2.1.1. Synthesis of Neo-Glycoprobes Exposing Relevant Mammalian Carbohydrate Epitopes
We aimed to study the lectin-binding profile of PDC-109 using purified protein material from biological origin, hence initial work was focused on the synthesis, purification and validation of new glycoprobes displaying some of the most relevant glycotopes in the mammalian system (
Table 1).
All the glycopeptides were synthesized as previously reported by our group [
16] using oxime ligation between the oligosaccharides in
Table 1 and the peptide module
N[Me]-
O-Aoa-GFKKG at 25 and 20 mM, respectively. Ligation reactions were performed at pH 3.5 (NAc-hexoses) or pH 4.6 (hexoses) and 37 °C for 72 h, the conjugates purified by high performance liquid chromatography (HPLC) and characterized by mass spectrometry (MS) as detailed in Materials and Methods. In addition to choosing an optimal pH for every hexose type, the use of aniline as catalyst to reduce incubation time was evaluated, as some authors have reported significant yield improvements (nearly double in 3 h) in conjugations involving non-methylated aminoxyacetic acid (Aoa)-containing peptides and monosaccharides [
17]. However, with our
N[Me]-
O-Aoa-peptide the yield increased only marginally after 72 h incubation, except for reactions with Fuc-containing i.e., [Fuc-α1,(3,4,6)-GlcNAc] disaccharides (e.g., from 8% to 16% for Fuc-α1,4-GlcNAc).
A substantial problem during the production of the glycopeptides was the purification by reversed phase (RP) HPLC, as the small change in hydrophobicity between the glycopeptide product and unreacted
N[Me]-
O-Aoa-peptide precursor rendered separations difficult, particularly in conjugations with <20% conversion. Attempts to increase the retention time of unreacted peptide by addition of carbonyl scavengers such as formaldehyde and acetone, or the alkylating agent
N-ethylmaleimide [
18], did not improve resolution. An additional difficulty, also related to purification, was that residual (trifluoroacetic) acid from HPLC eluents caused degradation of the
N[Me]-
O-Aoa-peptides during lyophilization. This problem was conveniently solved by adjusting to pH 5 the glycoprobe solution immediately after collection. A further test of the suitability of the conjugation conditions involved the highly acid-labile trisaccharides containing terminal sialic (
N-acetyl neuraminic) acid (Sia), which could be successfully conjugated to the
N[Me]-
O-Aoa-peptide module at mildly acidic pH, purified and characterized by MS like other glycoconjugates. Finally, the long-term stability of all compounds was shown to be satisfactory (>95% unaltered after 2 years) provided they are stored in lyophilized form. This renders our glycopeptide probe approach useful for longitudinal studies in carbohydrate research.
2.1.2. Neo-Glycoprobe Evaluation by SPR Studies with Known Lectins
With neo-glycopeptides displaying the most relevant glycotopes of the mammalian system in hand, their functionality was validated in SPR interaction studies using plant lectins (
Table 1) with known selectivity for the indicated carbohydrate epitopes. Lectin solutions were flown across a surface with the immobilized cognate sugar unit. Ac-GFKKG-amide, a non-glycosylated probe, was used as reference surface, given that the previously described
N[Me]-
O-Aoa-GFKKG-amide [
16] displayed undesirable, non-specific binding with PDC-109.
Disaccharides with terminal β-galactose were first evaluated. As in our proof of concept publication [
16], a sensor chip with the same β-galactosides [i.e., Gal(β1-4)GlcNAc, Gal(β1-3)GlcNAc, Gal(β1-6)GlcNAc)] and the new Ac-GFKKG-amide reference surface was prepared. Interaction between β-galactose-specific lectin ECA and these β-galactosides yielded the same kinetic results [
16], hence confirmed sensor surface functionality.
