhter, Y.; Yellaboina, S.; Farhana, A.; Ranjan, A.; Ahmed, N.; Hasnain, S.E. Genome scale portrait of cAMP-receptor protein (CRP) regulons in mycobacteria points to their role in pathogenesis. Gene 2008, 407, 148–158. [Google Scholar] [CrossRef] [PubMed]
- Matange, N. Revisiting bacterial cyclic nucleotide phosphodiesterases: Cyclic AMP hydrolysis and beyond. FEMS Microbiol. Lett. 2015, 362, fnv183. [Google Scholar] [CrossRef] [PubMed]
- Dass, B.K.M.; Sharma, R.; Shenoy, A.R.; Mattoo, R.; Visweswariah, S.S. Cyclic AMP in mycobacteria: Characterization and functional role of the Rv1647 ortholog in Mycobacterium smegmatis. J. Bacteriol. 2008, 190, 3824–3834. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Knapp, G.S.; McDonough, K.A. Cyclic AMP signalling in mycobacteria: Redirecting the conversation with a common currency. Cell. Microbiol. 2011, 13, 349–358. [Google Scholar] [CrossRef] [PubMed]
- Dittrich, D.; Keller, C.; Ehlers, S.; Schultz, J.E.; Sander, P. Characterization of a Mycobacterium tuberculosis mutant deficient in pH-sensing adenylate cyclase Rv1264. Int. J. Med. Microbiol. 2006, 296, 563–566. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Lamichhane, G.; Gupta, R.; Nolan, S.; Bishai, W.R. Cyclic AMP intoxication of macrophages by a Mycobacterium tuberculosis adenylate cyclase. Nature 2009, 460, 98–102. [Google Scholar] [CrossRef] [PubMed]
- Daniel, J.; Abraham, L.; Martin, A.; Pablo, X.; Reyes, S. Rv2477c is an antibiotic-sensitive manganese-dependent ABC-F ATPase in Mycobacterium tuberculosis. Biochem. Biophys. Res. Commun. 2017, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Rishi, P.; Jindal, N.; Bharrhan, S.; Tiwari, R.P. Salmonella-macrophage interactions upon manganese supplementation. Biol. Trace Elem. Res. 2010, 133, 110–119. [Google Scholar] [CrossRef] [PubMed]
- Diaz-ochoa, V.E.; Lam, D.; Lee, C.S.; Chazin, W.J.; Skaar, E.P.; Raffatellu, M.; Behnsen, J.; Liu, J.Z.; Chim, N. Salmonella mitigates oxidative stress and thrives in the inflamed gut by evading calprotectin-mediated article salmonella mitigates oxidative stress and thrives in the inflamed gut by evading calprotectin-mediated manganese sequestration. Cell Host Microbe 2016, 19, 814–825. [Google Scholar] [CrossRef] [PubMed]
- Agranoff, D.; Monahan, I.M.; Mangan, J.A.; Butcher, P.D.; Krishna, S. Mycobacterium tuberculosis expresses a novel pH-dependent divalent cation transporter belonging to the Nramp family. J. Exp. Med. 1999, 190, 717–724. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Samten, B.; Cao, G.; Wang, X.; Tvinnereim, A.R.; Chen, X.-L.; Howard, S.T. MprAB regulates the espA operon in Mycobacterium tuberculosis and modulates ESX-1 function and host cytokine response. J. Bacteriol. 2013, 195, 66–75. [Google Scholar] [CrossRef] [PubMed]
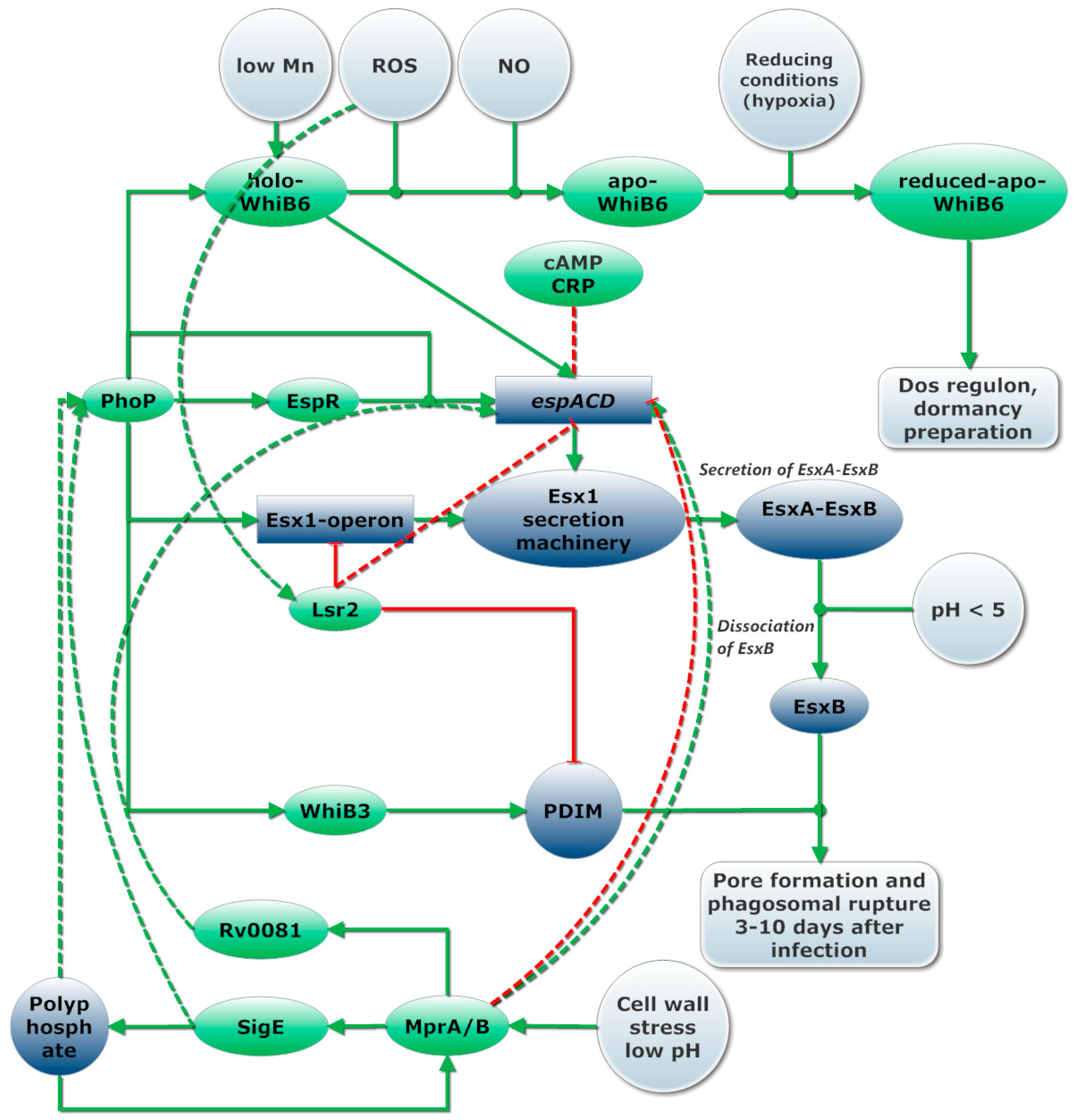
- Chen, Z.; Hu, Y.; Cumming, B.M.; Lu, P.; Feng, L.; Deng, J.; Steyn, A.J.C.; Chen, S. Mycobacterial WhiB6 differentially regulates ESX-1 and the dos regulon to modulate granuloma formation and virulence in zebrafish. Cell Rep. 2016, 16, 2512–2524. [Google Scholar] [CrossRef] [PubMed]
- Korch, S.B.; Contreras, H.; Clark-curtiss, J.E. Three Mycobacterium tuberculosis Rel Toxin-antitoxin modules inhibit mycobacterial growth and are expressed in infected human macrophages. J. Bacteriol. 2009, 191, 1618–1630. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Gao, C.H.; Hu, J.; Dong, C.; He, Z.G. Characterization of the interaction between a SirR family transcriptional factor of Mycobacterium tuberculosis, encoded by Rv2788, and a pair of toxin-antitoxin proteins RelJ/K, encoded by Rv3357 and Rv3358. FEBS J. 2014, 281, 2726–2737. [Google Scholar] [CrossRef] [PubMed]
- Kang, S.-M.; Kim, D.-H.; Lee, K.-Y.; Park, S.J.; Yoon, H.-J.; Lee, S.J.; Im, H.; Lee, B.-J. Functional details of the Mycobacterium tuberculosis VapBC26 toxin-antitoxin system based on a structural study: Insights into unique binding and antibiotic peptides. Nucleic Acids Res. 2017, 45, 8564–8580. [Google Scholar] [CrossRef] [PubMed]
- Lee, I.; Lee, S.J.; Chae, S.; Lee, K.; Kim, J.; Lee, B. Structural and functional studies of the Mycobacterium tuberculosis VapBC30 toxin-antitoxin system: Implications for the design of novel antimicrobial peptides. Nucleic Acids Res. 2015, 43, 7624–7637. [Google Scholar] [CrossRef] [PubMed]
- Serafini, A.; Pisu, D.; Palù, G.; Rodriguez, G.M.; Manganelli, R. The ESX-3 secretion system is necessary for iron and zinc homeostasis in Mycobacterium tuberculosis. PLoS ONE 2013, 8, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bretl, D.J.; Bigley, T.M.; Terhune, S.S.; Zahrt, T.C. The MprB extracytoplasmic domain negatively regulates activation of the Mycobacterium tuberculosis MprAB two-component system. J. Bacteriol. 2014, 196, 391–406. [Google Scholar] [CrossRef] [PubMed]
