An Organic Solvent-Tolerant Lipase with Both Hydrolytic and Synthetic Activities from the Oleaginous Fungus Mortierella echinosphaera


 ,
,  and
and 
Abstract
:1. Introduction
2. Results and Discussion
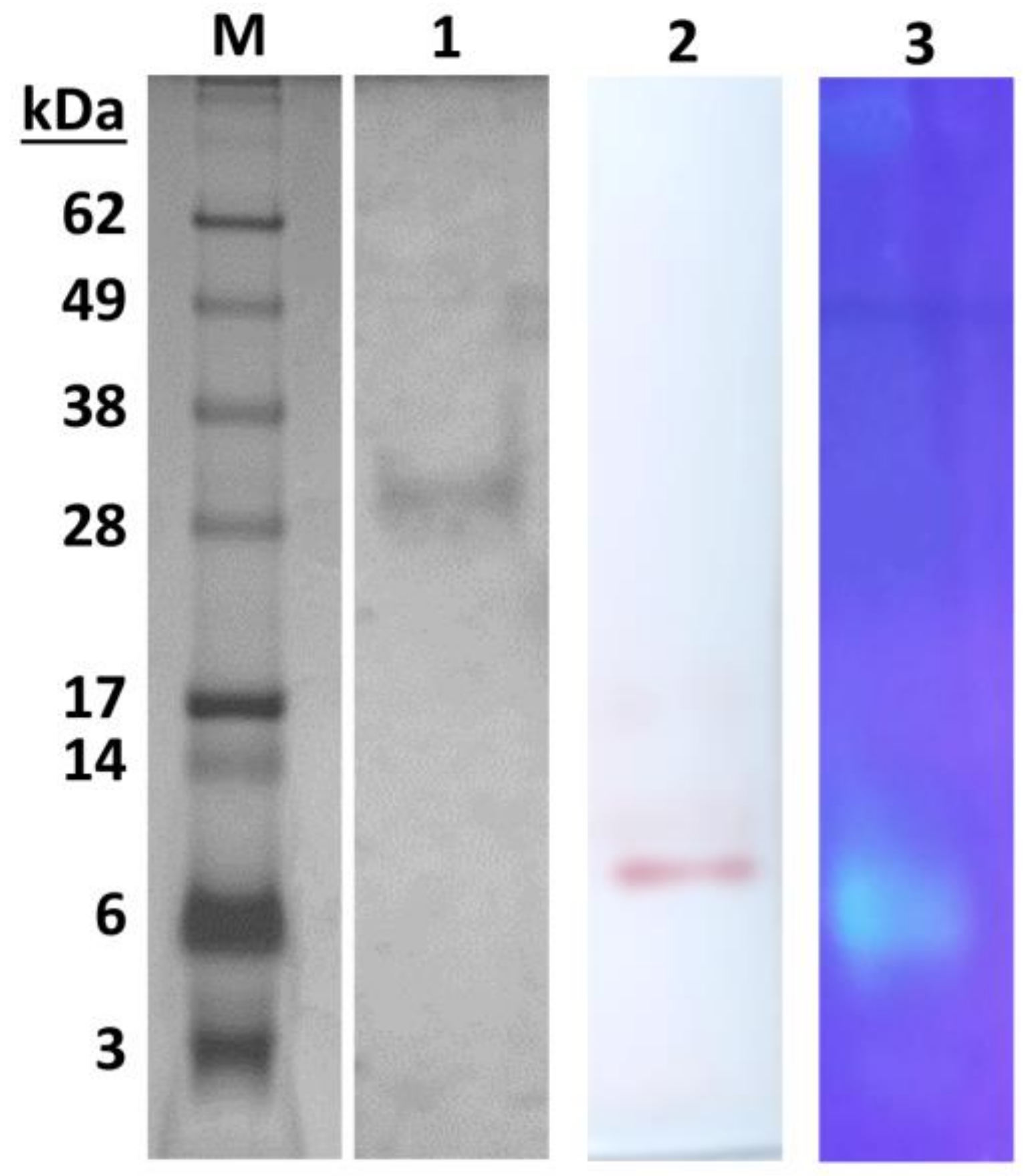
2.1. Purification of Extracellular Lipolytic Enzyme
2.2. Characterization of the Purified Lipase
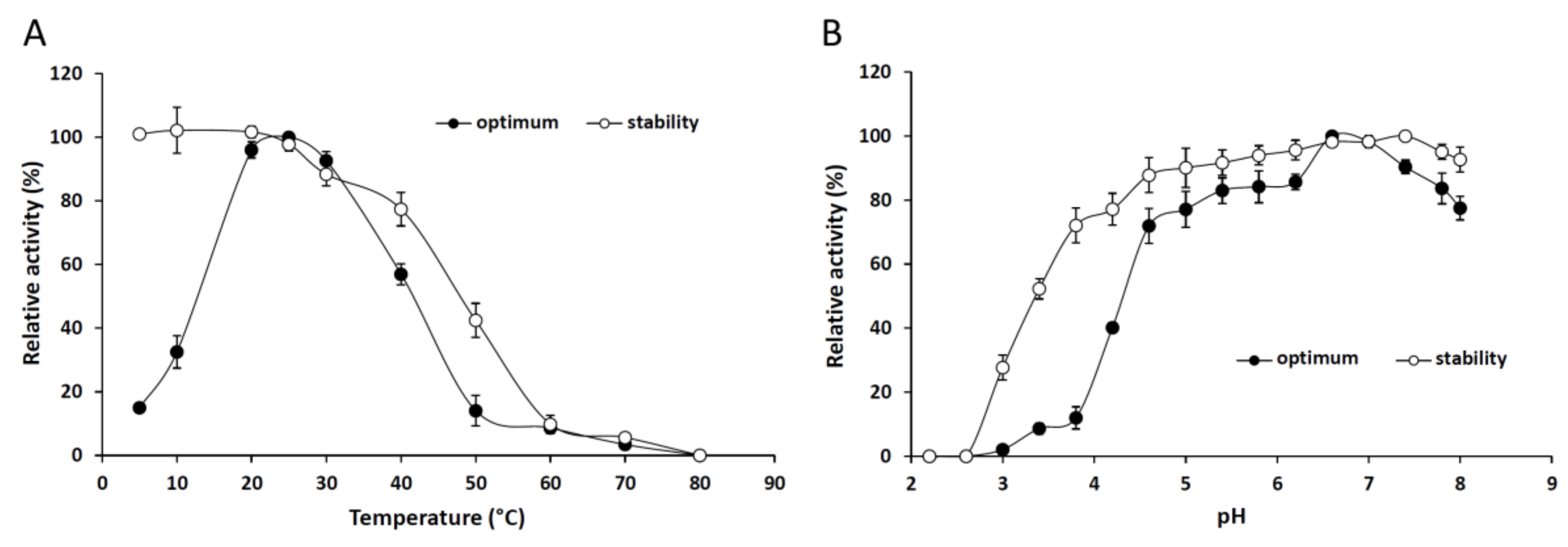
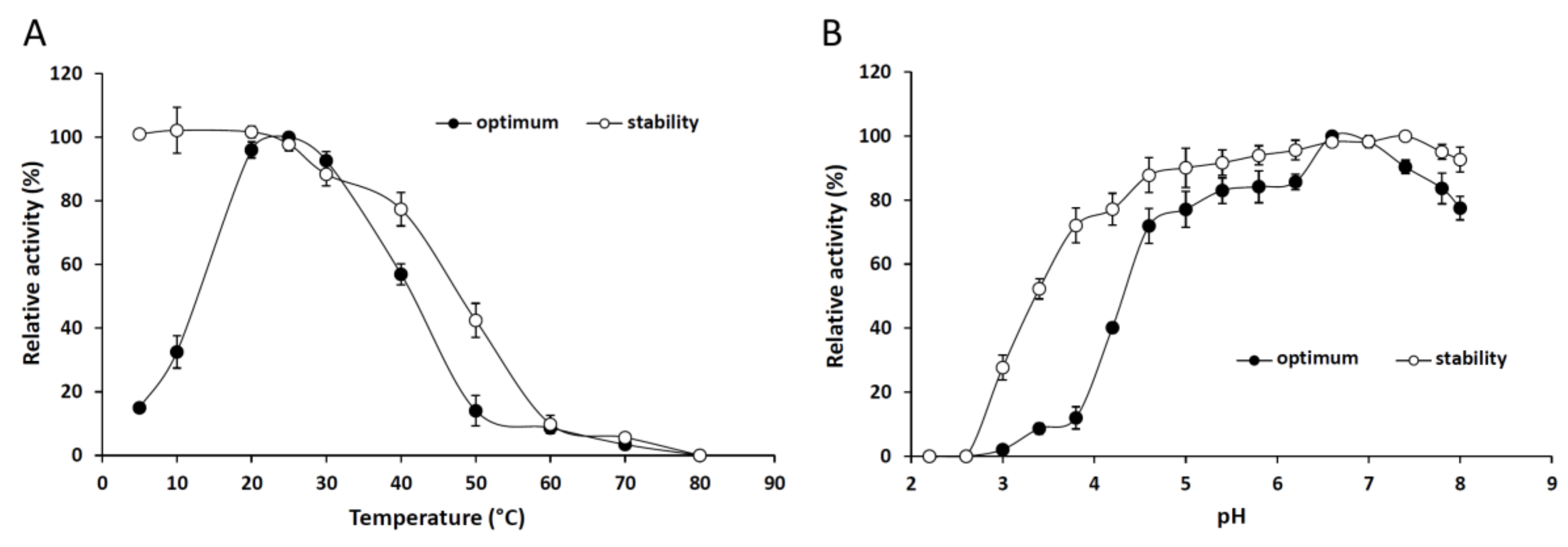
2.2.1. Effect of the Temperature and pH on the Activity and Stability of the Purified Lipase
2.2.2. Determination of Kinetic Constants
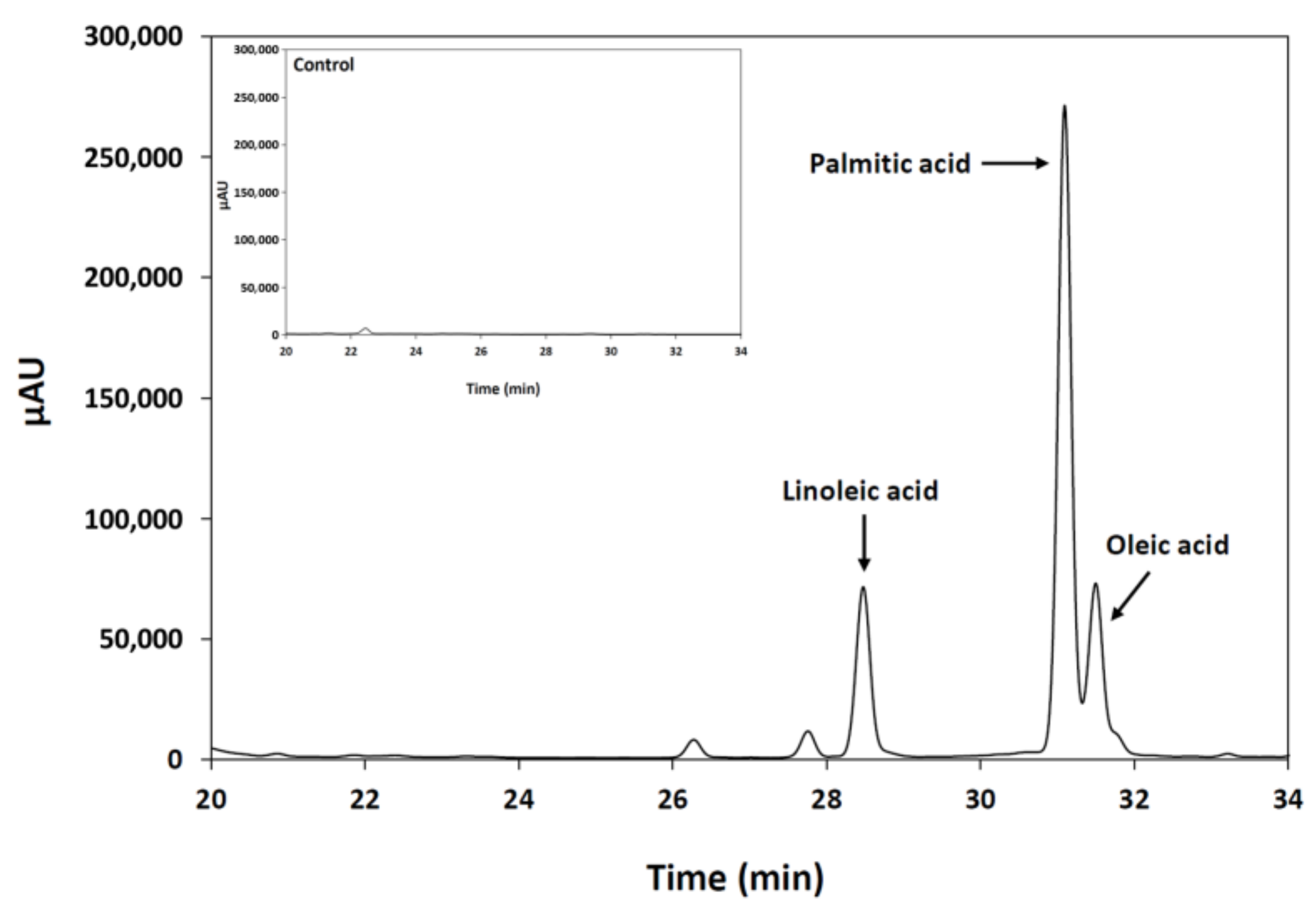
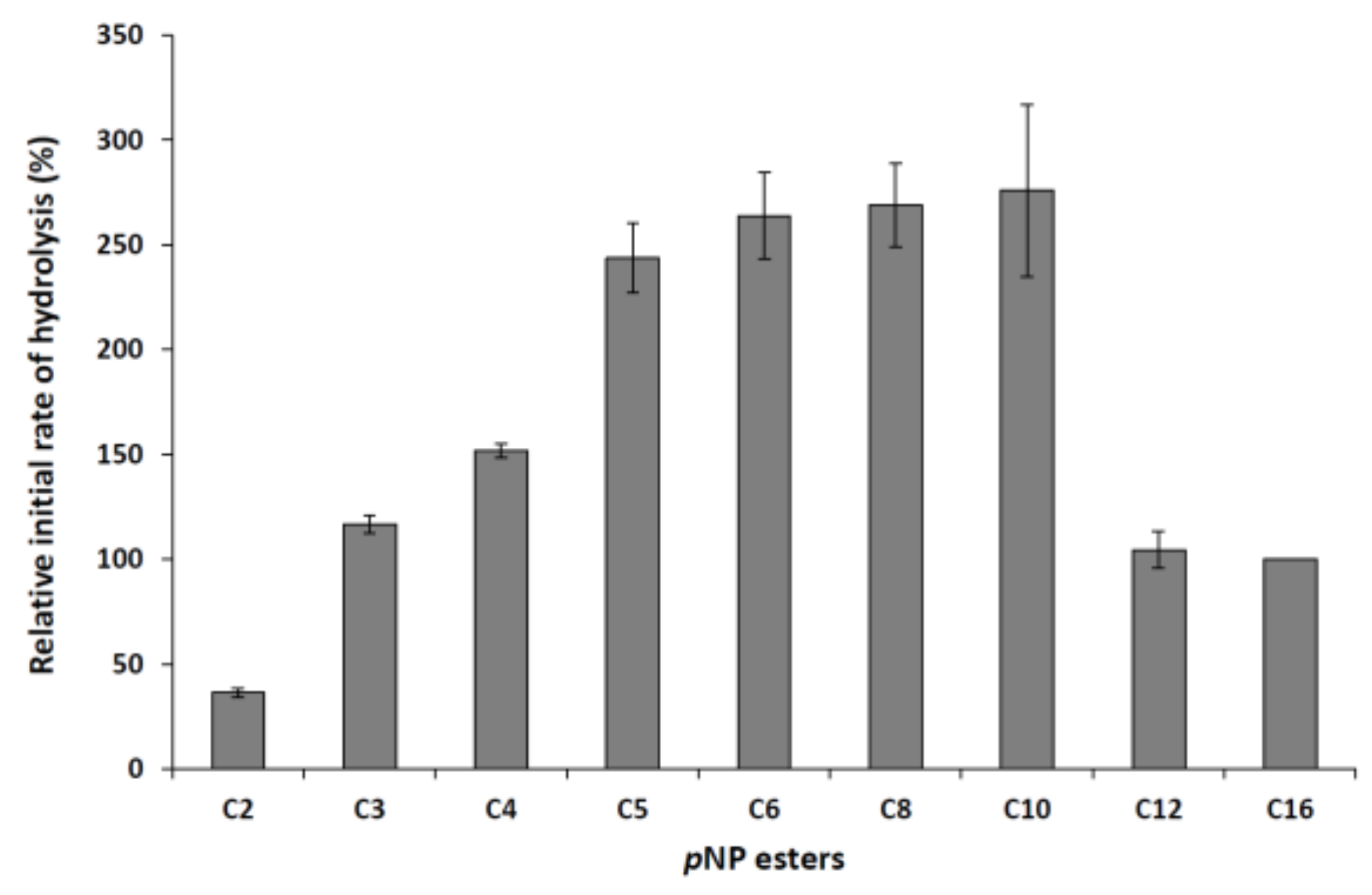
2.2.3. Determination of the Substrate Specificity
2.2.4. Positional Specificity
2.2.5. Effect of Metal Ions and Reagents
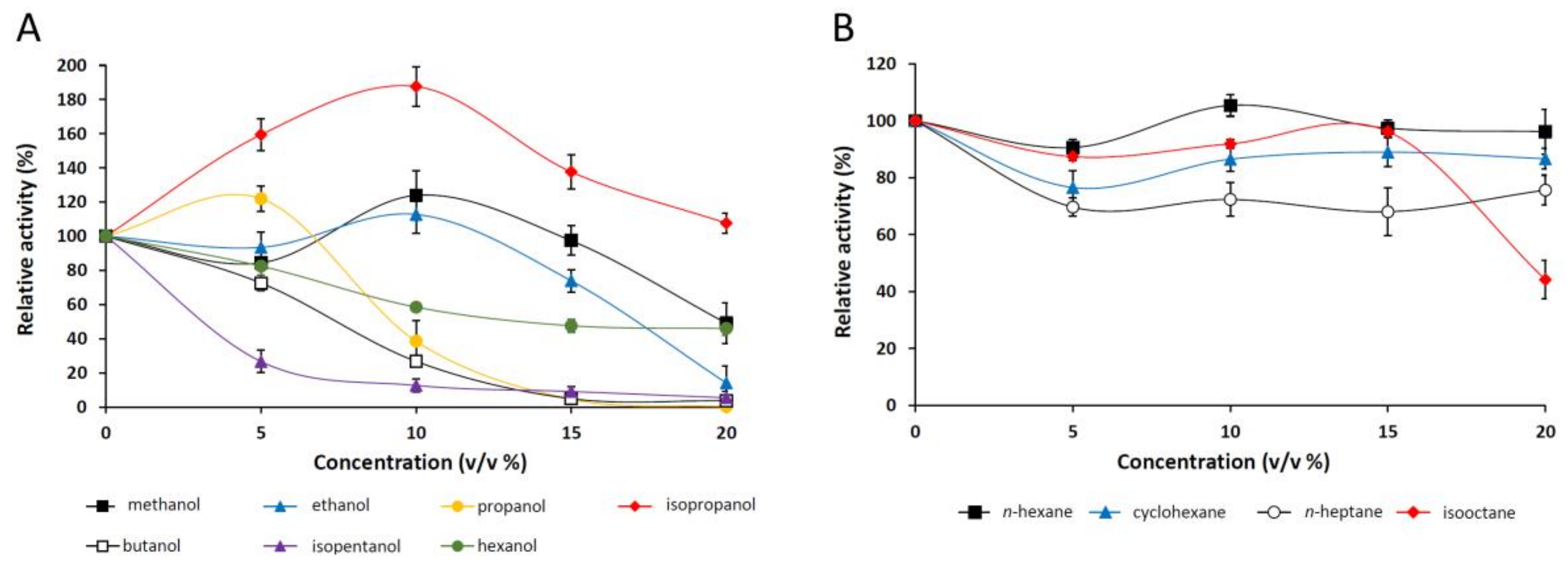
2.2.6. Effect of Organic Solvents on the Purified Lipase
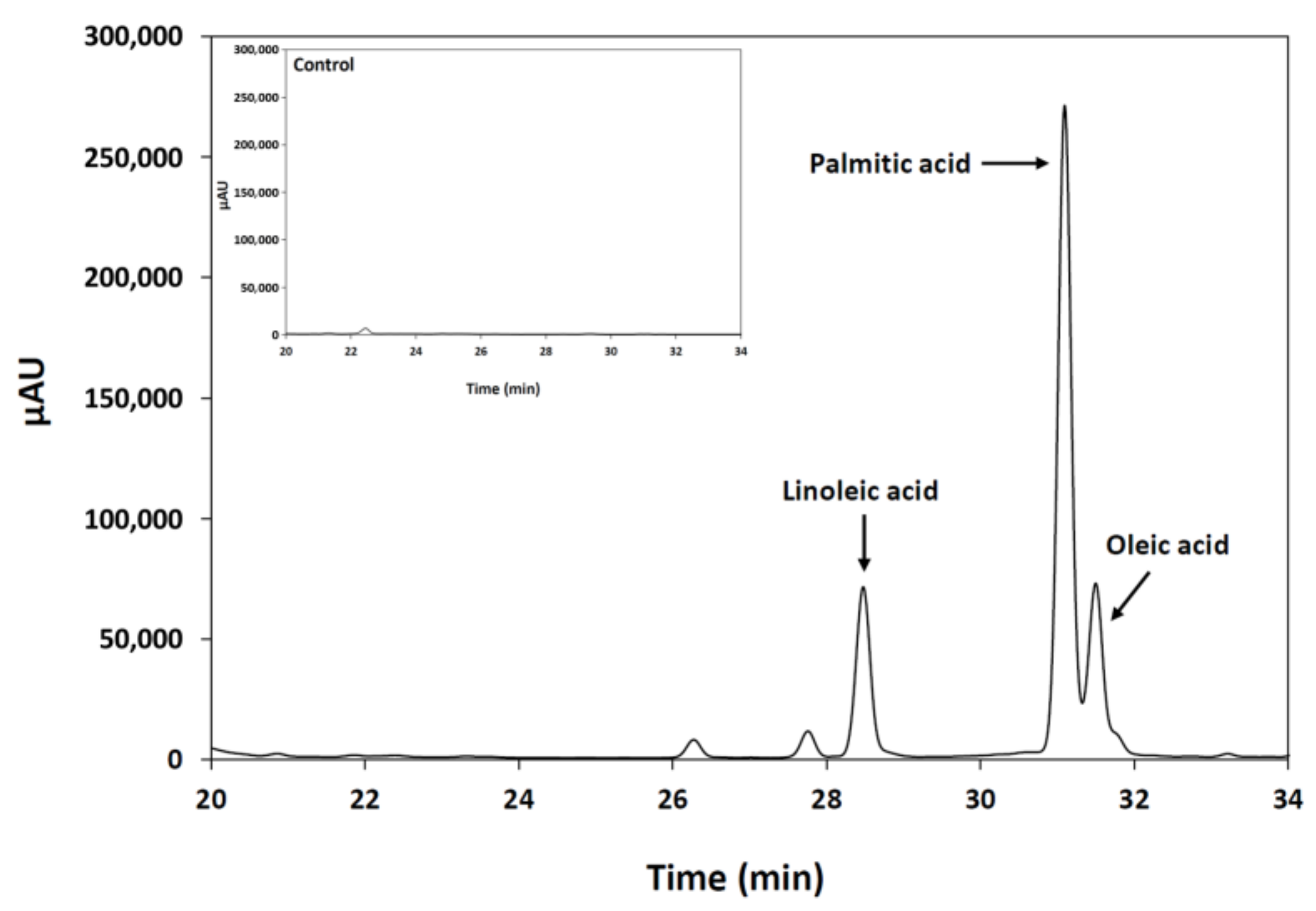
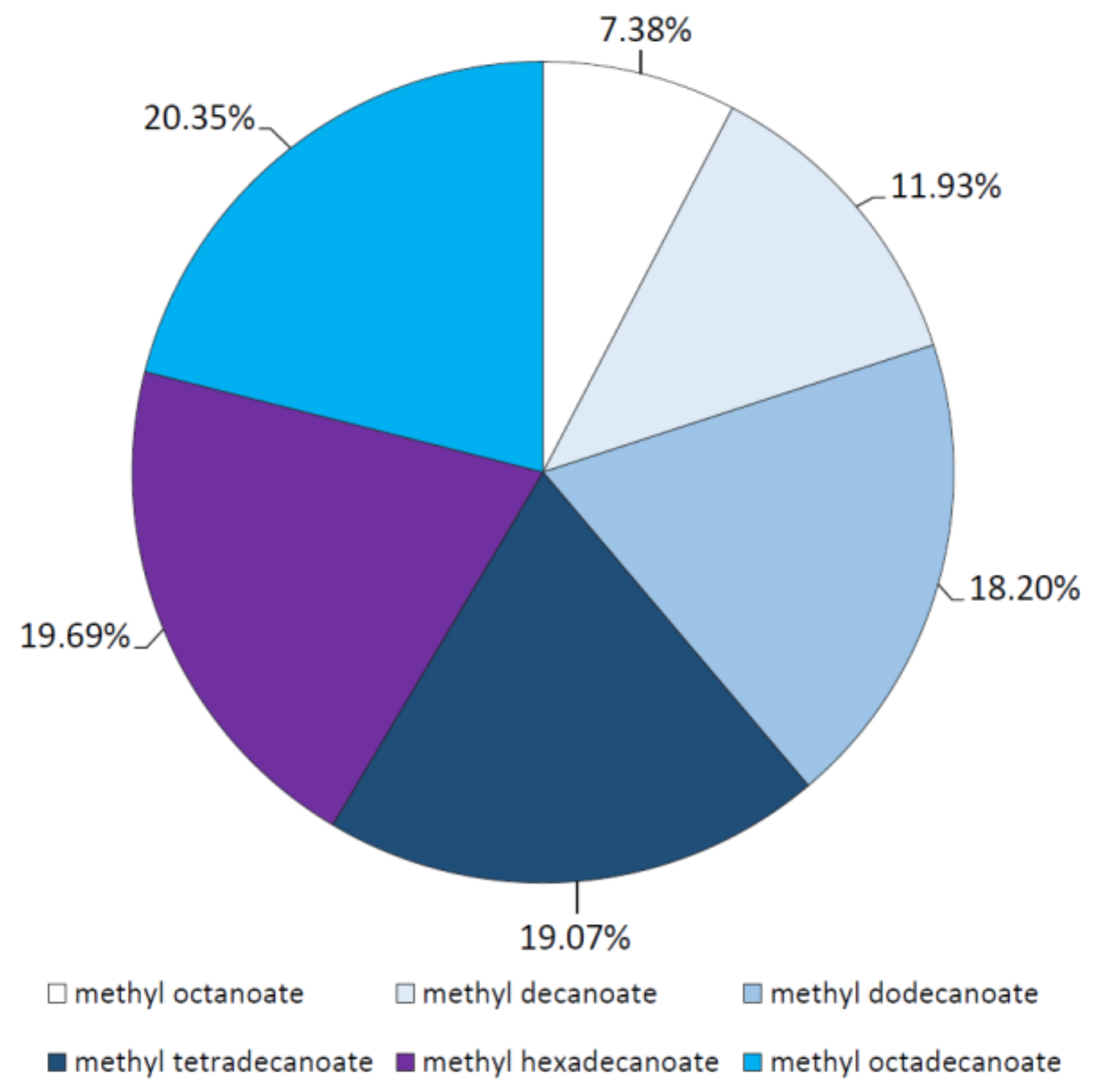
2.3. Analysis of Esterification Reactions Catalyzed by Purified Lipase
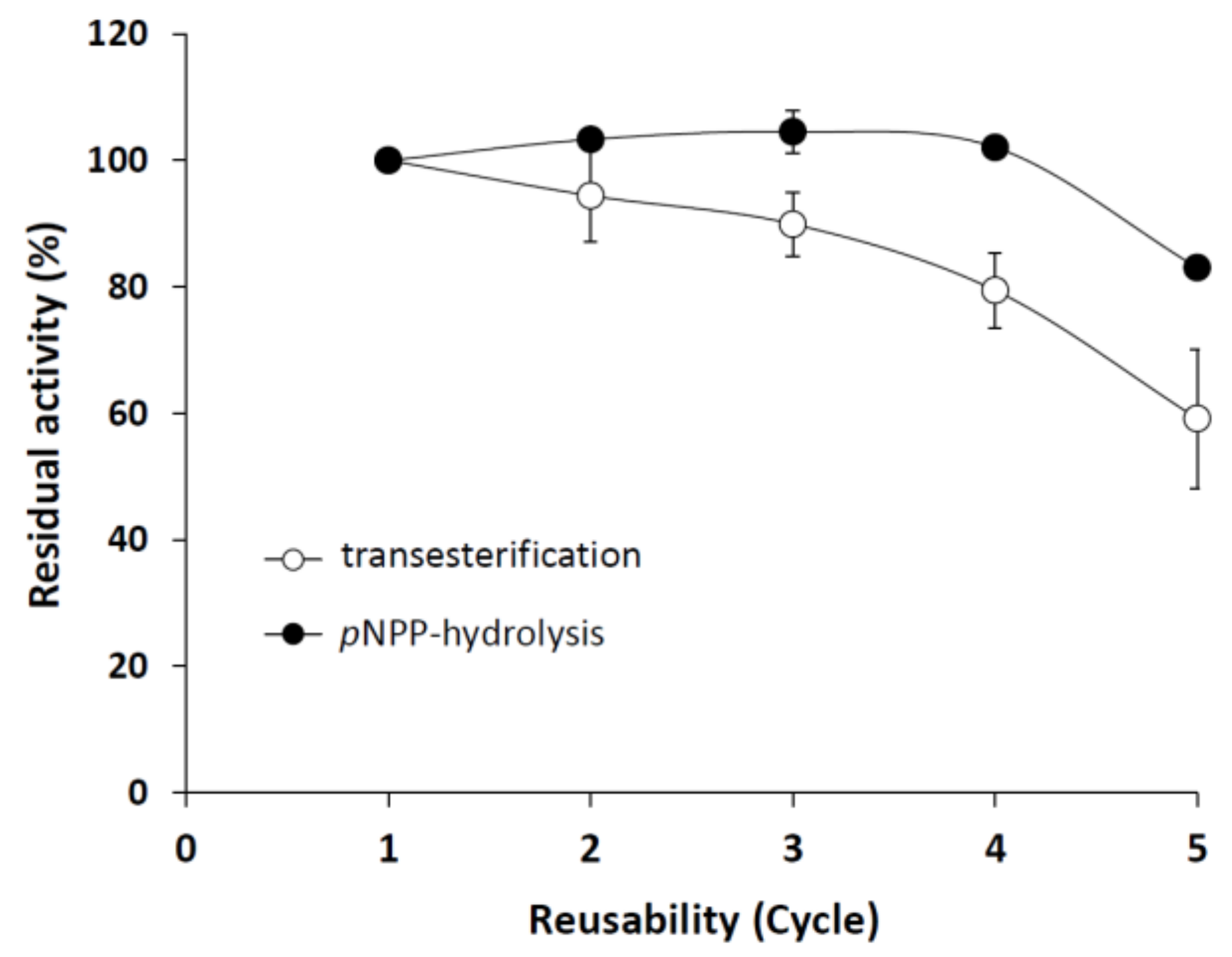
2.4. Immobilization of the M. echinosphaera Lipase
3. Materials and Methods
3.1. Microorganism and Submerged Fermentation
3.2. Detection of the Lipase Activities
3.3. Purification of Extracellular Lipase
3.4. Protein Concentration Assay
3.5. Gel Electrophoresis and Zymography
3.6. Isoelectric Focusing
3.7. Characterization of the Purified Lipolytic Enzyme
3.7.1. Effect of Temperature and pH
3.7.2. Determination of Kinetic Parameters
3.7.3. Determination of the Substrate Specificity
3.7.4. Positional Specificity Assays
3.7.5. Effect of Metal Ions and Reagents
3.7.6. Effect of Organic Solvents on the Purified Lipase
3.8. Esterification Activity Studies
3.9. Immobilization of the Lipase
4. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Li, S.; Yang, X.; Yang, S.; Zhu, M.; Wang, X. Technology prospecting on enzymes: Application, marketing and engineering. Comput. Struct. Biotechnol. J. 2012, 2, e201209017. [Google Scholar] [CrossRef] [PubMed]