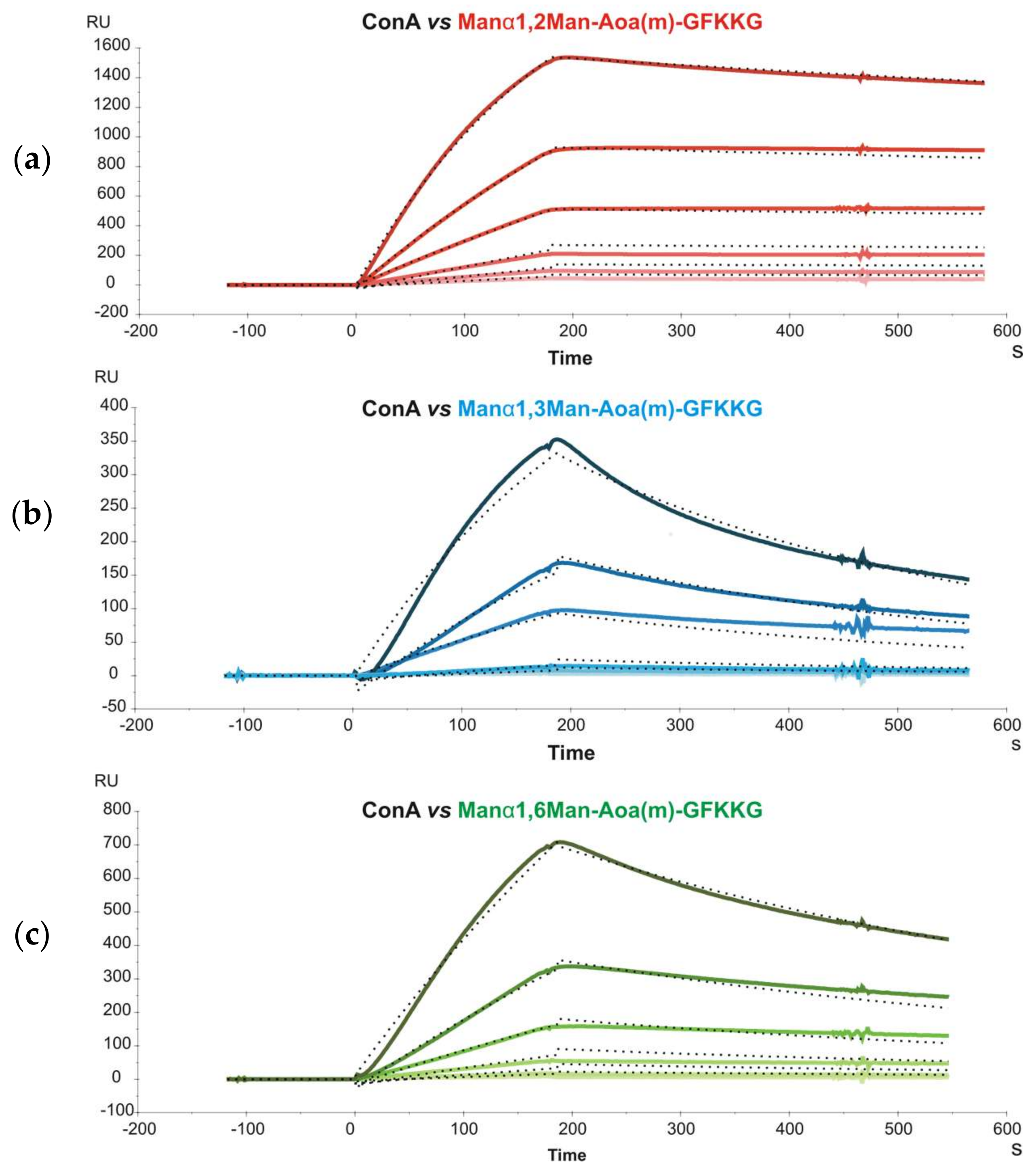
Subsequently, glycoprobes displaying mannobioses only differing in their glycosidic linkage (Man-α1,2-Man, Man-α1,3-Man and Man-α1,6-Man) were immobilized on a sensor chip and their interaction with
concanavalin A (Con A) was studied. Con A is a relatively complex lectin, organized as a β-barrel-like tetramer at pH > 7, each dimer subunit (D2 symmetry) with two CRDs situated on opposing faces of the protein. Sensorgrams of different Con A concentrations were recorded and kinetic constants determined by simultaneously fitting the experimental curves to a bivalent kinetic model (
Figure 1). This model allows the first and second binding events to be separately described by two sets of rate constants (
Table 2).
