Peripheral B-Cell Subset Distribution in Primary Antiphospholipid Syndrome

 and
and
Abstract
:1. Introduction
2. Results
2.1. Patients with pAPS Differ in the B-Cell Repertoire Compared to Patients with SLE
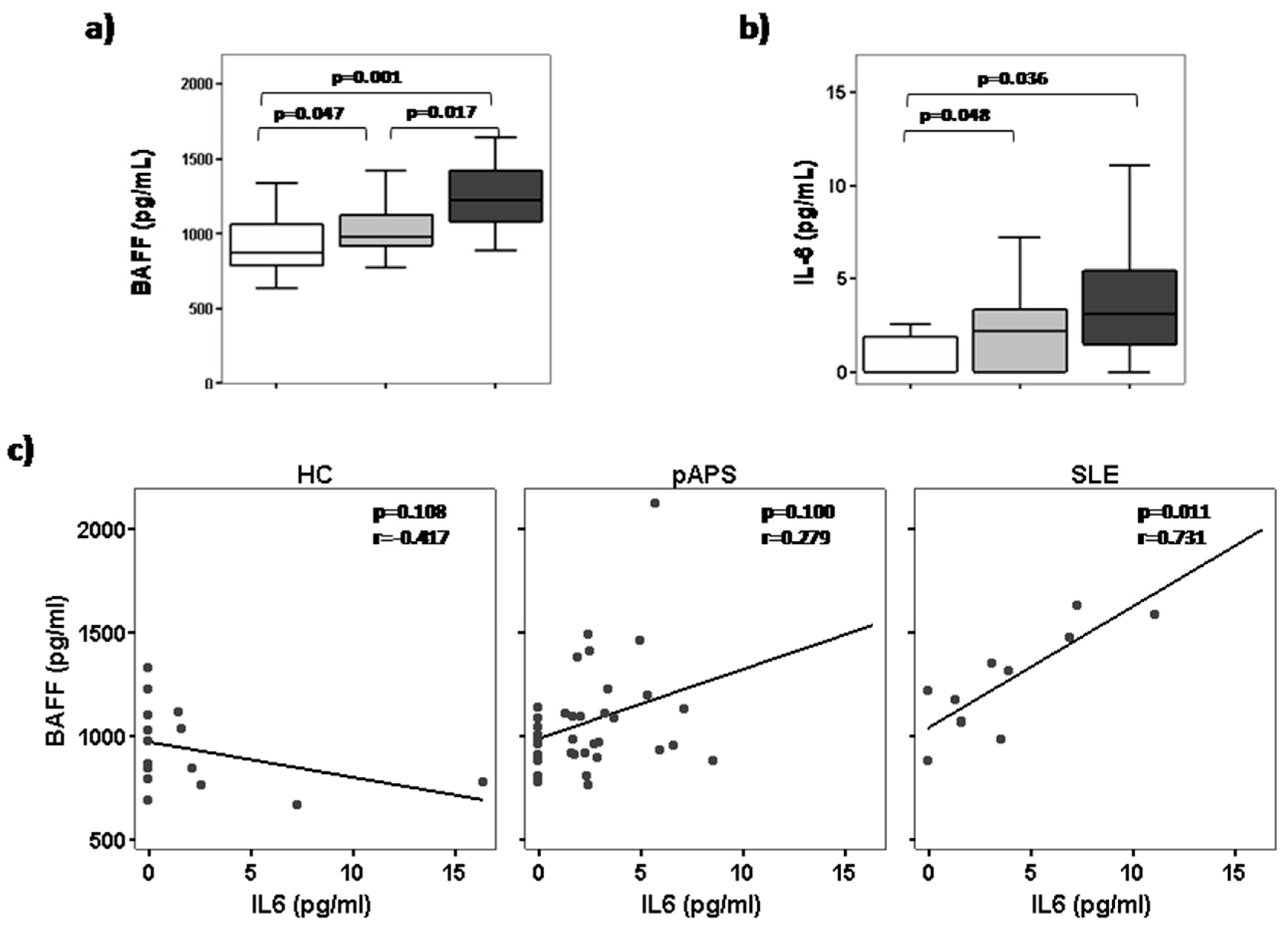
2.2. High Serum BAFF Levels Correlate with IL-6 in Patients with SLE but Not in pAPS Patients
3. Discussion
4. Materials and Methods
4.1. Subjects
4.2. Quantification of B Cells
4.3. Circulating B Cells Immunophenotyping
4.4. Serum BAFF Measurement
4.5. Detection of Soluble Cytokine in Serum
4.6. Statistical Analysis
Supplementary Materials
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Miyakis, S.; Lockshin, M.D.; Atsumi, T.; Branch, D.W.; Brey, R.L.; Cervera, R.; Derkesen, R.H.W.M.; de Groot, P.G.; Koike, T.; Meroni, P.L.; et al. International consensus statement on an update of the classification criteria for definite antiphospholipid syndrome (APS). J. Thromb. Haemost. 2006, 4, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Van den Hoogen, L.L.; van Roon, J.A.G.; Radstake, T.R.D.J.; Fritsch-Stork, R.D.E.; Derksen, R.H.W.M. Delineating the deranged immune system in the antiphospholipid syndrome. Autoimmun. Rev. 2016, 15, 50–60. [Google Scholar] [CrossRef] [PubMed]
- De Groot, P.G.; Urbanus, R.T. Antiphospholipid Syndrome—Not a Non inflammatory Disease. Semin. Thromb. Hemost. 2015, 41, 607–614. [Google Scholar] [PubMed]
- Manzi, S.; Ahearn, J.M.; Salmon, J. New insights into complement: A mediator of injury and marker of disease activity in systemic lupus erythematosus. Lupus 2004, 13, 298–303. [Google Scholar] [CrossRef] [PubMed]
- Abrahams, V.; Chamley, L.; Salmon, J. Emerging Treatment Models in Rheumatology: Antiphospholipid Syndrome and Pregnancy: Pathogenesis to Translation. Arthritis Rheumatol. 2017, 69, 1710–1721. [Google Scholar] [CrossRef] [PubMed]
- Arachchillage, D.R.J.; Laffan, M. Pathogenesis and management of antiphospholipid syndrome. Br. J. Haematol. 2017, 178, 181–195. [Google Scholar] [CrossRef] [PubMed]
- Yanaba, K.; Bouaziz, J.-D.; Matsushita, T.; Magro, C.M.; St. Clair, E.W.; Tedder, T.F. B-lymphocyte contributions to human autoimmune disease. Immunol. Rev. 2008, 223, 284–299. [Google Scholar] [CrossRef] [PubMed]
- Yurasov, S.; Wardemann, H.; Hammersen, J.; Tsuiji, M.; Meffre, E.; Pascual, V.; Nussenzweig, M.C. Defective B cell tolerance checkpoints in systemic lupus erythematosus. J. Exp. Med. 2005, 201, 703–711. [Google Scholar] [CrossRef] [PubMed]
- Kinnunen, T.; Chamberlain, N.; Morbach, H.; Cantaert, T.; Lynch, M.; Preston-Hurlburt, P.; Herold, K.C.; Hafler, D.A.; O’Connor, K.C.; Meffre, E. Specific peripheral B cell tolerance defects in patients with multiple sclerosis. J. Clin. Investig. 2013, 123, 2737–2741. [Google Scholar] [CrossRef] [PubMed]