- Aravindan, R.; Anbumathi, P.; Viruthagiri, T. Lipase applications in food industry. Indian J. Biotechnol. 2007, 6, 141–158. [Google Scholar]
- Sharma, S.; Kanwar, S.S. Organic solvent tolerant lipases and applications. Sci. World J. 2014, 2014, 625258. [Google Scholar] [CrossRef] [PubMed]
- Andualema, B.; Gessesse, A. Microbial lipases and their industrial applications: Review. Biotechnology 2012, 11, 100–118. [Google Scholar] [CrossRef]
- Satari, B.; Karimi, K. Mucoralean fungi for sustainable production of bioethanol and biologically active molecules. Appl. Microbiol. Biotechnol. 2018, 102, 1097–1117. [Google Scholar] [CrossRef] [PubMed]
- Dyal, S.D.; Narine, S.S. Implications for the use of Mortierella fungi in the industrial production of essential fatty acids. Food Res. Int. 2005, 38, 445–467. [Google Scholar] [CrossRef]
- Certik, M.; Shimizu, S. Biosynthesis and regulation of microbial polyunsaturated fatty acid production. J. Biosci. Bioeng. 1999, 87, 1–14. [Google Scholar] [CrossRef]
- Szczęsna-Antczak, M.; Antczak, T.; Piotrowicz-Wasiak, M.; Rzyska, M.; Binkowska, N.; Bielecki, S. Relationships between lipases and lipids in mycelia of two Mucor strains. Enzyme Microb. Technol. 2006, 39, 1214–1222. [Google Scholar] [CrossRef]
- Singh, A.K.; Mukhopadhyay, M. Overview of fungal lipase: A review. Appl. Biochem. Biotechnol. 2012, 166, 486–520. [Google Scholar] [CrossRef] [PubMed]
- Kotogán, A.; Kecskeméti, A.; Szekeres, A.; Papp, T.; Chandrasekaran, M.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, Cs.; Takó, M. Characterization of transesterification reactions by Mucoromycotina lipases in non-aqueous media. J. Mol. Catal. B Enzym. 2016, 127, 47–55. [Google Scholar] [CrossRef]
- Gaspar, M.L.; Cunningham, M.; Pollero, R.; Cabello, M. Occurrence and properties of an extracellular lipase in Mortierella vinacea. Mycologia 1999, 91, 108–113. [Google Scholar] [CrossRef]