The unconventional response units (RU
−1) used to describe the second association event made comparison with constants derived from other methods (e.g., Isothermal Titration Calorimetry, ITC) difficult, and only the affinity constants for the first event, K
A1 (K
A1 = k
a1/k
d1) could be accurately compared. For this event, a higher affinity of Man-α1,3-Man over Man-α1,6-Man was found, in agreement with previous ITC data [
19], although the K
A1 values obtained with the ITC method were one order of magnitude lower (~10
4) than with our SPR approach (~10
5). One possible explanation for this difference, keeping in mind the two-CRDs-per-dimer model of Con A, is that although the bivalent binding model used in SPR only allows to derive standard-unit values for K
A1, the apparent k
d1, used for K
A1 determination is also influenced by the second event. If this contribution is not factored in, a lower k
d1, and consequently a higher K
A1, results.
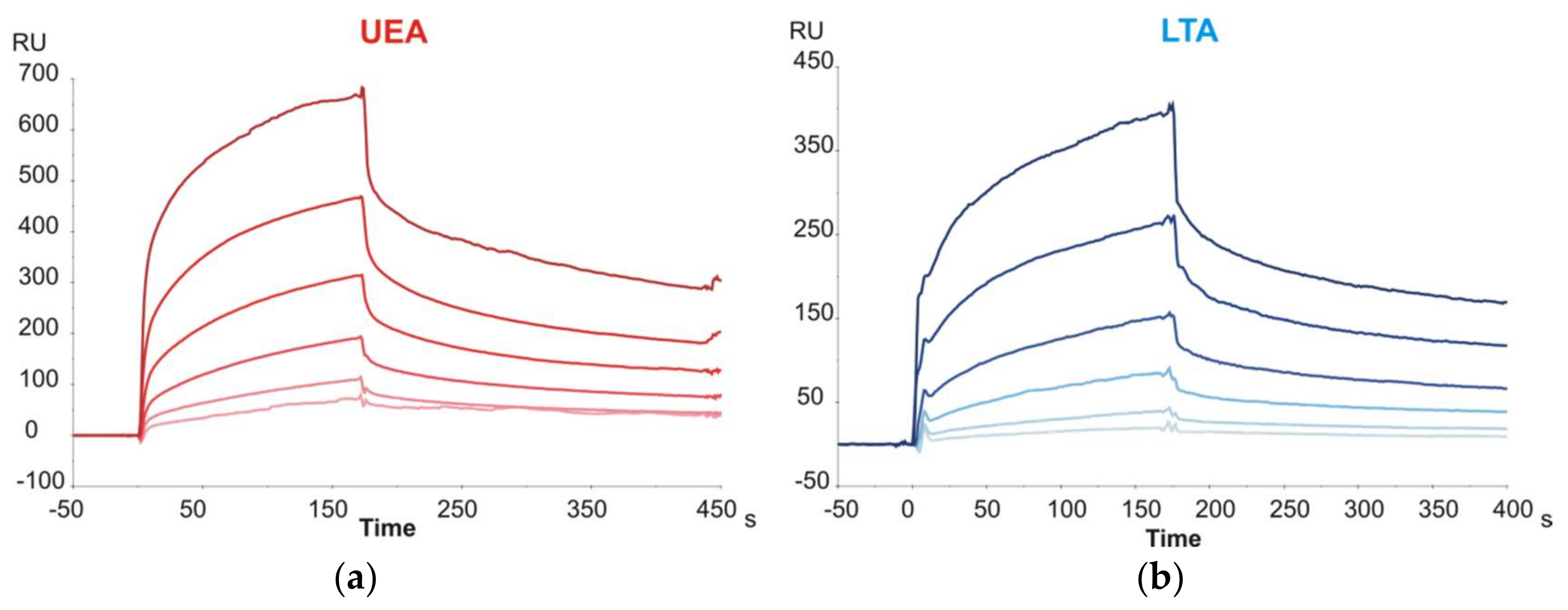
The last group of disaccharide glycoprobes to be evaluated included those with terminal Fuc units (Fuc-α1,3-GlcNAc, Fuc-α1,4-GlcNAc and Fuc-α1,6-GlcNAc). The latter epitope corresponds to the core-fucosylation of N-glycans, and the former two are partial Lewis
x and Lewis
a epitopes, respectively. The glycoprobe surfaces were tested against two Fuc-specific lectins from
Lotus tetragonolobus (LTA) and
Ulex europeaus (UEA). For both lectins, sensorgrams clearly demonstrated affinity for Fuc epitopes (
Figure 2; representative example for Fuc-α1,4-GlcNAc), even though the carbohydrate affinity for both lectins has been described to be strongly specific for the blood group H determinant (Fuc-α1,2-Gal) [
20,
21], not considered in our study.