- Glauzy, S.; Sng, J.; Bannock, J.; Gottenberg, J.E.; Korganow, A.S.; Cacoub, P.; Saadoun, D.M.E. Defective early B cell tolerance checkpoints in Sjögren’s Syndrome patients. Arthritis Rheumatol. 2017. [Google Scholar] [CrossRef] [PubMed]
- Carbone, J.; Gallego, A.; Lanio, N.; Navarro, J.; Orera, M.; Aguaron, A.; Fernandez-Cruz, E.; Sarmiento, E. Quantitative abnormalities of peripheral blood distinct T, B, and natural killer cell subsets and clinical findings in obstetric antiphospholipid syndrome. J. Rheumatol. 2009, 36, 1217–1225. [Google Scholar] [CrossRef] [PubMed]
- Simonin, L.; Pasquier, E.; Leroyer, C.; Cornec, D.; Lemerle, J.; Bendaoud, B.; Hillion, S.; Pers, J.O.; Couturaud, F.; Renaudineau, Y. Lymphocyte disturbances in primary antiphospholipid syndrome and application to venous thromboembolism follow-up. Clin. Rev. Allergy Immunol. 2016, 3878, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Youinou, P.; Renaudineau, Y. The antiphospholipid syndrome as a model for B cell-induced autoimmune diseases. Thromb. Res. 2004, 114, 363–369. [Google Scholar] [CrossRef] [PubMed]
- Schiemann, B. An essential role for baff in the normal development of B cells through a bcma-independent pathway. Science 2001, 293, 2111–2114. [Google Scholar] [CrossRef] [PubMed]
- Zharkova, O.; Celhar, T.; Cravens, P.D.; Satterthwaite, A.B.; Fairhurst, A.-M.; Davis, L.S. Pathways leading to an immunological disease: Systemic lupus erythematosus. Rheumatology 2017, 56, i55–i66. [Google Scholar] [CrossRef] [PubMed]
- Brink, R. Regulation of B cell self-tolerance by BAFF. Semin. Immunol. 2006, 18, 276–283. [Google Scholar] [CrossRef] [PubMed]
- Rawlings, D.J.; Metzler, G.; Wray-Dutra, M.; Jackson, S.W. Altered B cell signalling in autoimmunity. Nat. Rev. Immunol. 2017, 17, 421–436. [Google Scholar] [CrossRef] [PubMed]
- Chang, S.K.; Arendt, B.K.; Darce, J.R.; Wu, X.; Jelinek, D.F.; Blys, B. IMMUNOBIOLOGY A role for BLyS in the activation of innate immune cells. Blood 2006, 108, 2687–2695. [Google Scholar] [CrossRef] [PubMed]
- Bernales, I.; Fullaondo, A.; Marín-Vidalled, M.J.; Ucar, E.; Martínez-Taboada, V.; López-Hoyos, M.; Zubiaga, A.M. Innate immune response gene expression profiles characterize primary antiphospholipid syndrome. Genes Immun. 2008, 9, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Dal Ben, E.R.R.; do Prado, C.H.; Baptista, T.S.A.; Bauer, M.E.; Staub, H.L. Decreased levels of circulating CD4+CD25+Foxp3+ regulatory T cells in patients with primary antiphospholipid syndrome. J. Clin. Immunol. 2013, 33, 876–879. [Google Scholar] [CrossRef] [PubMed]