- Ilghari, D.; Lightbody, K.L.; Veverka, V.; Waters, L.C.; Muskett, F.W.; Renshaw, P.S.; Carr, M.D.; Muskett, W. Solution structure of the Mycobacterium tuberculosis EsxG·EsxH complex: Functional implications and comparisons with other M. tuberculosis Esx family complexes. J. Biol. Chem. 2011, 286, 29993–30002. [Google Scholar] [CrossRef] [PubMed]
- Gabriel, S.E.; Helmann, J.D. Contributions of zur-controlled ribosomal proteins to growth under zinc starvation conditions. J. Bacteriol. 2009, 191, 6116–6122. [Google Scholar] [CrossRef] [PubMed]
- Houben, E.N.G.; Korotkov, K.V.; Bitter, W. Take five—Type VII secretion systems of mycobacteria. Biochim. Biophys. Acta 2013. [Google Scholar] [CrossRef] [PubMed]
- Newton-Foot, M. The Mycobacterium tuberculosis ESX-3 Secretion System Interactome. Master’s Thesis, University of Stellenbsoch, Stellenbsoch, South Africa, 2010. [Google Scholar]
- Simeone, R.; Bottai, D.; Brosch, R. ESX/type VII secretion systems and their role in host-pathogen interaction. Curr. Opin. Microbiol. 2009, 12, 4–10. [Google Scholar] [CrossRef] [PubMed]
- Mehra, A.; Zahra, A.; Thompson, V.; Sirisaengtaksin, N.; Wells, A.; Porto, M.; Köster, S.; Penberthy, K.; Kubota, Y.; Dricot, A.; et al. Mycobacterium tuberculosis Type VII Secreted Effector EsxH Targets Host ESCRT to Impair Trafficking. PLoS Pathog. 2013, 9. [Google Scholar] [CrossRef] [PubMed]
- Tufariello, J.M.; Chapman, J.R.; Kerantzas, C.A.; Wong, K.-W.; Vilchèze, C.; Jones, C.M.; Cole, L.E.; Tinaztepe, E.; Thompson, V.; Fenyö, D.; et al. Separable roles for Mycobacterium tuberculosis ESX-3 effectors in iron acquisition and virulence. Proc. Natl. Acad. Sci. USA 2016, 113, E348–E357. [Google Scholar] [CrossRef] [PubMed]
- Van Dam, J.C.J.; Schaap, P.J.; Martins dos Santos, V.A.P.; Suárez-diez, M. Integration of heterogeneous molecular networks to unravel gene-regulation in Mycobacterium tuberculosis. BMC Med. Genom. 2014, 8, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Tiwari, B.M.; Kannan, N.; Vemu, L.; Raghunand, T.R. The Mycobacterium tuberculosis PE proteins Rv0285 and Rv1386 modulate innate immunity and mediate bacillary survival in macrophages. PLoS ONE 2012, 7, e51686. [Google Scholar] [CrossRef] [PubMed]
- Li, W.; Zhao, Q.; Deng, W.; Chen, T.; Liu, M.; Xie, J. Mycobacterium tuberculosis Rv3402c Enhances mycobacterial survival within macrophages and modulates the host pro-inflammatory cytokines production via NF-Kappa B/ERK/p38 signaling. PLoS ONE 2014, 9, e94418. [Google Scholar] [CrossRef] [PubMed]
- Daim, S.; Kawamura, I.; Tsuchiya, K.; Hara, H.; Kurenuma, T.; Shen, Y.; Dewamitta, S.R.; Sakai, S.; Nomura, T.; Qu, H.; et al. Expression of the Mycobacterium tuberculosis PPE37 protein in Mycobacterium smegmatis induces low tumour necrosis factor alpha and interleukin 6 production in murine macrophages. J. Med. Microbiol. 2011, 60, 582–591. [Google Scholar] [CrossRef] [PubMed]
- Yeruva, V.C.; Kulkarni, A.; Khandelwal, R.; Sharma, Y.; Raghunand, T.R. The PE_PGRS proteins of Mycobacterium tuberculosis are Ca2+ binding mediators of host–pathogen interaction. Biochemistry 2016, 55, 4675–4687. [Google Scholar] [CrossRef] [PubMed]
- Meng, L.; Tong, J.; Wang, H.; Tao, C.; Wang, Q.; Niu, C.; Zhang, X.; Gao, Q. PPE38 protein of Mycobacterium tuberculosis inhibits macrophage MHC class I expression and dampens CD8+ T cell responses. Front. Cell. Infect. Microbiol. 2017, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Dong, D.; Wang, D.; Li, M.; Wang, H.; Yu, J.; Wang, C.; Liu, J.; Gao, Q. PPE38 modulates the innate immune response and is required for Mycobacterium marinum virulence. Infect. Immun. 2012, 80, 43–54. [Google Scholar] [CrossRef] [PubMed]
- Cyktor, J.C.; Carruthers, B.; Kominsky, R.A.; Beamer, G.L.; Stromberg, P.; Turner, J. IL-10 inhibits mature fibrotic granuloma formation during Mycobacterium tuberculosis infection. J. Immunol. 2013, 190, 2778–2790. [Google Scholar] [CrossRef] [PubMed]
- Mahajan, S.; Dkhar, H.K.; Chandra, V.; Dave, S.; Nanduri, R.; Janmeja, A.K.; Agrewala, J.N.; Gupta, P. Mycobacterium tuberculosis modulates macrophage lipid-sensing nuclear receptors PPARγ and TR4 for survival. J. Immunol. 2012, 188, 5593–5603. [Google Scholar] [CrossRef] [PubMed]
- Han, X.; Kitamoto, S.; Wang, H.; Boisvert, W.A. Interleukin-10 overexpression in macrophages suppresses atherosclerosis in hyperlipidemic mice. FASEB J. 2010, 24, 2869–2880. [Google Scholar] [CrossRef] [PubMed]
- Prados-Rosales, R.; Weinrick, B.C.; Piqué, D.G.; Jacobs, W.R.; Casadevall, A.; Rodriguez, G.M. Role for Mycobacterium tuberculosis membrane vesicles in iron acquisition. J. Bacteriol. 2014, 196, 1250–1256. [Google Scholar] [CrossRef] [PubMed]
- Seto, S.; Tsujimura, K.; Koide, Y. Rab GTPases regulating phagosome maturation are differentially recruited to mycobacterial phagosomes. Traffic 2011, 12, 407–420. [Google Scholar] [CrossRef] [PubMed]