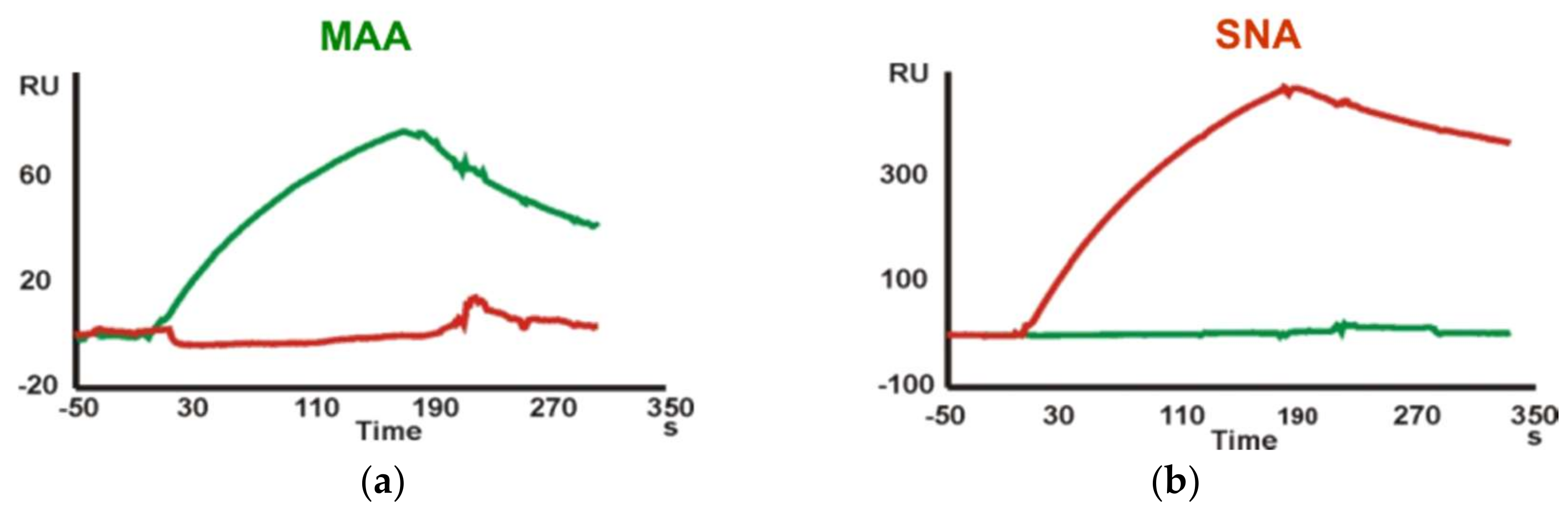
Finally, kinetic data for the interaction of Neu5Ac-containing trisaccharide glycoprobes with the
Maackia amurensis (MAA) and
Sambucus nigra (SNA) lectins were determined. A sensor chip with the Neu5Ac-α2,3-Gal-β1,4-GlcNAc-
N[Me]-
O-Aoa-GFKKG-amide, Neu5Ac-α2,6-Gal-β1,4-GlcNAc-
N[Me]-
O-Aoa-GFKKG-amide and Neu5Ac-α2,3/6-Gal-β1,4-Glc-
N[Me]-
O-Aoa-GFKKG-amide glycoprobes, representative of the two existing linkage types in adult mammalian glycoproteins, was prepared and the two Sia-specific lectins were flown over the surfaces. As the MAA lectin reportedly requires three intact sugar units for binding [
22], our results support a native ring-closed structure for the first monosaccharide. The binding responses observed for the two lectins were in perfect agreement with their reported carbohydrate specificity. Thus, whereas SNA showed a marked preference for the Sia-α2,6-lacNAc isomer, MAA recognized only the Sia-α2,3-lacNAc containing glycoprobe (
Figure 3).
