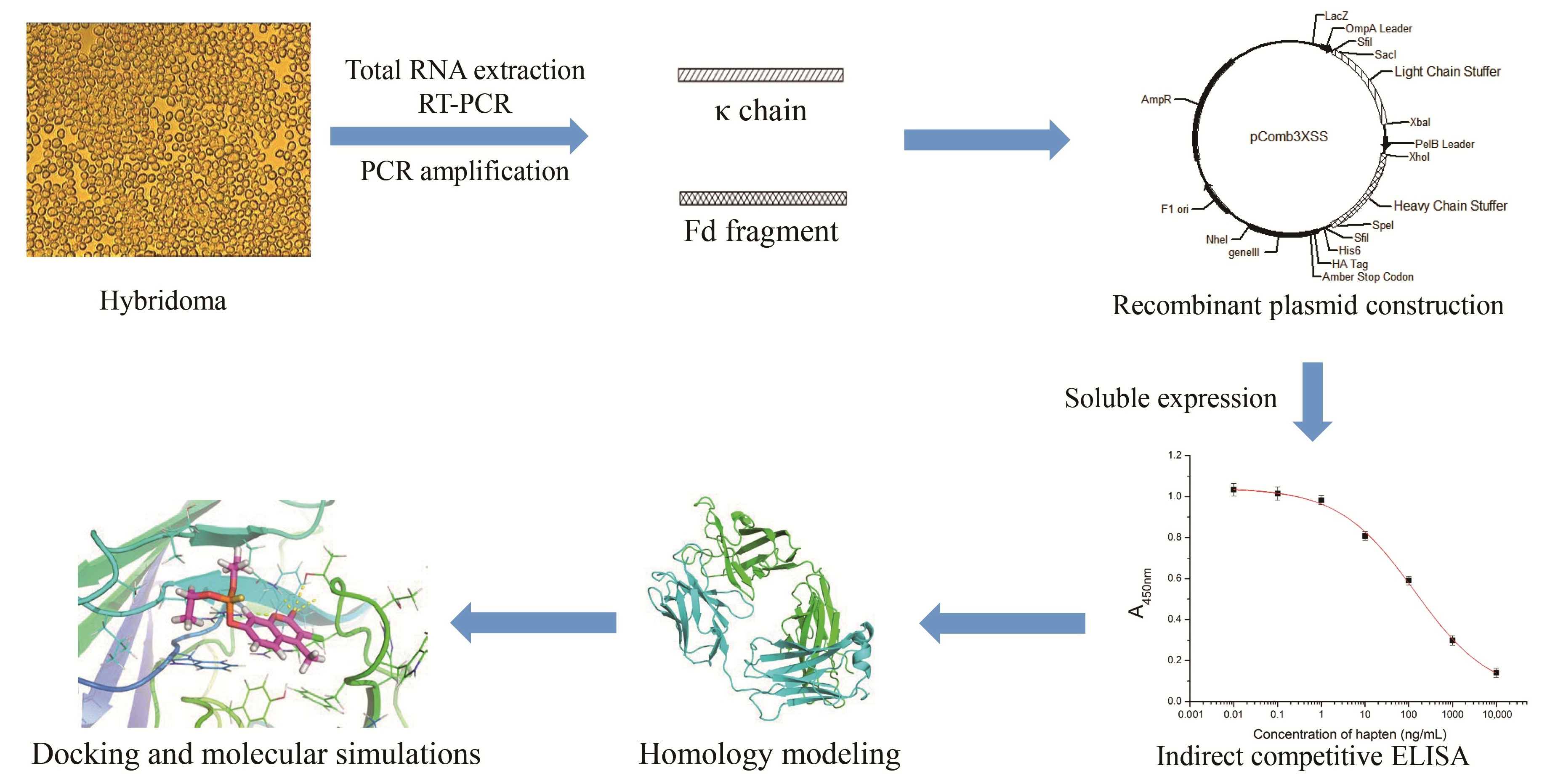
Production of Antigen-Binding Fragment against O,O-Diethyl Organophosphorus Pesticides and Molecular Dynamics Simulations of Antibody Recognition


Abstract
:
1. Introduction
2. Results
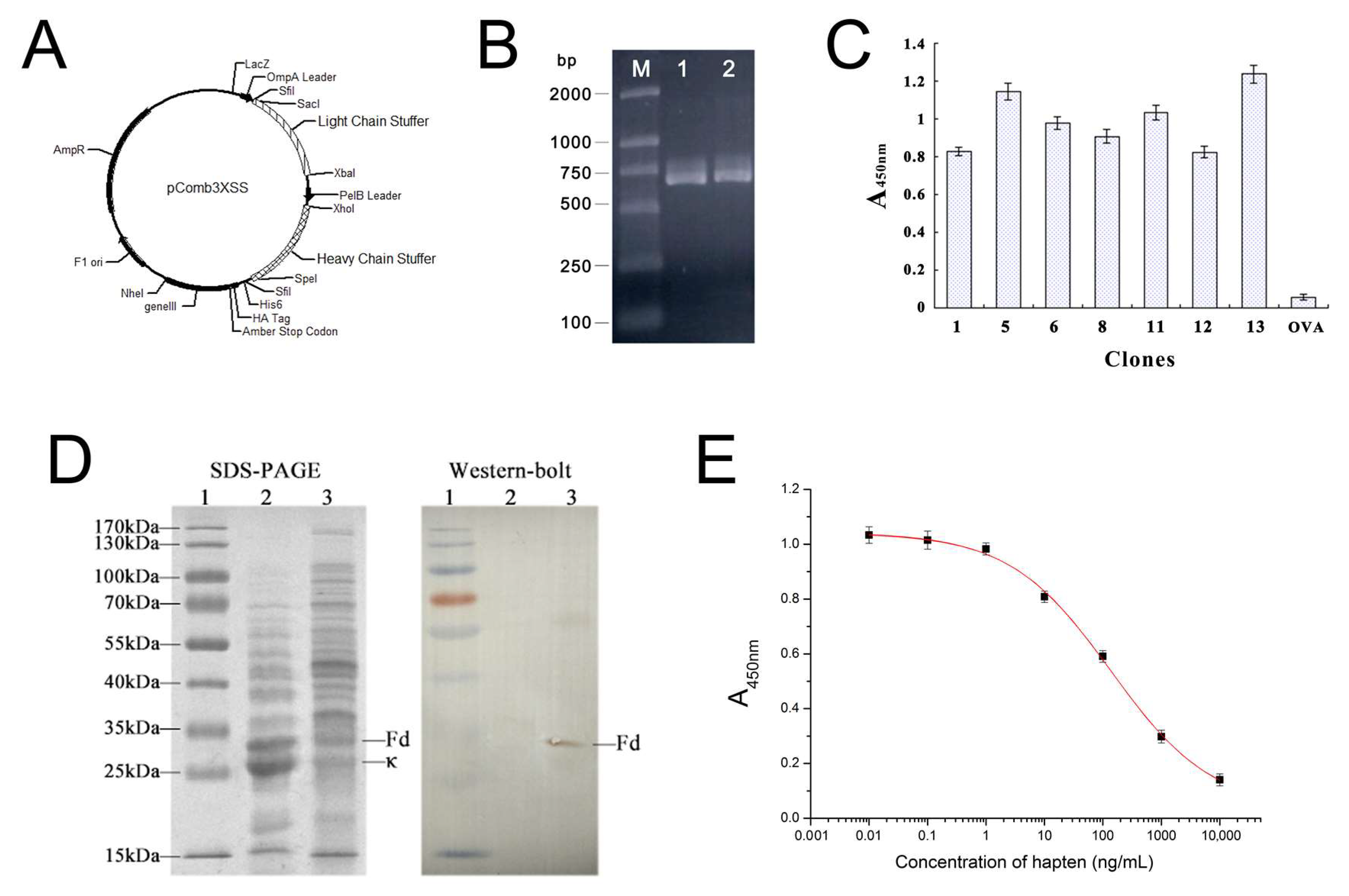
2.1. Amplification of Anti-DOPs Fab Gene Fragment and Construction of the Expression Vector
2.2. Phage Displaying of Anti-DOPs Fab Fragment
2.3. DNA Sequence Analysis
2.4. SDS-PAGE and Western Blotting Analysis of Soluble Anti-DOPs Fab Fragment
2.5. Broad-Specific Binding of Fab Fragment to DOPs
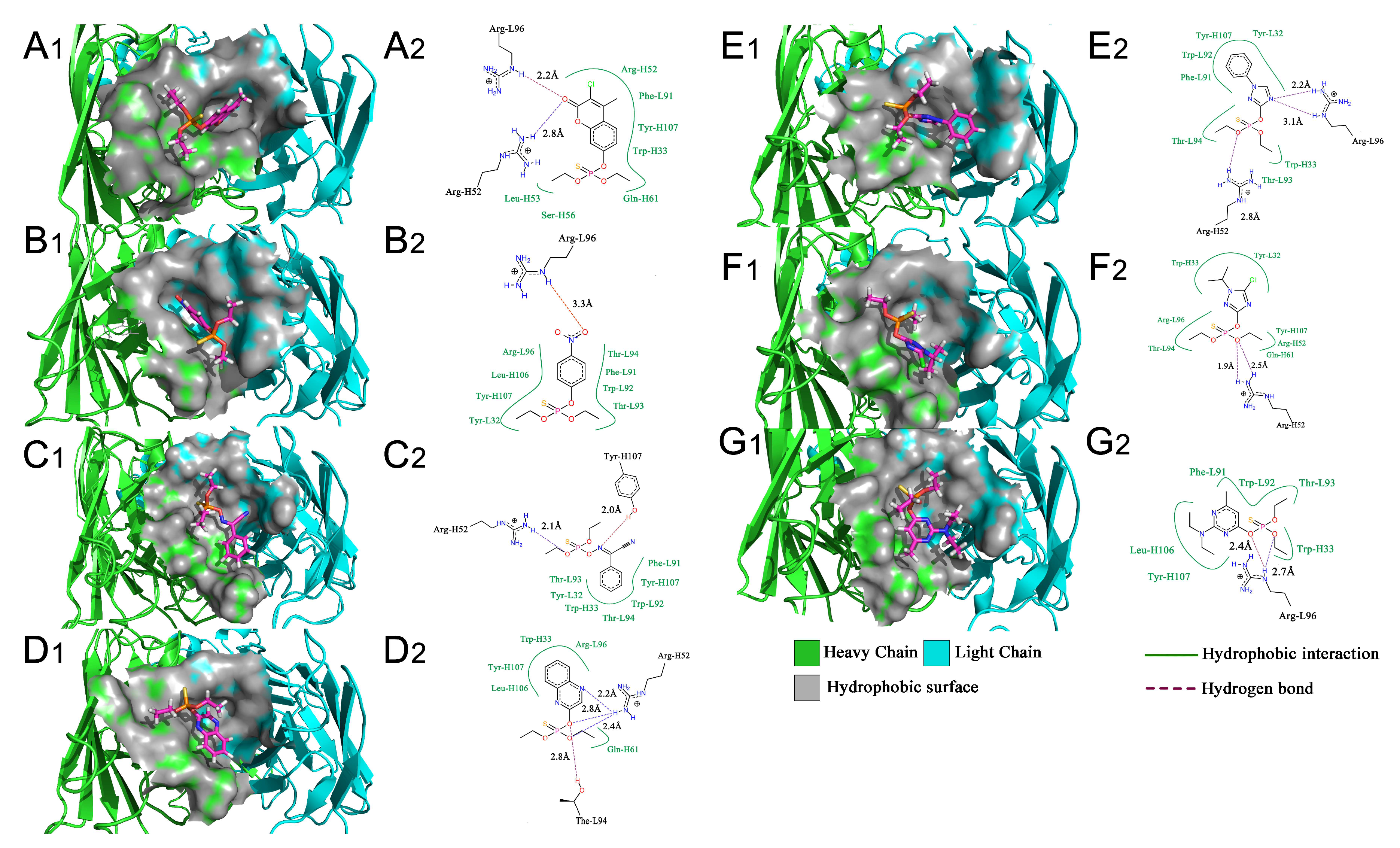
2.6. Construction of the Fab 3D Structure
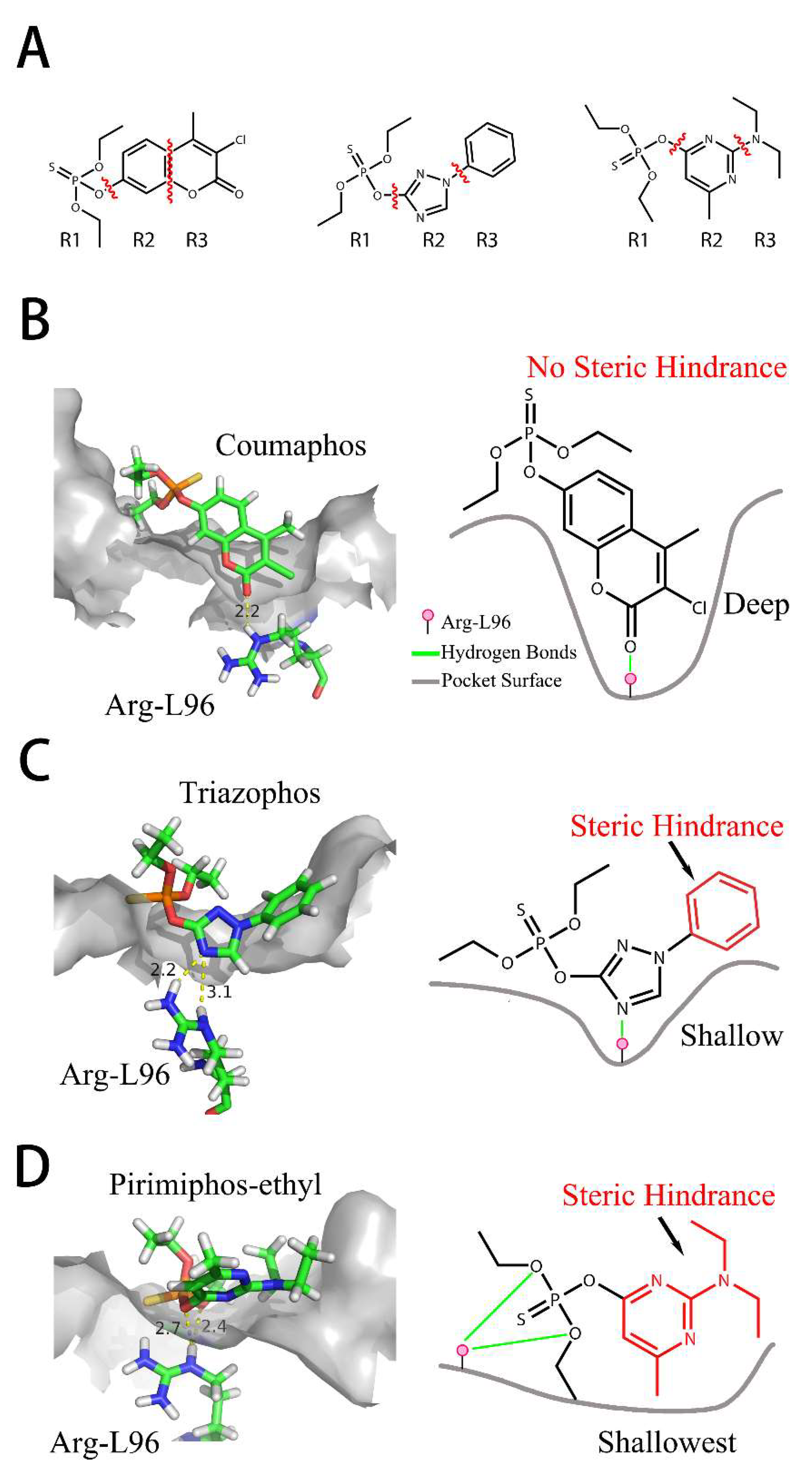
2.7. Interaction of Fab and DOPs
3. Discussion
4. Materials and Methods
4.1. Construction of Anti-DOPs Fab Fragment
4.2. Construction of Recombinant Plasmid
4.3. Phage Display
4.4. Expression of Anti-DOPs Fab Fragment
4.5. SDS-PAGE and Western Blot Analysis
4.6. Indirect Competitive ELISA
4.7. Cross-Reactivity Study
4.8. Homology Modeling and Molecular Simulations
4.9. Site-Specific Mutagenesis
5. Conclusions
Acknowledgments
Author Contributions
Conflicts of Interest
References
- Lu, C.; Barr, D.B.; Pearson, M.A.; Waller, L.A. Dietary Intake and Its Contribution to Longitudinal Organophosphorus Pesticide Exposure in Urban/Suburban Children. Environ. Health Perspect. 2008, 116, 537–542. [Google Scholar] [CrossRef] [PubMed]
- Rezg, R.; Mornagui, B.; Benahmed, M.; Gharsalla Chouchane, S.; Belhajhmida, N.; Abdeladhim, M.; Kamoun, A.; El-fazaa, S.; Gharbi, N. Malathion exposure modulates hypothalamic gene expression and induces dyslipedemia in Wistar rats. Food Chem. Toxicol. 2010, 48, 1473–1477. [Google Scholar] [CrossRef] [PubMed]