- Via, L.E.; Dusanka, D.; Roseann, J.U.; Nina, S.H.; Huber, L.A.; Deretic, V. Arrest of mycobacterial phagosome maturation is caused by a block in vesicle fusion between stages controlled by rab5 and rab7. J. Biol. Chem. 1997, 272, 13326–13331. [Google Scholar] [CrossRef] [PubMed]
- Wong, D.; Bach, H.; Sun, J.; Hmama, Z.; Av-Gay, Y. Mycobacterium tuberculosis protein tyrosine phosphatase (PtpA) excludes host vacuolar-H+-ATPase to inhibit phagosome acidification. Proc. Natl. Acad. Sci. USA 2011, 108, 19371–19376. [Google Scholar] [CrossRef] [PubMed]
- Kalamidas, S.A.; Kuehnel, M.P.; Peyron, P.; Rybin, V.; Rauch, S.; Kotoulas, O.B.; Houslay, M.; Hemmings, B.A.; Gutierrez, M.G.; Anes, E.; et al. cAMP synthesis and degradation by phagosomes regulate actin assembly and fusion events: Consequences for mycobacteria. J. Cell Sci. 2006, 119, 3686–3694. [Google Scholar] [CrossRef] [PubMed]
- Gupta, A.; Kaul, A.; Tsolaki, A.G.; Kishore, U.; Bhakta, S. Mycobacterium tuberculosis: Immune evasion, latency and reactivation. Immunobiology 2012, 217, 363–374. [Google Scholar] [CrossRef] [PubMed]
- Thi, E.P.; Lambertz, U.; Reiner, N.E. Sleeping with the enemy: How intracellular pathogens cope with a macrophage lifestyle. PLoS Pathog. 2012, 8, e1002551. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, L.; Pieters, J. Mycobacterial subversion of chemotherapeutic reagents and host defense tactics: Challenges in tuberculosis drug development. Annu. Rev. Pharmacol. Toxicol. 2009, 49, 427–453. [Google Scholar] [CrossRef] [PubMed]
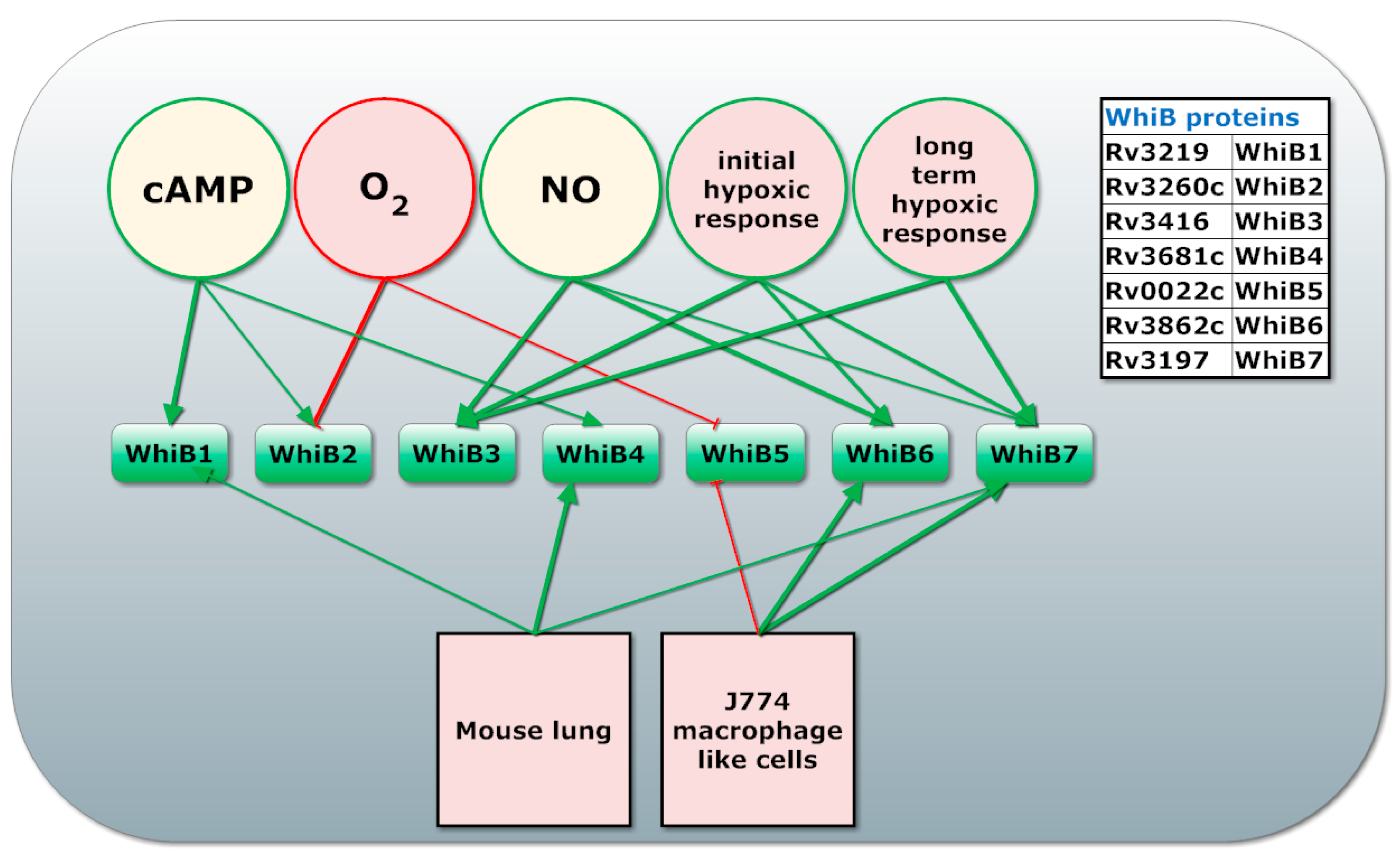
- Alam, M.S.; Garg, S.K.; Agrawal, P. Studies on structural and functional divergence among seven WhiB proteins of Mycobacterium tuberculosis H37Rv. FEBS J. 2009, 276, 76–93. [Google Scholar] [CrossRef] [PubMed]
- Larsson, C.; Luna, B.; Ammerman, N.C.; Maiga, M.; Agarwal, N.; Bishai, W.R. Gene expression of Mycobacterium tuberculosis putative transcription factors whiB1-7 in redox environments. PLoS ONE 2012, 7, e37516. [Google Scholar] [CrossRef] [PubMed]
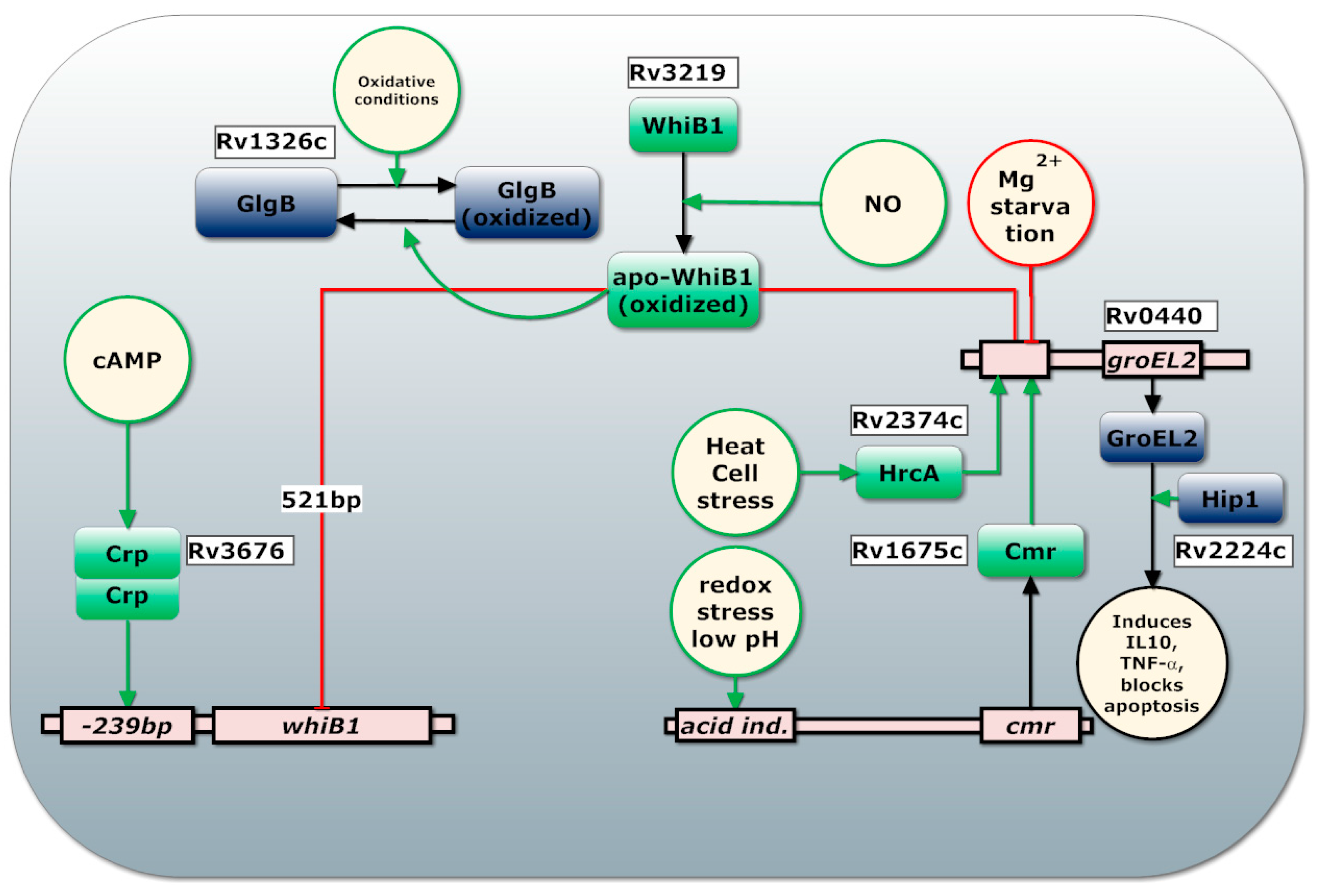
- Smith, L.J.; Stapleton, M.R.; Fullstone, G.J.M.; Crack, J.C.; Thomson, J.; Le Brun, N.E.; Hunt, D.M.; Harvey, E.; Adinolfi, S.; Buxton, R.S.; et al. Europe PMC Funders Group Mycobacterium tuberculosis WhiB1 is an essential DNA-binding protein with a nitric oxide sensitive iron-sulphur cluster. Biochem. J. 2010, 432, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Ranganathan, S.; Bai, G.; Lyubetskaya, A.; Gwendowlyn, S.; Galagan, E.; Mcdonough, K.A. Characterization of a cAMP responsive transcription factor, Cmr (Rv1675c), in TB complex mycobacteria reveals overlap with the DosR (DevR) dormancy regulon. Nucleic Acids Res. 2002, 1–54. [Google Scholar] [CrossRef] [PubMed]
- Chawla, M.; Parikh, P.; Saxena, A.; Munshi, M.; Mehta, M.; Mai, D.; Srivastava, A.K.; Narasimhulu, K.V.; Redding, K.E.; Vashi, N.; et al. Mycobacterium tuberculosis WhiB4 regulates oxidative stress response to modulate survival and dissemination in vivo. Mol. Microbiol. 2012, 85, 1148–1165. [Google Scholar] [CrossRef] [PubMed]
- Casonato, S.; Cervantes Sánchez, A.; Haruki, H.; Rengifo González, M.; Provvedi, R.; Dainese, E.; Jaouen, T.; Gola, S.; Bini, E.; Vicente, M.; et al. WhiB5, a transcriptional regulator that contributes to Mycobacterium tuberculosis virulence and reactivation. Infect. Immun. 2012, 80, 3132–3144. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Guidry, L.; Narasimhulu, K.V.; Mai, D.; Trombley, J.; Redding, K.E.; Giles, G.I.; Lancaster, J.R.; Steyn, A.J.C. Mycobacterium tuberculosis WhiB3 responds to O2 and nitric oxide via its [4Fe-4S] cluster and is essential for nutrient starvation survival. Proc. Natl. Acad. Sci. USA 2007, 104, 11562–11567. [Google Scholar] [CrossRef] [PubMed]
- Stapleton, M.R.; Smith, L.J.; Hunt, D.M.; Buxton, R.S.; Green, J. Mycobacterium tuberculosis WhiB1 represses transcription of the essential chaperonin GroEL2. Tuberculosis 2012, 92, 328–332. [Google Scholar] [CrossRef] [PubMed]
- Abramovitch, R.B.; Rohde, K.H.; Hsu, F.-F.; Russell, D.G. aprABC: A Mycobacterium tuberculosis complex-specific locus that modulates pH-driven adaptation to the macrophage phagosome. Mol. Microbiol. 2011, 80, 678–694. [Google Scholar] [CrossRef] [PubMed]
- Singh, A.; Crossman, D.K.; Mai, D.; Guidry, L.; Voskuil, M.I.; Renfrow, M.B.; Steyn, A.J.C. Mycobacterium tuberculosis WhiB3 maintains redox homeostasis by regulating virulence lipid anabolism to modulate macrophage response. PLoS Pathog. 2009, 5, e1000545. [Google Scholar] [CrossRef] [PubMed]