- Jermsuntiea, W.; Aki, T.; Toyoura, R.; Iwashita, K.; Kawamoto, S.; Ono, K. Purification and characterization of intracellular lipase from the polyunsaturated fatty acid-producing fungus Mortierella alliacea. New Biotechnol. 2011, 28, 158–164. [Google Scholar] [CrossRef] [PubMed]
- Kotogán, A.; Németh, B.; Vágvölgyi, Cs.; Papp, T.; Takó, M. Screening for extracellular lipase enzymes with transesterification capacity in Mucoromycotina strains. Food Technol. Biotechnol. 2014, 52, 73–82. [Google Scholar]
- Speranza, P.; Macedo, G.A. Lipase-mediated production of specific lipids with improved biological and physicochemical properties. Process Biochem. 2012, 47, 1699–1706. [Google Scholar] [CrossRef]
- Sá, A.G.A.; de Meneses, A.C.; de Araújo, P.H.H.; de Oliveira, D. A review on enzymatic synthesis of aromatic esters used as flavor ingredients for food, cosmetics and pharmaceuticals industries. Trends Food Sci. Technol. 2017, 69, 95–105. [Google Scholar] [CrossRef]
- Manoel, E.A.; Ribeiro, M.F.P.; dos Santos, J.C.S.; Coelho, M.A.Z.; Simas, A.B.C.; Fernandez-Lafuente, R.; Freire, D.M.G. Accurel MP 1000 as a support for the immobilization of lipase from Burkholderia cepacia: Application to the kinetic resolution of myo-inositol derivatives. Process Biochem. 2015, 50, 1557–1564. [Google Scholar] [CrossRef]
- Patel, A.K.; Singhania, R.R; Pandey, A. Production, purification, and application of microbial enzymes. In Biotechnology of Microbial Enzymes. Production, Biocatalysis and Industrial Applications; Brahmachari, G., Demain, A.L., Adrio, J.L., Eds.; Academic Press: Oxford, UK, 2017; pp. 13–41. [Google Scholar]
- Saxena, R.K.; Sheoran, A.; Giri, B.; Davidson, W.S. Purification strategies for microbial lipases. J. Microbiol. Meth. 2003, 52, 1–18. [Google Scholar] [CrossRef]