- Su, R.; Xu, X.; Wang, X.; Li, D.; Li, X.; Zhang, H.; Yu, A. Determination of organophosphorus pesticides in peanut oil by dispersive solid phase extraction gas chromatography–mass spectrometry. J. Chromatogr. B 2011, 879, 3423–3428. [Google Scholar] [CrossRef] [PubMed]
- Inoue, S.; Saito, T.; Mase, H.; Suzuki, Y.; Takazawa, K.; Yamamoto, I.; Inokuchi, S. Rapid simultaneous determination for organophosphorus pesticides in human serum by LC–MS. J. Pharm. Biomed. Anal. 2007, 44, 258–264. [Google Scholar] [CrossRef] [PubMed]
- Boro, R.C.; Singh, K.V.; Suri, C.R. Characterization of Hapten-Protein Conjugates: Antibody Generation and Immunoassay Development for Chlorophenoxyacetic Acid Pesticides. J. AOAC Int. 2009, 92, 1773–1779. [Google Scholar] [PubMed]
- Spinks, C.A. Broad-specificity immunoassay of low molecular weight food contaminants: New paths to Utopia! Trends Food Sci. Technol. 2000, 11, 210–217. [Google Scholar] [CrossRef]
- Alcocer, M.J.C.; Dillon, P.P.; Manning, B.M.; Doyen, C.; Lee, H.A.; Daly, S.J.; O’Kennedy, R.; Morgan, M.R.A. Use of Phosphonic Acid as a Generic Hapten in the Production of Broad Specificity Anti-Organophosphate Pesticide Antibody. J. Agric. Food Chem. 2000, 48, 2228–2233. [Google Scholar] [CrossRef] [PubMed]
- Liang, Y.; Liu, X.J.; Liu, Y.; Yu, X.Y.; Fan, M.T. Synthesis of three haptens for the class-specific immunoassay of O,O-dimethyl organophosphorus pesticides and effect of hapten heterology on immunoassay sensitivity. Anal. Chim. Acta 2008, 615, 174–183. [Google Scholar] [CrossRef] [PubMed]