- Dal Ben, E.R.R.; do Prado, C.H.; Baptista, T.S.A.; Bauer, M.E.; Staub, H.L. Patients with systemic lupus erythematosus and secondary antiphospholipid syndrome have decreased numbers of circulating CD4+CD25+Foxp3+ treg and CD3−CD19+ B cells. Rev. Bras. Reumatol. 2014, 54, 241–246. [Google Scholar] [PubMed]
- Riancho-Zarrabeitia, L.; Daroca, G.; Muñoz, P.; López-Hoyos, M.; Haya, A.; Martínez-Taboada, V.M. Serological evolution in women with positive antiphospholipid antibodies. Semin. Arthritis Rheum. 2017. [Google Scholar] [CrossRef] [PubMed]
- Sims, G.P.; Ettinger, R.; Shirota, Y.; Yarboro, C.H.; Illei, G.G.; Lipsky, P.E.; B-cell, M. Identification and characterization of circulating human transitional B cells. Blood 2005, 105, 4390–4399. [Google Scholar] [CrossRef] [PubMed]
- Blair, P.A.; Noreña, L.Y.; Flores-Borja, F.; Rawlings, D.J.; Isenberg, D.A.; Ehrenstein, M.R.; Mauri, C. CD19+CD24hiCD38hi B Cells Exhibit Regulatory Capacity in Healthy Individuals but Are Functionally Impaired in Systemic Lupus Erythematosus Patients. Immunity 2010, 32, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Sanz, I.; Wei, C.; Lee, F.E.-H.; Anolik, J. Phenotypic and functional heterogeneity of human memory B cells. Semin. Immunol. 2008, 20, 67–82. [Google Scholar] [CrossRef] [PubMed]
- Kaminski, D.A.; Wei, C.; Qian, Y.; Rosenberg, A.F.; Sanz, I. Advances in human B cell phenotypic profiling. Front. Immunol. 2012, 3. [Google Scholar] [CrossRef] [PubMed]
- Anolik, J.H.; Looney, R.J.; Lund, F.E.; Randall, T.D.; Sanz, I. Insights into the heterogeneity of human B cells: Diverse functions, roles in autoimmunity, and use as therapeutic targets. Immunol. Res. 2009, 45, 144–158. [Google Scholar] [CrossRef] [PubMed]
- Korganow, A.S.; Knapp, A.M.; Nehme-Schuster, H.; Soulas-Sprauel, P.; Poindron, V.; Pasquali, J.L.; Martin, T. Peripheral B cell abnormalities in patients with systemic lupus erythematosus in quiescent phase: Decreased memory B cells and membrane CD19 expression. J. Autoimmun. 2010, 34, 426–434. [Google Scholar] [CrossRef] [PubMed]
- Nemazee, D. Mechanisms of central tolerance for B cells. Nat. Rev. Immunol. 2017, 17, 281–294. [Google Scholar] [CrossRef] [PubMed]
- Meffre, E. The establishment of early B cell tolerance in humans: Lessons from primary immunodeficiency diseases. Ann. N. Y. Acad. Sci. 2011, 1246, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Doreau, A.; Belot, A.; Bastid, J.; Riche, B.; Trescol-Biemont, M.-C.; Ranchin, B.; Fabien, N.; Cochat, P.; Pouteil-Noble, C.; Trolliet, P.; et al. Interleukin 17 acts in synergy with B cell-activating factor to influence B cell biology and the pathophysiology of systemic lupus erythematosus. Nat. Immunol. 2009, 10, 778–785. [Google Scholar] [CrossRef] [PubMed]