For kinetic analysis, several concentrations in the 250 nM to 1.9 μM range for MAA and in the 74 to 563 nM range for SNA were analyzed. A 1:1 Langmuir binding model was used for sensorgram fitting (
Figure 4,
Table 3). As expected for interactions involving trisaccharides, the K
A equilibrium constants obtained were in the 10
6–10
7 M
−1 range. For MAA, the affinity constant determined by our approach was 9.12 × 10
5 M
−1; of the same magnitude than reported previously [
23], but one order lower than other constants determined using complex neoglycoproteins and immobilized lectins [
24], in both cases by SPR. The higher affinity for neoglycoproteins can easily be explained by their multivalent nature. For SNA, a K
A of 6.27 × 10
6 M
−1 was determined, consistent with the previously reported value [
25] of 6.7 × 10
6 M
−1.
In conclusion, the kinetic and affinity constants of well-known plant lectins for their specific carbohydrate ligands determined by our SPR approach were consistent with previously reported data, corroborating that the glycoprobes attached to sensor surface chips provide a reliable setting for accurate determination of the carbohydrate binding profile of the mammalian lectin PDC-109.
2.1.3. PDC-109 Binding Profile and Kinetic Studies
While the lipid selectivity and kinetics of PDC-109 has been characterized in considerable detail [
11,
26], its interaction with carbohydrate epitopes remains largely uncharted, despite its potential role in sperm reservoir formation in the female genital tract. We have therefore performed SPR experiments with PDC-109 and the most relevant mammalian glycotopes, particularly those containing Fuc epitopes. Once SPR sensor chips were functionalized with the different synthetic glycoprobes and evaluated with specific lectins (
Section 2.1.2) they were used to characterize both qualitatively and quantitatively the interaction of PDC-109. For kinetic analysis, several concentrations in the 6.25 to 100 μM range were analyzed and a 1:1 Langmuir binding model was chosen for sensorgram fitting.
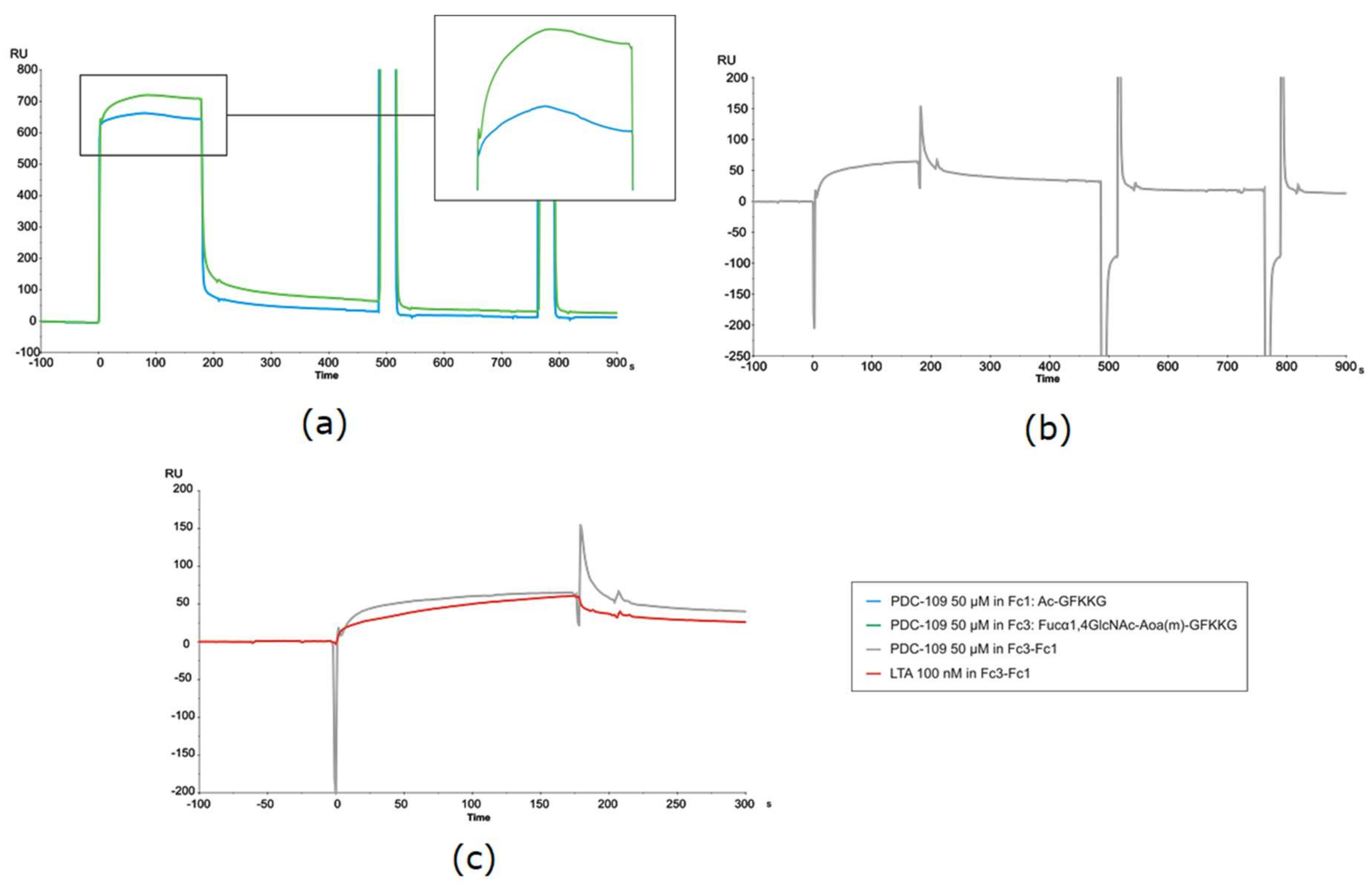
In general, considerable variability was observed in the kinetic experiments, with CV (from 3 to 6 repeat experiments) exceeding 50% in particular cases. Data dispersion was attributable to the unusual sensorgrams observed, reflecting a complex binding event, particularly in the association phase of PDC-109. As shown in
Figure 5a, the usual signal rise observed right after sample injection and due to bulk effects—refractive index differences between analyte solution and flow buffer—was followed by a rather intriguing association phase. In a conventional, straightforward binding event, the response—i.e., the signal rise over time, reflecting protein binding to the sensor surface—reaches a constant value once equilibrium is established, and remains so until the protein solution passed over the sensor chip is replaced with buffer, giving way to the spontaneous dissociation process and subsequent surface regeneration. However, in the present case visual inspection of all PDC-109 sensorgrams (those in