- Rienksma, R.A.; Suárez-Diez, M.; Mollenkopf, H.-J.; Dolganov, G.M.; Dorhoi, A.; Schoolnik, G.K.; Martins dos Santos, V.A.P.; Kaufmann, S.H.E.; Schaap, P.J.; Gengenbacher, M. Comprehensive insights into transcriptional adaptation of intracellular mycobacteria by microbe-enriched dual RNA sequencing. BMC Genom. 2015, 16, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, M.; Kogadeeva, M.; Gengenbacher, M.; McEwen, G.; Mollenkopf, H.; Zamboni, N.; Kaufmann, S.H.E.; Sauer, U. Integration of metabolomics and transcriptomics reveals a complex diet of mycobacterium tuberculosis during early macrophage infection. MSystems 2017, 2, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Galagan, J.E.; Minch, K.; Peterson, M.; Lyubetskaya, A.; Azizi, E.; Sweet, L.; Gomes, A.; Rustad, T.; Dolganov, G.; Glotova, I.; et al. The Mycobacterium tuberculosis regulatory network and hypoxia. Nature 2013, 499, 178–183. [Google Scholar] [CrossRef] [PubMed]
- Saini, V.; Farhana, A.; Steyn, A.J.C. Mycobacterium tuberculosis WhiB3: A novel iron-sulfur cluster protein that regulates redox homeostasis and virulence. Antioxid. Redox Signal. 2012, 16, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Gazdik, M.A.; Mcdonough, K.A. Identification of cyclic AMP-regulated genes in Mycobacterium tuberculosis complex bacteria under low-oxygen conditions identification of cyclic AMP-regulated genes in Mycobacterium tuberculosis complex bacteria under low-oxygen conditions. J. Bacteriol. 2005, 187, 2681–2692. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.; Yuen, A.; Singh, V.; Hmama, Z. Mycobacterium tuberculosis Cpn60. 2 (GroEL2) blocks macrophage apoptosis via interaction with mitochondrial mortalin. Biol. Open 2017, 2, 481–488. [Google Scholar] [CrossRef] [PubMed]
- Naffin-Olivos, J.L.; Georgieva, M.; Goldfarb, N.; Madan-Lala, R.; Dong, L.; Bizzell, E.; Valinetz, E.; Brandt, G.S.; Yu, S.; Shabashvili, D.E.; et al. Mycobacterium tuberculosis Hip1 modulates macrophage responses through proteolysis of GroEL2. PLoS Pathog. 2014, 10. [Google Scholar] [CrossRef] [PubMed]
- Forrellad, M.A.; Klepp, L.I.; Gioffré, A.; Sabio y García, J.; Morbidoni, H.R.; de la Paz Santangelo, M.; Cataldi, A.A.; Bigi, F. Virulence factors of the Mycobacterium tuberculosis complex. Virulence 2013, 4, 3–66. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, S.; Midha, M.K.; Verma, H.N.; Basu, A.; Rao, K.V.S.; Manivel, V. Characterizing virulence-specific perturbations in the mitochondrial function of macrophages infected with Mycobacterium tuberculosis. Sci. Rep. 2013, 3, 1328. [Google Scholar] [CrossRef] [PubMed]
- Stewart, G.R.; Wernisch, L.; Stabler, R.; Mangan, J.A.; Hinds, J.; Laing, K.G.; Young, D.B.; Butcher, P.D. Dissection of the heat-shock response in Mycobacterium tuberculosis using mutants and microarrays. Microbiology 2002, 148, 3129–3138. [Google Scholar] [CrossRef] [PubMed]
- Agarwal, N.; Raghunand, T.R.; Bishai, W.R. Regulation of the expression of whiB1 in Mycobacterium tuberculosis: Role of cAMP receptor protein. Microbiology 2006, 152, 2749–2756. [Google Scholar] [CrossRef] [PubMed]
- Gazdik, M.A.; Bai, G.; Wu, Y.; McDonough, K.A. Rv1675c (cmr) regulates intramacrophage and cyclic AMP-induced gene expression in Mycobacterium tuberculosis-complex mycobacteria. Mol. Microbiol. 2009, 71, 434–448. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Manoranjan, J.; Pan, M.; Bohsali, A.; Xu, J.; Liu, J.; McDonald, K.L.; Szyk, A.; LaRonde-LeBlanc, N.; Gao, L.-Y. Evidence for pore formation in host cell membranes by ESX-1-secreted ESAT-6 and its role in Mycobacterium marinum escape from the vacuole. Infect. Immun. 2008, 76, 5478–5487. [Google Scholar] [CrossRef] [PubMed]
- Lim, Y.-J.; Choi, H.-H.; Choi, J.-A.; Jeong, J.A.; Cho, S.-N.; Lee, J.-H.; Park, J.B.; Kim, H.-J.; Song, C.-H. Mycobacterium kansasii-induced death of murine macrophages involves endoplasmic reticulum stress responses mediated by reactive oxygen species generation or calpain activation. Apoptosis 2013, 18, 150–159. [Google Scholar] [CrossRef] [PubMed]
- Mba Medie, F.; Champion, M.M.; Williams, E.A.; Champion, P.A.D. Homeostasis of N-α-terminal acetylation of EsxA correlates with virulence in Mycobacterium marinum. Infect. Immun. 2014, 82, 4572–4586. [Google Scholar] [CrossRef] [PubMed]
- Augenstreich, J.; Simeone, R.; Wegener, A.; Sayes, F.; Le, F.; Christian, C.; Malaga, W.; Guilhot, C.; Brosch, R.; Astarie, C. ESX-1 and phthiocerol dimycocerosates of Mycobacterium tuberculosis act in concert to cause phagosomal rupture and host cell apoptosis. Cell. Microbiol. 2017, 1–19. [Google Scholar] [CrossRef] [PubMed]
- Francis, R.J.; Butler, R.E.; Stewart, G.R. Mycobacterium tuberculosis ESAT-6 is a leukocidin causing Ca2+ influx, necrosis and neutrophil extracellular trap formation. Cell Death Dis. 2014, 5, e1474. [Google Scholar] [CrossRef] [PubMed]
- Jamwal, S.V.; Mehrotra, P.; Singh, A.; Siddiqui, Z.; Basu, A. Mycobacterial escape from macrophage phagosomes to the cytoplasm represents an alternate adaptation mechanism. Sci. Rep. 2016, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Clemmensen, H.S.; Peter, N.; Knudsen, H.; Rasmussen, E.M.; Winkler, J.; Rosenkrands, I.; Ahmad, A.; Lillebaek, T.; Sherman, D.R.; Andersen, P.L.; et al. An attenuated Mycobacterium tuberculosis clinical strain with a defect in ESX-1 secretion induces minimal host immune responses and pathology. Sci. Rep. 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Deng, W.; Xiang, X.; Xie, J. Comparative genomic and proteomic anatomy of Mycobacterium ubiquitous Esx family proteins: Implications in pathogenicity and virulence. Curr. Microbiol. 2014, 68, 558–567. [Google Scholar] [CrossRef] [PubMed]