- Piao, Y.Z.; Kim, Y.J.; Kim, Y.A.; Lee, H.; Hammock, B.D.; Lee, Y.T. Development of ELISAs for the Class-Specific Determination of Organophosphorus Pesticides. J. Agric. Food Chem. 2009, 57, 10004–10013. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Shen, G.; Zhu, H.; Jiang, G. A Class-Specific Enzyme-Linked Immunosorbent Assay Based on Magnetic Particles for Multiresidue Organophosphorus Pesticides. J. Agric. Food Chem. 2010, 58, 2801–2806. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Shen, Y.; Zheng, W.; Beier, R.C.; Xie, G.; Dong, J.; Yang, J.; Wang, H.; Lei, H.; She, Z.; et al. Broad-Specificity Immunoassay for O,O-Diethyl Organophosphorus Pesticides: Application of Molecular Modeling to Improve Assay Sensitivity and Study Antibody Recognition. Anal. Chem. 2010, 82, 9314–9321. [Google Scholar] [CrossRef] [PubMed]
- Xu, Z.; Sun, W.; Yang, J.; Jiang, Y.; Campbell, K.; Shen, Y.; Lei, H.; Zeng, D.; Wang, H.; Sun, Y. Development of a Solid-Phase Extraction Coupling Chemiluminescent Enzyme Immunoassay for Determination of Organophosphorus Pesticides in Environmental Water Samples. J. Agric. Food Chem. 2012, 60, 2069–2075. [Google Scholar] [CrossRef] [PubMed]
- Clark, L.A.; Boriack-Sjodin, P.A.; Eldredge, J.; Fitch, C.; Friedman, B.; Hanf, K.J.M.; Jarpe, M.; Liparoto, S.F.; Li, Y.; Lugovskoy, A.; et al. Affinity enhancement of an in vivo matured therapeutic antibody using structure-based computational design. Protein Sci. 2006, 15, 949–960. [Google Scholar] [CrossRef] [PubMed]