- Groom, J.; Mackay, F. B cells flying solo. Immunol. Cell Biol. 2008, 86, 40–46. [Google Scholar] [CrossRef] [PubMed]
- MacKay, F.; Figgett, W.A.; Saulep, D.; Lepage, M.; Hibbs, M.L. B-cell stage and context-dependent requirements for survival signals from BAFF and the B-cell receptor. Immunol. Rev. 2010, 237, 205–225. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.B.; Saulep-Easton, D.; Figgett, W.A.; Fairfax, K.A.; Mackay, F. The BAFF/APRIL system: Emerging functions beyond B cell biology and autoimmunity. Cytokine Growth Factor Rev. 2013, 24, 203–215. [Google Scholar] [CrossRef] [PubMed]
- Thien, M.; Phan, T.G.; Gardam, S.; Amesbury, M.; Basten, A.; MacKay, F.; Brink, R. Excess BAFF rescues self-reactive B cells from peripheral deletion and allows them to enter forbidden follicular and marginal zone niches. Immunity 2004, 20, 785–798. [Google Scholar] [CrossRef] [PubMed]
- Lesley, R.; Xu, Y.; Kalled, S.L.; Hess, D.M.; Schwab, S.R.; Shu, H.B.; Cyster, J.G. Reduced competitiveness of autoantigen-engaged B cells due to increased dependence on BAFF. Immunity 2004, 20, 441–453. [Google Scholar] [CrossRef]
- Stohl, W.; Metyas, S.; Tan, S.M.; Cheema, G.S.; Oamar, B.; Xu, D.; Roschke, V.; Wu, Y.; Baker, K.P.; Hilbert, D.M.B. Lymphocyte stimulator overexpression in patients with systemic lupus erythematosus: longitudinal observations. Arthritis Rheum. 2003, 48, 3475–3486. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Roschke, V.; Baker, K.P.; Wang, Z.; Alarcón, G.S.; Fessler, B.J.; Bastian, H.; Kimberly, R.P.; Zhou, T. Cutting edge: A role for B lymphocyte stimulator in systemic lupus erythematosus. J. Immunol. 2001, 166, 6–10. [Google Scholar] [CrossRef] [PubMed]
- Vincent, F.B.; Morand, E.F.; Schneider, P.; Mackay, F. The BAFF/APRIL system in SLE pathogenesis. Nat. Rev. Rheumatol. 2014, 10, 365–373. [Google Scholar] [CrossRef] [PubMed]
- Salazar-Camarena, D.C.; Ortiz-Lazareno, P.C.; Cruz, A.; Oregon-Romero, E.; Machado-Contreras, J.R.; Muñoz-Valle, J.F.; Orozco-López, M.; Marín-Rosales, M.; Palafox-Sánchez, C.A. Association of BAFF, APRIL serum levels, BAFF-R, TACI and BCMA expression on peripheral B-cell subsets with clinical manifestations in systemic lupus erythematosus. Lupus 2016, 25, 582–592. [Google Scholar] [CrossRef] [PubMed]
- Taher, T.E.; Bystrom, J.; Ong, V.H.; Isenberg, D.A.; Renaudineau, Y.; Abraham, D.J.; Mageed, R.A. Intracellular B lymphocyte signalling and the regulation of humoral immunity and autoimmunity. Clin. Rev. Allergy Immunol. 2017, 1–28. [Google Scholar] [CrossRef]
- Hochberg, M.C. Updating the American College of Rheumatology revised criteria for the classification of systemic lupus erythematosus. Arthritis Rheum. 1997, 40, 1725. [Google Scholar] [CrossRef] [PubMed]