Figure 5a being representative) clearly shows a biphasic phenomenon during the association phase (displayed enlarged in
Figure 5a).
Thus, response increased until a high value (maximum binding) was reached, then slowly decreased even though PDC-109 was still being flown across the surface. Such biphasic behavior during the association phase was observed not only with all immobilized glycoprobes but also with the reference surface (
Figure 5a; green and blue trace sensorgrams, respectively), hence the differential curve resulting from reference subtraction (
Figure 5b) displayed an uneven shape, with large spikes at both the beginning and the end of the injection that complicated data fitting. As a result, values for the kinetic constants may include some variability, although a reliable epitope preference could nonetheless be established.
This unusual biphasic profile, reflecting loss of mass from the sensor surface during the association phase, was not observed with any of the standard lectins studied earlier (
Figure 5c), and suggested an uncharacteristic binding behavior of PDC-109 that could be confirmed after meticulous evaluation of all possible variables (surface, ligand, analyte, buffer conditions, competitive reagents, solute stability, etc.) and with some precedents observed in other systems [
26].
Plausible explanations for the unusual behavior of PDC-109, accounting for the biphasic shape of the SPR sensorgrams, would assume the existence of two different affinities, namely two competing analyte reactions, in the protein preparation. This can be due either to a single analyte with two different affinities, or to two different analytes, each with its own affinity.
Regarding the former situation, the possibility of non-specific binding between PDC-109 and the SPR surface was investigated, since the carboxymethyled dextran matrix on the sensor surface is structurally not too distant from the glycoprobe ligands, hence a weak interaction with the lectin could not be excluded. To this end, a “saturated” solution of free carboxymethylated dextran (CM) (identical to that present as CM5 surface coating) was added to the analyte solution; if the analyte had any affinity for either dextran or carboxymethylated variant, it would bind preferentially the free polymer over that on the sensor chip, thus reducing non-specific binding and avoiding competing reactions. This modification, however, did not have any effect on the binding curves, which remained biphasic in all cases. In a complementary attempt, a C1 chip lacking the dextran matrix (only carboxymethyl groups directly bound to the gold surface) was tested to exclude any influence of the matrix; again, the same biphasic profile could be observed during the association phase. From these results, we concluded that no competing reactions with the SPR surface existed, hence the biphasic behavior must proceed from analyte heterogeneity, i.e., two analytes competing for the same ligand. If such was the case, the decreasing region in the association phase curve could result from a smaller molecule with higher ligand affinity but slower kinetics competing with a faster but lower-affinity interaction.