- Di Giuseppe Champion, P.A.; Champion, M.M.; Manzanillo, P.; Cox, J.S. ESX-1 secreted virulence factors are recognized by multiple cytosolic AAA ATPases in mycobactria. Mol. Microbiol. 2009, 73, 950–962. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.M.; Boy-Röttger, S.; Dhar, N.; Sweeney, N.; Buxton, R.S.; Pojer, F.; Rosenkrands, I.; Cole, S.T. EspD is critical for the virulence-mediating ESX-1 secretion system in Mycobacterium tuberculosis. J. Bacteriol. 2012, 194, 884–893. [Google Scholar] [CrossRef] [PubMed]
- Ize, B.; Palmer, T. Microbiology. Mycobacteria’s export strategy. Science 2006, 313, 1583–1584. [Google Scholar] [CrossRef] [PubMed]
- Fortune, S.M.; Jaeger, A.; Sarracino, D.A.; Chase, M.R.; Sassetti, C.M.; Sherman, D.R.; Bloom, B.R.; Rubin, E.J. Mutually dependent secretion of proteins required for mycobacterial virulence. Proc. Natl. Acad. Sci. USA 2005, 102, 10676–10681. [Google Scholar] [CrossRef] [PubMed]
- Garces, A.; Atmakuri, K.; Chase, M.R.; Woodworth, J.S.; Rothchild, A.C.; Ramsdell, T.L.; Lopez, M.F.; Behar, S.M.; Sarracino, D.A.; Fortune, S.M. EspA acts as a critical mediator of ESX1-dependent virulence in Mycobacterium tuberculosis by affecting bacterial cell wall integrity. PLoS Pathog. 2010, 6, 15–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ates, L.S.; Brosch, R. Micro commentary discovery of the type VII ESX-1 secretion needle? Mol. Microbiol. 2017, 103, 7–12. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Rybniker, J.; Sala, C.; Cole, S.T. EspC forms a filamentous structure in the cell envelope of Mycobacterium tuberculosis and impacts ESX-1 secretion. Mol. Microbiol. 2017, 103, 26–38. [Google Scholar] [CrossRef] [PubMed]
- Ohol, Y.M.; Goetz, D.H.; Chan, K.; Shiloh, M.U.; Charles, C.S.; Cox, J.S. Mycobacterium tuberculosis MycP1 protease plays a dual role in regulation of ESX-1 secretion and virulence. Cell Host Microbe 2011, 7, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Van Winden, V.J.C.; Ummels, R.; Piersma, S.R.; Jiménez, C.R.; Korotkov, K.V.; Bitter, W. Mycosins are required for the stabilization of the ESX-1 and ESX-5. mBio 2016, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Sinha, A.; Gupta, S.; Bhutani, S.; Pathak, A.; Sarkar, D. PhoP-PhoP interaction at adjacent PhoP binding sites is influenced by protein phosphorylation. J. Bacteriol. 2008, 190, 1317–1328. [Google Scholar] [CrossRef] [PubMed]
- Pérez, E.; Samper, S.; Bordas, Y.; Guilhot, C.; Gicquel, B.; Martín, C. An essential role for phoP in Mycobacterium tuberculosis virulence. Mol. Microbiol. 2001, 41, 179–187. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Kumar, V.A. Mycobacterium tuberculosis virulence-regulator PhoP interacts with alternative sigma factor SigE during acid-stress response. Mol. Microbiol. 2017, 104, 400–411. [Google Scholar] [CrossRef] [PubMed]
- Singh, R.; Singh, M.; Arora, G.; Kumar, S.; Tiwari, P.; Kidwai, S. Polyphosphate deficiency in Mycobacterium tuberculosis is associated with enhanced drug susceptibility and impaired growth in guinea pigs. J. Bacteriol. 2013, 195, 2839–2851. [Google Scholar] [CrossRef] [PubMed]
- Sureka, K.; Dey, S.; Datta, P.; Singh, A.K.; Dasgupta, A.; Rodrigue, S.; Basu, J.; Kundu, M. Polyphosphate kinase is involved in stress-induced mprAB-sigE-rel signalling in mycobacteria. Mol. Microbiol. 2007, 65, 261–276. [Google Scholar] [CrossRef] [PubMed]
- Bretl, D.J.; Demetriadou, C.; Zahrt, T.C. Adaptation to environmental stimuli within the host: Two-component signal transduction systems of Mycobacterium tuberculosis. Microbiol. Mol. Biol. Rev. 2011, 75, 566–582. [Google Scholar] [CrossRef] [PubMed]
- Kumar, V.A.; Goyal, R.; Bansal, R.; Singh, N.; Sevalkar, R.R.; Kumar, A.; Sarkar, D. EspR-dependent ESAT-6 protein secretion of Mycobacterium tuberculosis requires the presence of virulence regulator PhoP. J. Biol. Chem. 2016, 291, 19018–19030. [Google Scholar] [CrossRef] [PubMed]
- Gröschel, M.I.; Sayes, F.; Simeone, R.; Majlessi, L.; Brosch, R. ESX secretion systems: Mycobacterial evolution to counter host immunity. Nat. Rev. Microbiol. 2016, 14, 677–691. [Google Scholar] [CrossRef] [PubMed]
- Bitter, W.; Houben, E.N.G.; Bottai, D.; Brodin, P.; Brown, E.J.; Cox, J.S.; Derbyshire, K.; Fortune, S.M.; Gao, L.-Y.; Liu, J.; et al. Systematic genetic nomenclature for type VII secretion systems. PLoS Pathog. 2009, 5, e1000507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kahramanoglou, C.; Cortes, T.; Matange, N.; Hunt, D.M.; Visweswariah, S.S.; Young, D.B.; Buxton, R.S. Genomic mapping of cAMP receptor protein (CRPMt) in Mycobacterium tuberculosis: Relation to transcriptional start sites and the role of CRPMt as a transcription factor. Nucleic Acids Res. 2014, 42, 8320–8329. [Google Scholar] [CrossRef] [PubMed]