- Kotogán, A.; Papp, T.; Vágvölgyi, Cs.; Takó, M. Extracellular lipase production in solid state fermentation using agricultural and food industrial by-products as substrates. In Proceedings of the Food Science Conference 2013—With Research for the Success of Darányi Program, Budapest, Hungary, 7–8 November 2013; Faculty of Food Science, Corvinus University of Budapest: Budapest, Hungary, 2013; pp. 173–176. [Google Scholar]
- Abbas, H.; Hiol, A.; Deyris, V.; Comeau, L. Isolation and characterization of an extracellular lipase from Mucor sp. strain isolated from palm fruit. Enzyme Microb. Technol. 2002, 31, 968–975. [Google Scholar] [CrossRef]
- Mateos Diaz, J.C.; Rodríguez, J.A.; Roussos, S.; Cordova, J.; Abousalham, A.; Carriere, F.; Baratti, J. Lipase from the thermotolerant fungus Rhizopus homothallicus is more thermostable when produced using solid state fermentation than liquid fermentation procedures. Enzyme Microb. Technol. 2006, 39, 1042–1050. [Google Scholar] [CrossRef]
- Dobrev, G.; Zhekova, B.; Nedelcheva, P.; Chochkov, R.; Krastanov, A. Characterization of crude lipase from Rhizopus arrhizus and purification of multiplicity forms of the enzyme. Biotechnol. Biotechnol. Equip. 2011, 25, 2295–2300. [Google Scholar] [CrossRef]
- Takó, M.; Kotogán, A.; Papp, T.; Kadaikunnan, S.; Alharbi, N.S.; Vágvölgyi, Cs. Purification and properties of extracellular lipases with transesterification activity and 1,3-regioselectivity from Rhizomucor miehei and Rhizopus oryzae. J. Microbiol. Biotechnol. 2017, 27, 277–288. [Google Scholar] [CrossRef] [PubMed]
- Joseph, B.; Ramteke, P.W.; Thomas, G.; Shrivastava, N. Standard review cold-active microbial lipases: A versatile tool for industrial applications. Biotechnol. Mol. Biol. Rev. 2007, 2, 39–48. [Google Scholar]
- Yu, X.W.; Wang, L.L.; Xu, Y. Rhizopus chinensis lipase: Gene cloning, expression in Pichia pastoris and properties. J. Mol. Catal. B Enzym. 2009, 57, 304–311. [Google Scholar] [CrossRef]