Different possibilities of analyte heterogeneity can be envisaged: (i) PDC-109 may undergo conformational changes upon binding to the epitope; (ii) more than one isoform/glycoform is present in the PDC-109 sample; or (iii) PDC-109 has different aggregation states; all these situations might produce sensorgrams with biphasic association curves, such as those in
Figure 6. Previous reports suggest PDC-109 undergoes a conformational change upon binding to PC membranes [
27]. However, as SPR can only detect refractive index variations related to mass changes over time, subtle conformational changes not resulting in mass changes will go unnoticed. However, while beyond the scope of the SPR technique, heterogeneity due to conformational change would not explain the SPR signal reduction.
To investigate the possibility of a heterogeneous analyte, several different approaches were followed. Firstly, considering that PDC-109 exists in two major forms (i.e., glycosylated and non-glycosylated) [
10], enzymatic deglycosylation was performed in order to obtain a single analyte isoform. Given the limited availability of purified PDC-109, the amount of protein used for deglycosylation did not allow a quantitative purification step. The extent of the reaction was assessed by matrix assisted laser desorption ionization—time of flight (MALDI-TOF) MS, which showed that glycan removal could not be brought to completion. Even so, the partially deglycosylated PDC-109 was tested on the same sensor surfaces previously used with native protein. The resulting sensorgrams clearly showed a rising association phase without biphasic behavior, yet the incomplete deglycosylation prevented formulating clear conclusions, as the standard binding response could not be unequivocally attributed to a single isoform. Secondly, to investigate the possibility of aggregation, PDC-109 was flown across a chip where itself was immobilized. The resulting sensorgrams displayed the previously observed biphasic shape, suggesting that interaction of PDC-109 with itself is the cause of the anomalous biphasic sensorgrams, and that the glycosylated version plays a significant role in this self-recognition, possibly by virtue of the single sialic acid residue present.
Evidence from the literature points out that PDC-109 is naturally produced as a mixture of several protein forms [
28] and aggregation states [
29] that play an important role in modulating its interaction with other biomolecules. This would allow concluding that, whichever of the situations proposed in
Figure 6 applies –including a combination thereof–, PDC-109 acts as a lectin-like molecule albeit with quite unique features that, though not abolishing its ability to recognize carbohydrates, give rise to unusual SPR sensorgrams.
Despite the above-discussed limitations, we were able to determine the SPR binding parameters of PDC-109. Affinity constants (
Table 4) indicated weak to moderate binding for carbohydrates, at least if compared to the considerably higher affinity (K
A~10
7 M
−1) found for PC membranes [
26]. This would suggest that PDC-109 binding to carbohydrates in oviductal epithelial cells must likely involve some sort of multivalency in order to efficiently retain the sperm in the oviductal reservoir.
From the above data it can also be seen that PDC-109 appears to have the highest binding affinity for Fuc-α1,4-GlcNAc among all glycotopes in this study. This is in agreement with earlier findings which propose that bull sperm binds to an oligosaccharide ligand on the oviductal epithelium that resembles Lewis
a trisaccharide (Galβ1-3[Fucα1-4]GlcNAc) [
30]. For PDC-109 vs. Fuc-α1,4-GlcNAc, the higher affinity was a consequence of a lower dissociation rate constant, consistent with a privileged interaction with this glycosidic configuration. The observation that PDC-109 displays favored interaction with the Neu5Ac-α2,6-Gal-β1,4-GlcNAc epitope may account for the observed self-recognition. Part of PDC-109 bears an O-linked trisaccharide consistent with this mass [
10] and, even though this aspect has never been fully elucidated, it could well be that the Sia residue, when exposed in an α-2,6-configuration, facilitates such binding. Finally, the absence of significant binding with the mannobiose epitopes is in contrast to what was reported by Amari and coworkers. However, they employed oligomannose type pentassacharides and the complex architecture of this oligosaccharide may well account for the difference in binding characteristics.










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}