- Kusharyoto, W.; Pleiss, J.; Bachmann, T.T.; Schmid, R.D. Mapping of a hapten-binding site: Molecular modeling and site-directed mutagenesis study of an anti-atrazine antibody. Protein Eng. 2002, 15, 233–241. [Google Scholar] [CrossRef] [PubMed]
- Kavanagh, O.; Elliott, C.T.; Campbell, K. Progress in the development of immunoanalytical methods incorporating recombinant antibodies to small molecular weight biotoxins. Anal. Bioanal. Chem. 2015, 407, 2749–2770. [Google Scholar] [CrossRef] [PubMed]
- Corisdeo, S.; Wang, B. Functional expression and display of an antibody Fab fragment in Escherichia coli: Study of vector designs and culture conditions. Protein Expr. Purif. 2004, 34, 270–279. [Google Scholar] [CrossRef] [PubMed]
- Wu, X.; Sereno, A.J.; Huang, F.; Lewis, S.M.; Lieu, R.L.; Weldon, C.; Torres, C.; Fine, C.; Batt, M.A.; Fitchett, J.R.; et al. Fab-based bispecific antibody formats with robust biophysical properties and biological activity. MABS 2015, 7, 470–482. [Google Scholar] [CrossRef] [PubMed]
- Teerinen, T.; Valjakka, J.; Rouvinen, J.; Takkinen, K. Structure-based Stability Engineering of the Mouse IgG1 Fab Fragment by Modifying Constant Domains. J. Mol. Biol. 2006, 361, 687–697. [Google Scholar] [CrossRef] [PubMed]
- Rao, M.F.; Li, Y.J.; Dong, J.X.; Wu, W.J.; Xu, Z.L.; Sun, Y.M.; Wang, H. Production and characterization of a single-chain Fab fragment for the detection of O,O-diethyl organophosphorus pesticides. Anal. Methods 2016, 8, 3140–3147. [Google Scholar] [CrossRef]