- Sun, S.Y.; Xu, Y. Solid-state fermentation for ‘whole-cell synthetic lipase’ production from Rhizopus chinensis and identification of the functional enzyme. Process Biochem. 2008, 43, 219–224. [Google Scholar] [CrossRef]
- Garcia-Galan, C.; Barbosa, O.; Ortiz, C.; Torres, R.; Rodrigues, R.C.; Fernandez-Lafuente, R. Biotechnological prospects of the lipase from Mucor javanicus. J. Mol. Catal. B Enzym. 2013, 93, 34–43. [Google Scholar] [CrossRef]
- Saxena, R.K.; Agarwal, L.; Meghwanshi, G.K. Diversity of fungal and yeast lipases: Present and future scenario for the 21st century. In Microbial Diversity: Current Perspectives and Potential Applications; Satyanarayana, T., Johri, B.N., Eds.; I.K. International Publishing House Pvt. Ltd.: New Delhi, India, 2005; pp. 791–814. [Google Scholar]
- Norjannah, B.; Ong, H.C.; Masjuki, H.H.; Juan, J.C.; Chong, W.T. Enzymatic transesterification for biodiesel production: A comprehensive review. RSC Adv. 2016, 6, 60034–60055. [Google Scholar] [CrossRef]
- Fregolente, P.B.L.; Fregolente, L.V.; Maciel, M.R.W.; Carvalho, P.O. Screening of microbial lipases and evaluation of their potential to produce glycerides with high gamma linolenic acid concentration. Braz. J. Microbiol. 2009, 40, 747–756. [Google Scholar] [CrossRef] [PubMed]
- Haas, M.J.; Cichowicz, D.J.; Bailey, D.G. Purification and characterization of an extracellular lipase from the fungus Rhizopus delemar. Lipids 1992, 27, 571–576. [Google Scholar] [CrossRef]
- Hiol, A.; Jonzo, M.D.; Druet, D.; Comeau, L.C. Production, purification and characterization of an extracellular lipase from Mucor hiemalis f. hiemalis. Enzyme. Microb. Technol. 1999, 25, 80–87. [Google Scholar] [CrossRef]