- De Jonge, M.I.; Pehau-Arnaudet, G.; Fretz, M.M.; Romain, F.; Bottai, D.; Brodin, P.; Honoré, N.; Marchal, G.; Jiskoot, W.; England, P.; et al. ESAT-6 from Mycobacterium tuberculosis dissociates from its putative chaperone CFP-10 under acidic conditions and exhibits membrane-lysing activity. J. Bacteriol. 2007, 189, 6028–6034. [Google Scholar] [CrossRef] [PubMed]
- Blasco, B.; Chen, J.M.; Hartkoorn, R.; Sala, C.; Uplekar, S.; Rougemont, J.; Pojer, F.; Cole, S.T. Virulence regulator EspR of Mycobacterium tuberculosis is a nucleoid-associated protein. PLoS Pathog. 2012, 8, e1002621. [Google Scholar] [CrossRef] [PubMed]
- Trauner, A.; Lougheed, K.E.A.; Bennett, M.H.; Hingley-Wilson, S.M.; Williams, H.D. The dormancy regulator DosR controls ribosome stability in hypoxic mycobacteria. J. Biol. Chem. 2012, 287, 24053–24063. [Google Scholar] [CrossRef] [PubMed]
- Raman, K.; Bhat, A.G.; Chandra, N. A systems perspective of host-pathogen interactions: Predicting disease outcome in tuberculosis. Mol. Biosyst. 2010, 6, 516–530. [Google Scholar] [CrossRef] [PubMed]
- Marino, S.; El-Kebir, M.; Kirschner, D. A hybrid multi-compartment model of granuloma formation and T cell priming in tuberculosis. J. Theor. Biol. 2011, 280, 50–62. [Google Scholar] [CrossRef] [PubMed]
- Leistikow, R.L.; Morton, R.A.; Bartek, I.L.; Frimpong, I.; Wagner, K.; Voskuil, M.I. The Mycobacterium tuberculosis DosR regulon assists in metabolic homeostasis and enables rapid recovery from nonrespiring dormancy. J. Bacteriol. 2010, 192, 1662–1670. [Google Scholar] [CrossRef] [PubMed]
- Gautam, U.S.; Chauhan, S.; Tyagi, J.S. Determinants outside the DevR C-terminal domain are essential for cooperativity and robust activation of dormancy genes in Mycobacterium tuberculosis. PLoS ONE 2011, 6, e16500. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.; Tyagi, J.S. Cooperative binding of phosphorylated DevR to upstream sites is necessary and sufficient for activation of the Rv3134c-devRS operon in Mycobacterium tuberculosis: Implication in the induction of DevR target genes. J. Bacteriol. 2008, 190, 4301–4312. [Google Scholar] [CrossRef] [PubMed]
- Chauhan, S.; Sharma, D.; Singh, A.; Surolia, A.; Tyagi, J.S. Comprehensive insights into Mycobacterium tuberculosis DevR (DosR) regulon activation switch. Nucleic Acids Res. 2011, 39, 7400–7414. [Google Scholar] [CrossRef] [PubMed]
- Sharma, S.; Tyagi, J.S. Mycobacterium tuberculosis DevR/DosR dormancy regulator activation mechanism: Dispensability of phosphorylation, cooperativity and essentiality of α10 Helix. PLoS ONE 2016, 11, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Toledo, J.C.; Patel, R.P.; Lancaster, J.R.; Steyn, A.J.C. Mycobacterium tuberculosis DosS is a redox sensor and DosT is a hypoxia sensor. Proc. Natl. Acad. Sci. USA 2007, 104, 11568–11573. [Google Scholar] [CrossRef] [PubMed]
- Honaker, R.W.; Leistikow, R.L.; Bartek, I.L.; Voskuil, M.I. Unique roles of DosT and DosS in DosR regulon induction and Mycobacterium tuberculosis dormancy. Infect. Immun. 2009, 77, 3258–3263. [Google Scholar] [CrossRef] [PubMed]
- Kaur, K.; Kumari, P.; Sharma, S.; Sehgal, S.; Tyagi, J.S. DevS/DosS sensor is bifunctional and its phosphatase activity precludes aerobic DevR/DosR regulon expression in Mycobacterium tuberculosis. FEBS J. 2016, 283, 2949–2962. [Google Scholar] [CrossRef] [PubMed]
- Honaker, R.W.; Dhiman, R.K.; Narayanasamy, P.; Crick, D.C.; Voskuil, M.I. DosS responds to a reduced electron transport system to induce the Mycobacterium tuberculosis DosR regulon. J. Bacteriol. 2010, 192, 6447–6455. [Google Scholar] [CrossRef] [PubMed]
- Kumar, A.; Deshane, J.S.; Crossman, D.K.; Bolisetty, S.; Yan, B.-S.; Kramnik, I.; Agarwal, A.; Steyn, A.J.C. Heme oxygenase-1-derived carbon monoxide induces the Mycobacterium tuberculosis dormancy regulon. J. Biol. Chem. 2008, 283, 18032–18039. [Google Scholar] [CrossRef] [PubMed]
- Silva-Gomes, S.; Appelberg, R.; Larsen, R.; Soares, M.P.; Gomes, M.S. Heme catabolism by heme oxygenase-1 confers host resistance to Mycobacterium infection. Infect. Immun. 2013, 81, 2536–2545. [Google Scholar] [CrossRef] [PubMed]
- Bunker, R.D.; Mandal, K.; Bashiri, G.; Chaston, J.J.; Pentelute, B.L.; Lott, J.S.; Kent, S.B.H.; Baker, E.N. A functional role of Rv1738 in Mycobacterium tuberculosis persistence suggested by racemic protein crystallography. Proc. Natl. Acad. Sci. USA. 2015, 112, 4310–4315. [Google Scholar] [CrossRef] [PubMed]