- Chames, P.; Hufton, S.E.; Coulie, P.G.; Uchanska-Ziegler, B.; Hoogenboom, H.R. Direct selection of a human antibody fragment directed against the tumor T-cell epitope HLA-A1-MAGE-A1 from a nonimmunized phage-Fab library. Proc. Nat. Acad. Sci. USA 2000, 97, 7969–7974. [Google Scholar] [CrossRef] [PubMed]
- Moreno De Alboran, I.; Martinez-Alonso, C.; Barbasi, C.F.I.; Burton, D.R.; Ditzel, H.J. Human monoclonal Fab fragments specific for viral antigens from combinatorial IgA libraries. Immunotechnology 1995, 1, 21–28. [Google Scholar] [CrossRef]
- Lai, Y.; Wu, Y.; Tsai, Y.; Tsai, Y.; Lin, Y.; Lai, S.; Huang, C.; Lok, Y.; Hu, C.; Lai, J. Generation of Potent Anti-Vascular Endothelial Growth Factor Neutralizing Antibodies from Mouse Phage Display Library for Cancer Therapy. Int. J. Mol. Sci. 2016, 17, 214. [Google Scholar] [CrossRef] [PubMed]
- Maillaud, C.; Barguil, Y.; Mikulski, M.; Cheze, M.; Pivert, C.; Deveaux, M.; Lapostolle, F. First successful curative use of digoxin-specific Fab antibody fragments in a life-threatening coconut crab (Birgus latro L.) poisoning. Toxicon 2012, 60, 1013–1017. [Google Scholar] [CrossRef] [PubMed]
- He, K.; Du, X.; Sheng, W.; Zhou, X.; Wang, J.; Wang, S. Crystal Structure of the Fab Fragment of an Anti-ofloxacin Antibody and Exploration of Its Specific Binding. J. Agric. Food Chem. 2016, 64, 2627–2634. [Google Scholar] [CrossRef] [PubMed]
- Chatterjee, D.K.; Esposito, D. Enhanced soluble protein expression using two new fusion tags. Protein Expr. Purif. 2006, 46, 122–129. [Google Scholar] [CrossRef] [PubMed]
- Moghaddam, A.; Løbersli, I.; Gebhardt, K.; Braunagel, M.; Marvik, O.J. Selection and characterisation of recombinant single-chain antibodies to the hapten Aflatoxin-B1 from naive recombinant antibody libraries. J. Immunol. Methods 2001, 254, 169–181. [Google Scholar] [CrossRef]