- Çolak, A.; Şişik, D.; Saglam, N.; Güner, S.; Çanakçi, S.; Beldüz, A.O. Characterization of a thermoalkalophilic esterase from a novel thermophilic bacterium, Anoxybacillus gonensis G2. Bioresour. Technol. 2005, 96, 625–631. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Li, X.; Wang, Y.; Wang, Y.; Wang, F.; Jiang, J. Expression and characterization of recombinant Rhizopus oryzae lipase for enzymatic biodiesel production. Bioresour. Technol. 2011, 102, 9810–9813. [Google Scholar] [CrossRef] [PubMed]
- Salameh, M.A.; Wiegel, J. Purification and characterization of two highly thermophilic alkaline lipases from Thermosyntropha lipolytica. Appl. Environ. Microbiol. 2007, 73, 7725–7731. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.R.; Kwon, D.Y.; Yoon, S.H.; Kim, W.Y.; Kim, K.H. Purification, refolding, and characterization of recombinant Pseudomonas fluorescens lipase. Protein Expr. Purif. 2005, 39, 124–129. [Google Scholar] [CrossRef] [PubMed]
- Ebrahimpour, A.; Rahman, R.N.Z.R.A.; Basri, M.; Salleh, A.B. High level expression and characterization of a novel thermostable, organic solvent tolerant, 1,3-regioselective lipase from Geobacillus sp. strain ARM. Bioresour. Technol. 2011, 102, 6972–6981. [Google Scholar] [CrossRef] [PubMed]
- Atadashi, I.M.; Aroua, M.K.; Abdul Aziz, A.R.; Sulaiman, N.M.N. The effects of water on biodiesel production and refining technologies: A review. Renew. Sustain. Energy Rev. 2012, 16, 3456–3470. [Google Scholar] [CrossRef]
- Verma, M.L.; Azmi, W.; Kanwar, S.S. Microbial lipases: At the interface of aqueous and non-aqueous media. A review. Acta Microbiol. Immunol. Hung. 2008, 55, 265–294. [Google Scholar] [CrossRef] [PubMed]
- Silva, J.E.S.; Jesus, P.C. Evaluation of the catalytic activity of lipases immobilized on chrysotile for esterification. An. Acad. Bras. Ciênc. 2003, 75, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y.; Xu, Y. Membrane-bound ‘synthetic lipase’ specifically cultured under solid-state fermentation and submerged fermentation by Rhizopus chinensis: A comparative investigation. Bioresour. Technol. 