- Maris, A.E.; Sawaya, M.R.; Kaczor-Grzeskowiak, M.; Jarvis, M.R.; Bearson, S.M.D.; Kopka, M.L.; Schröder, I.; Gunsalus, R.P.; Dickerson, R.E. Dimerization allows DNA target site recognition by the NarL response regulator. Nat. Struct. Biol. 2002, 9, 771–778. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.-N.; Jung, K.-E.; Ko, I.-J.; Baik, H.S.; Oh, J.-I. Protein-protein interactions between histidine kinases and response regulators of Mycobacterium tuberculosis H37Rv. J. Microbiol. 2012, 50, 270–277. [Google Scholar] [CrossRef] [PubMed]
- Jung, J.-Y.; Madan-Lala, R.; Georgieva, M.; Rengarajan, J.; Sohaskey, C.D.; Bange, F.-C.; Robinson, C.M. The intracellular environment of human macrophages that produce nitric oxide promotes growth of mycobacteria. Infect. Immun. 2013, 81, 3198–3209. [Google Scholar] [CrossRef] [PubMed]
- Nambu, S.; Matsui, T.; Goulding, C.W.; Takahashi, S.; Ikeda-Saito, M. A new way to degrade heme: The Mycobacterium tuberculosis enzyme MhuD catalyzes heme degradation without generating CO. J. Biol. Chem. 2013, 288, 10101–10109. [Google Scholar] [CrossRef] [PubMed]
- Arya, S.; Sethi, D.; Singh, S.; Hade, M.D.; Singh, V.; Raju, P.; Chodisetti, S.B.; Verma, D.; Varshney, G.C.; Agrewala, J.N.; et al. Truncated hemoglobin, HbN, is post-translationally modified in Mycobacterium tuberculosis and modulates host-pathogen interactions during intracellular infection. J. Biol. Chem. 2013, 288, 29987–29999. [Google Scholar] [CrossRef] [PubMed]
- Tullius, M.V.; Harmston, C.A.; Owens, C.P.; Chim, N.; Morse, R.P.; McMath, L.M.; Iniguez, A.; Kimmey, J.M.; Sawaya, M.R.; Whitelegge, J.P.; et al. Discovery and characterization of a unique mycobacterial heme acquisition system. Proc. Natl. Acad. Sci. USA 2011, 108, 5051–5056. [Google Scholar] [CrossRef] [PubMed]
- Joseph, S.V.; Madhavilatha, G.K.; Kumar, R.A.; Mundayoor, S. Comparative analysis of mycobacterial truncated hemoglobin promoters and the groEL2 promoter in free-living and intracellular mycobacteria. Appl. Environ. Microbiol. 2012, 78, 6499–6506. [Google Scholar] [CrossRef] [PubMed]
- Sethi, D.; Mahajan, S.; Singh, C.; Lama, A.; Hade, M.D.; Gupta, P.; Dikshit, K.L. Lipoprotein LprI of Mycobacterium tuberculosis acts as a lysozyme inhibitor. J. Biol. Chem. 2016, 291, 2938–2953. [Google Scholar] [CrossRef] [PubMed]
- Phetsuksiri, B.; Baulard, A.R.; Cooper, A.M.; Minnikin, D.E.; Douglas, J.D.; Besra, G.S.; Brennan, P.J. Antimycobacterial activities of isoxyl and new derivatives through the inhibition of mycolic acid synthesis antimycobacterial activities of isoxyl and new derivatives through the inhibition of mycolic acid synthesis. Antimicrob. Agents Chemother. 1999, 43, 1042–1051. [Google Scholar] [PubMed]
- Hall, G.; Bradshaw, T.D.; Laughton, C.A.; Stevens, M.F.; Emsley, J. Structure of Mycobacterium tuberculosis thioredoxin in complex with quinol inhibitor PMX464. Protein Sci. 2011, 20, 210–215. [Google Scholar] [CrossRef] [PubMed]
- Ascenzi, P.; Coletta, A.; Cao, Y.; Trezza, V.; Leboffe, L.; Fanali, G.; Fasano, M.; Pesce, A.; Ciaccio, C.; Marini, S.; et al. Isoniazid inhibits the heme-based reactivity of Mycobacterium tuberculosis truncated hemoglobin N. PLoS ONE 2013, 8, e69762. [Google Scholar] [CrossRef] [PubMed]
- Tan, M.P.; Sequeira, P.; Lin, W.W.; Phong, W.Y.; Cliff, P.; Ng, S.H.; Lee, B.H.; Camacho, L.; Schnappinger, D.; Ehrt, S.; et al. Nitrate respiration protects hypoxic Mycobacterium tuberculosis against acid- and reactive nitrogen species stresses. PLoS ONE 2010, 5, e13356. [Google Scholar] [CrossRef] [PubMed]
- Khan, A.; Sarkar, D. Nitrate reduction pathways in mycobacteria and their implications during latency. Microbiology 2012, 158, 301–307. [Google Scholar] [CrossRef] [PubMed]
- Rustad, T.R.; Harrell, M.I.; Liao, R.; Sherman, D.R. The enduring hypoxic response of Mycobacterium tuberculosis. PLoS ONE 2008, 3, e1502. [Google Scholar] [CrossRef] [PubMed]
- Veatch, A.V.; Kaushal, D. Opening Pandora’s Box: Mechanisms of Mycobacterium tuberculosis Resuscitation. Trends Microbiol. 2017, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Sun, F.; Yang, H. CRP acts as a transcriptional repressor of the YPO1635- phoPQ-YPO1632 operon in Yersinia pestis. Curr. Microbiol. 2015, 70, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Gupta, R.K.; Srivastava, B.S.; Srivastava, R. Comparative expression analysis of rpf-like genes of Mycobacterium tuberculosis H37Rv under different physiological stress and growth conditions. Microbiology 2010, 156, 2714–2722. [Google Scholar] [CrossRef] [PubMed]
- Bai, G.; Mccue, L.A.; Mcdonough, K.A.; Acteriol, J.B. Characterization of Mycobacterium tuberculosis Rv3676 (CRP Mt), a cyclic AMP receptor protein-like DNA binding protein. J. Bacteriol. 2005, 187, 7795–7804. [Google Scholar] [CrossRef] [PubMed]
- Rickman, L.; Scott, C.; Hunt, D.M.; Hutchinson, T.; Menéndez, M.C.; Whalan, R.; Hinds, J.; Colston, M.J.; Green, J.; Buxton, R.S. A member of the cAMP receptor protein family of transcription regulators in Mycobacterium tuberculosis is required for virulence in mice and controls transcription of the rpfA gene coding for a resuscitation promoting factor. Mol. Microbiol. 2005, 56, 1274–1286. [Google Scholar] [CrossRef] [PubMed]