- Korpimäki, T.; Brockmann, E.; Kuronen, O.; Saraste, M.; Lamminmäki, U.; Tuomola, M. Engineering of a Broad Specificity Antibody for Simultaneous Detection of 13 Sulfonamides at the Maximum Residue Level. J. Agric. Food Chem. 2004, 52, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Nishi, K.; Ishiuchi, M.; Morimune, K.; Ohkawa, H. Molecular and Immunochemical Characteristics of Monoclonal and Recombinant Antibodies Selective for the Triazine Herbicide Simetryn and Application to Environmental Analysis. J. Agric. Food Chem. 2005, 53, 5096–5104. [Google Scholar] [CrossRef] [PubMed]
- Kramer, K.; Hock, B. Recombinant antibodies for environmental analysis. Anal. Bioanal. Chem. 2003, 377, 417–426. [Google Scholar] [CrossRef] [PubMed]
- Alcocer, M.J.C.; Doyen, C.; Lee, H.A.; Morgan, M.R.A. Properties of Polyclonal, Monoclonal, and Recombinant Antibodies Recognizing the Organophosphorus Pesticide Chlorpyrifos-ethyl. J. Agric. Food Chem. 2000, 48, 4053–4059. [Google Scholar] [CrossRef] [PubMed]
- Wu, T.T. An analysis of the sequences of the variable regions of bence jones proteins and myeloma light chains and their implications for antibody complementarity. J. Exp. Med. 1970, 132, 211–250. [Google Scholar] [CrossRef] [PubMed]
- Kopp, J. The SWISS-MODEL Repository: New features and functionalities. Nucleic Acids Res. 2006, 34, D315–D318. [Google Scholar] [CrossRef] [PubMed]
- Vidarte, L.; Pastor, C.; Mas, S.; Blazquez, A.B.; de Los, R.V.; Guerrero, R.; Vivanco, F. Serine 132 is the C3 covalent attachment point on the CH1 domain of human IgG1. J. Biol. Chem. 2001, 276, 38217–38223. [Google Scholar] [PubMed]