2009, 100, 1336–1342. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.Y.; Xu, Y.; Wang, D. Novel minor lipase from Rhizopus chinensis during solid-state fermentation: Biochemical characterization and its esterification potential for ester synthesis. Bioresour. Technol. 2009, 100, 2607–2612. [Google Scholar] [CrossRef] [PubMed]
- Hari Krishna, S.; Divakar, S.; Prapulla, S.G.; Karanth, N.G. Enzymatic synthesis of isoamyl acetate using immobilized lipase from Rhizomucor miehei. J. Biotechnol. 2001, 87, 193–201. [Google Scholar] [CrossRef]
- Abbas, H.; Comeau, L. Aroma synthesis by immobilized lipase from Mucor sp. Enzyme Microb. Technol. 2003, 32, 589–595. [Google Scholar] [CrossRef]
- Alnoch, R.C.; Martini, V.P.; Glogauer, A.; dos Santos Costa, A.C.; Piovan, L.; Muller-Santos, M.; de Souza, E.M.; de Oliveira Pedrosa, F.; Mitchell, D.A.; Krieger, N. Immobilization and characterization of a new regioselective and enantioselective lipase obtained from a metagenomic library. PLoS ONE 2015, 10, e0114945. [Google Scholar] [CrossRef] [PubMed]
- Madalozzo, A.D.; Martini, V.P.; Kuniyoshi, K.K.; de Souza, E.M.; Pedrosa, F.O.; Glogauer, A.; Zanin, G.M.; Mitchell, D.A.; Krieger, N. Immobilization of LipC12, a new lipase obtained by metagenomics, and its application in the synthesis of biodiesel esters. J. Mol. Catal. B Enzym. 2015, 116, 45–51. [Google Scholar] [CrossRef]
- Jesionowski, T.; Zdarta, J.; Krajewska, B. Enzyme immobilization by adsorption: A review. Adsorption 2014, 20, 801–821. [Google Scholar] [CrossRef]
- Hita, E.; Robles, A.; Camacho, B.; González, P.A.; Esteban, L.; Jiménez, M.J.; Muñío, M.M.; Molina, E. Production of structured triacylglycerols by acidolysis catalyzed by lipases immobilized in a packed bed reactor. Biochem. Eng. J. 2009, 46, 257–264. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Purification Step | Total Protein (mg) | Total Activity (μmol/min) | Specific Activity (U/mg) | Purification Fold | Recovery (%) |