- Petri, M.; Orbai, A.M.; Alarcón, G.S.; Gordon, C.; Merrill, J.T.; Fortin, P.R.; Bruce, I.N.; Isenberg, D.; Wallace, D.J.; Nived, O.; et al. Derivation and validation of the systemic lupus international collaborating clinics classification criteria for systemic lupus erythematosus. Arthritis Rheum. 2012, 64, 2677–2686. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| HC | pAPS | SLE | |
|---|---|---|---|
| Number of patients, n | 21 | 37 | 11 |
| Age (mean ± SD) | 40.3 ± 11.8 | 36.7 ± 9.9 | 32.8 ± 13.1 |
| Females, n (%) | 15 (71.4) | 30 (81.1) | 11 (100) |
| Clinical manifestations associated with APS, n (%) | 0 | 37 (100) | 0 |
| - Obstetrical events, n (%) | - | 17 (45.9) | - |
| - Arterial thrombosis, n (%) | - | 12 (32.4) | - |
| - Venous thrombosis, n (%) | - | 8 (21.6) | - |
| Positive aPL Serology, n (%) | 0 | 37 (100) | 6 (54.5) |
| - Positivity for one Ab, n (%) | - | 19 (51.3) | 4 (36.3) |
| - Positivity for two Ab, n (%) | - | 12 (32.4) | 0 (0) |
| - Positivity for three Ab, n (%) | - | 6 (16.2) | 2 (18.2) |
| Serological profile | |||
| - aCL, n (%) | - | 29 (78.4) | 5 (45.5) |
| - aβ2GPI, n (%) | - | 17 (45.9) | 2 (18.2) |
| - Lupus Anticoagulant, n (%) | - | 15 (40.5) | 3 (27.3) |
| Peripheral blood cells | |||
| Lymphocytes (cells/mm3) | 2211.00 (1838.0–2915.0) | 1852.00 (1545.0–2590.5) | 1721.00 (1082.0–2475.0) |
| CD19+ B cells (cells/mm3) | 226.50 (181.75–341.25) | 185.00 (140.50–296.00) | 112.00 (87.00–325.00) |
| CD19+ B cells (%) | 11.50 (8.25–13.75) | 10 (7.00–12.50) | 10 (7.00–12.00) |
| Treatment | |||
| - Antiplatelet, n (%) | 23 (62.2) | 4 (36.4) | |
| - Anticoagulant, n (%) | 16 (43.2) | 1 (9.1) | |
| - Corticosteroids, n (%) | 4 (10.8) | 3(27.3) | |
| - Antimalarials, n (%) | 5 (13.5) | 9 (81.8) | |
| - Immunosuppresors, n (%) | 0 (0) | 2 (18.2) |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Álvarez-Rodríguez, L.; Riancho-Zarrabeitia, L.; Calvo-Alén, J.; López-Hoyos, M.; Martínez-Taboada, V. Peripheral B-Cell Subset Distribution in Primary Antiphospholipid Syndrome. Int. J. Mol. Sci. 2018, 19, 589. https://doi.org/10.3390/ijms19020589
Álvarez-Rodríguez L, Riancho-Zarrabeitia L, Calvo-Alén J, López-Hoyos M, Martínez-Taboada V. Peripheral B-Cell Subset Distribution in Primary Antiphospholipid Syndrome. International Journal of Molecular Sciences. 2018; 19(2):589. https://doi.org/10.3390/ijms19020589
Chicago/Turabian StyleÁlvarez-Rodríguez, Lorena, Leyre Riancho-Zarrabeitia, Jaime Calvo-Alén, Marcos López-Hoyos, and Víctor Martínez-Taboada. 2018. "Peripheral B-Cell Subset Distribution in Primary Antiphospholipid Syndrome" International Journal of Molecular Sciences 19, no. 2: 589. https://doi.org/10.3390/ijms19020589