- Erlandsson, A.; Holm, P.; Ullén, A.; Stigbrand, T.; Sundström, B.E. Studies of the interactions between the anticytokeratin 8 monoclonal antibody TS1, its antigen and its anti-idiotypic antibody αTS1. J. Mol. Recognit. 2003, 16, 157–163. [Google Scholar] [CrossRef] [PubMed]
- Niccheri, F.; Real-Fernàndez, F.; Ramazzotti, M.; Lolli, F.; Rossi, G.; Rovero, P.; Degl’Innocenti, D. Human recombinant domain antibodies against multiple sclerosis antigenic peptide CSF114(Glc). J. Mol. Recognit. 2014, 27, 618–626. [Google Scholar] [CrossRef] [PubMed]
- Lape, M.; Paula, S.; Ball, W.J. A molecular model for cocaine binding by the immunotherapeutic human/mouse chimeric monoclonal antibody 2E2. Eur. J. Med. Chem. 2010, 45, 2291–2298. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Li, P.; Zhang, Q.; Li, Y.; Zhang, W.; Ding, X. Molecular Characterization of Monoclonal Antibodies against Aflatoxins: A Possible Explanation for the Highest Sensitivity. Anal. Chem. 2012, 84, 5229–5235. [Google Scholar] [CrossRef] [PubMed]
- Jiang, W.; Li, W.; Hong, Y.; Wang, S.; Fang, B. Cloning, Expression, Mutagenesis Library Construction of Glycerol Dehydratase, and Binding Mode Simulation of Its Reactivase with Ligands. Appl. Biochem. Biotechnol. 2016, 178, 739–752. [Google Scholar] [CrossRef] [PubMed]