- Bretl, D.J.; He, H.; Demetriadou, C.; White, M.J.; Penoske, R.M.; Salzman, N.H.; Zahrt, T.C. MprA and DosR coregulate a Mycobacterium tuberculosis virulence operon encoding Rv1813c and Rv1812c. Infect. Immun. 2012, 80, 3018–3033. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Cao, G.; Neuenschwander, P.F.; Haydel, S.E.; Hou, G.; Howard, S.T. The β-propeller gene Rv1057 of Mycobacterium tuberculosis has a complex promoter directly regulated by both the MprAB and TrcRS two-component systems. Tuberculosis 2011, 91, S142–S149. [Google Scholar] [CrossRef] [PubMed]
- Pang, X.; Vu, P.; Byrd, T.F.; Ghanny, S.; Soteropoulos, P.; Mukamolova, G.V.; Wu, S.; Samten, B.; Howard, S.T. Evidence for complex interactions of stress-associated regulons in an mprAB deletion mutant of Mycobacterium tuberculosis. Microbiology 2007, 153, 1229–1242. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Y.; Wang, L.; Han, Y.; Yan, Y.; Tan, Y.; Zhou, L.; Cui, Y.; Du, Z.; Wang, X.; Bi, Y.; et al. Autoregulation of PhoP/PhoQ and positive regulation of the cyclic AMP receptor protein-cyclic AMP complex by PhoP in Yersinia pestis. J. Bacteriol. 2013, 195, 1022–1030. [Google Scholar] [CrossRef] [PubMed]
- Jofré, M.R.; Rodríguez, L.M.; Villagra, N.A.; Hidalgo, A.A.; Mora, G.C.; Fuentes, J.A. RpoS integrates CRP, Fis, and PhoP signaling pathways to control Salmonella Typhi hlyE expression. BMC Microbiol. 2014, 14, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Brown, M.R.W.; Kornberg, A. Inorganic polyphosphate in the origin and survival of species. Proc. Natl. Acad. Sci. USA 2004, 101, 16085–16087. [Google Scholar] [CrossRef] [PubMed]
- Chuang, Y.; Dutta, N.K.; Hung, C.; Wu, T.; Rubin, H.; Karakousis, C. Stringent response factors PPX1 and PPK2 play an important role in Mycobacterium tuberculosis metabolism, biofilm formation, and sensitivity to isoniazid in vivo. Antimicrob. Agents Chemother. 2016, 60, 6460–6470. [Google Scholar] [CrossRef] [PubMed]
- Sanyal, S.; Banerjee, S.K.; Banerjee, R.; Mukhopadhyay, J.; Kundu, M. Polyphosphate kinase 1, a central node in the stress response network of Mycobacterium tuberculosis, connects the two-component systems MprAB and SenX3-RegX3 and the extracytoplasmic function sigma factor, sigma E. Microbiology 2013, 159, 2074–2086. [Google Scholar] [CrossRef] [PubMed]
- Manganelli, R.; Provvedi, R. An integrated regulatory network including two positive feedback loops to modulate the activity of SigE in mycobacteria. Mol. Microbiol. 2010, 75, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Troudt, J.; Creissen, E.; Izzo, L.; Bielefeldt-ohmann, H.; Casonato, S.; Manganelli, R.; Izzo, A.A. Mycobacterium tuberculosis sigE mutant ST28 used as a vaccine induces protective immunity in the guinea pig model. Tuberculosis 2017, 106, 99–105. [Google Scholar] [CrossRef] [PubMed]
- VanderVen, B.C.; Fahey, R.J.; Lee, W.; Liu, Y.; Abramovitch, R.B.; Memmott, C.; Crowe, A.M.; Eltis, L.D.; Perola, E.; Deininger, D.D.; et al. Novel inhibitors of cholesterol degradation in Mycobacterium tuberculosis reveal how the bacterium’s metabolism is constrained by the intracellular environment. PLoS Pathog. 2015, 11, e1004679. [Google Scholar] [CrossRef] [PubMed]
- Los, F.C.O.; Randis, T.M.; Aroian, R.V.; Ratner, A.J. Role of pore-forming toxins in bacterial infectious diseases. Microbiol. Mol. Biol. Rev. 2013, 77, 173–207. [Google Scholar] [CrossRef] [PubMed]
- Gatfield, J.; Pieters, J. Essential role for cholesterol in entry of mycobacteria into macrophages. Science 2000, 288, 1647–1650. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | IdeR | Zur | MntR |
|---|---|---|---|
| esx3-operon 1 | − | − | − |
| esxG-esxH | − | − | − |
| esxQ | − | ||
| esxR-esxS | − | − | |
| esxW | − | ||
| ppe3 | − | − | |
| ppe4-pe5 | − | − | − |
| ppe9 | + | ||
| pe13 | 2 | − | |
| ppe19 | − | ||
| ppe20 | − | ||
| ppe37 | − | ||
| ppe38 | 2 | ||
| ppe48 | − | ||
| pe_pgrs61 | − |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zondervan, N.A.; Van Dam, J.C.J.; Schaap, P.J.; Martins dos Santos, V.A.P.; Suarez-Diez, M. Regulation of Three Virulence Strategies of Mycobacterium tuberculosis: A Success Story. Int. J. Mol. Sci. 2018, 19, 347. https://doi.org/10.3390/ijms19020347
Zondervan NA, Van Dam JCJ, Schaap PJ, Martins dos Santos VAP, Suarez-Diez M. Regulation of Three Virulence Strategies of Mycobacterium tuberculosis: A Success Story. International Journal of Molecular Sciences. 2018; 19(2):347. https://doi.org/10.3390/ijms19020347
Chicago/Turabian StyleZondervan, Niels A., Jesse C. J. Van Dam, Peter J. Schaap, Vitor A. P. Martins dos Santos, and Maria Suarez-Diez. 2018. "Regulation of Three Virulence Strategies of Mycobacterium tuberculosis: A Success Story" International Journal of Molecular Sciences 19, no. 2: 347. https://doi.org/10.3390/ijms19020347