|---|---|---|---|---|---|
| Culture filtrate | 2485 | 47,570 | 19.14 | 1 | 100 |
| (NH4)2SO4 precipitation (50–85%) | 265.6 | 14,548.8 | 54.77 | 2.86 | 30.6 |
| Sephadex G-75 | 12 | 2672 | 222.67 | 11.6 | 5.6 |
| Anion exchange (Macro-Prep High Q) | 1.1 | 291.6 | 264.13 | 13.79 | 0.61 |
| Sephacryl S200HR | 0.21 | 79.5 | 378.57 | 19.5 | 0.17 |
| Metal Ions and Reagents | Concentration (mM) | Residual Activity (% ± SD) 1 |
|---|---|---|
| HgCl2 | 5 | 42.3 ± 4.5 |
| CuSO4 | 5 | 104.2 ± 7.4 |
| ZnSO4 | 5 | 67.0 ± 4.0 |
| MnCl2 | 5 | 74.0 ± 10.5 |
| CoCl2 | 5 | 81.4 ± 1.0 |
| CaCl2 | 5 | 123.6 ± 5.7 |
| MgSO4 | 5 | 175.1 ± 8.2 |
| NaCl | 5 | 137.0 ± 13.9 |
| KCl | 5 | 130.0 ± 9.8 |
| NBS | 10 | 50.4 ± 5.4 |
| EDTA | 10 | 142.0 ± 11.8 |
| SDS | 10 | 10.1 ± 1.0 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kotogán, A.; Zambrano, C.; Kecskeméti, A.; Varga, M.; Szekeres, A.; Papp, T.; Vágvölgyi, C.; Takó, M. An Organic Solvent-Tolerant Lipase with Both Hydrolytic and Synthetic Activities from the Oleaginous Fungus Mortierella echinosphaera. Int. J. Mol. Sci. 2018, 19, 1129. https://doi.org/10.3390/ijms19041129
Kotogán A, Zambrano C, Kecskeméti A, Varga M, Szekeres A, Papp T, Vágvölgyi C, Takó M. An Organic Solvent-Tolerant Lipase with Both Hydrolytic and Synthetic Activities from the Oleaginous Fungus Mortierella echinosphaera. International Journal of Molecular Sciences. 2018; 19(4):1129. https://doi.org/10.3390/ijms19041129
Chicago/Turabian StyleKotogán, Alexandra, Carolina Zambrano, Anita Kecskeméti, Mónika Varga, András Szekeres, Tamás Papp, Csaba Vágvölgyi, and Miklós Takó. 2018. "An Organic Solvent-Tolerant Lipase with Both Hydrolytic and Synthetic Activities from the Oleaginous Fungus Mortierella echinosphaera" International Journal of Molecular Sciences 19, no. 4: 1129. https://doi.org/10.3390/ijms19041129