- Guerler, A.; Moll, S.; Weber, M.; Meyer, H.; Cordes, F. Selection and flexible optimization of binding modes from conformation ensembles. Biosystems 2008, 92, 42–48. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Wang, Y.; Li, Z.; Li, J.; Dong, Z. Humanization of a mouse monoclonal antibody neutralizing TNF-α by guided selection. J. Immunol. Methods 2000, 241, 171–184. [Google Scholar] [CrossRef]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Gan, C.F.; Wang, B.F.; Huang, J.Y.; Qileng, A.; He, Z.Y.; Lei, H.T.; Liu, W.P.; Liu, Y.J. Multiple amplified enzyme-free electrochemical immunosensor based on G-quadruplex/hemin functionalized mesoporous silica with redox-active intercalators for microcystin-LR detection. Biosens. Bioelectron. 2017, 98, 126–133. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| DOPs | Structure | Fab | MAb | ||
|---|---|---|---|---|---|
| IC50 (ng/mL) | CR (%) | IC50 (ng/mL) | CR (%) | ||
| Hapten 1 |  | 135.7 | 100.0 | 178.3 | 100.0 |
| Coumaphos |  | 1.0 | 17,747.0 | 7.6 | 2931.9 |
| Parathion |  | 3.1 | 4422.1 | 1.1 | 16,264.4 |
| Dichlofenthion |  | 41.2 | 357.7 | 62.7 | 308.7 |
| Phoxim |  | 44.9 | 310.9 | 64.0 | 286.3 |
| Quinalphos |  | 75.3 | 185.1 | 100.2 | 182.9 |
| Triazophos |  | 106.6 | 137.4 | 136.2 | 141.3 |
| Phosalone |  | 263.0 | 65.4 | 281.6 | 80.2 |
| Phorate |  | 325.8 | 37.4 | 837.9 | 19.1 |
| Chlorpyrifos |  | 895.3 | 18.3 | 2087.6 | 10.3 |
| Bromophos-ethyl |  | 1064.8 | 17.3 | 2673.2 | 9.1 |
| Sulfotep |  | 1547.8 | 9.7 | 3.5843 | 5.5 |
| Isazophos |  | 2289.5 | 6.4 | 6989.9 | 2.8 |
| Diazinon |  | 4515.6 | 3.2 | 10,865.0 | 1.7 |
| Pirimiphos-ethyl |  | 6333.0 | 2.5 | 12,266.9 | 1.7 |
| Fab Dilution Multiple | Titer * | |
|---|---|---|
| Original | Mutated | |
| 1 | 2.3045 | 0.1310 |
| 2 | 1.0645 | 0.1280 |
| 3 | 0.5510 | 0.1255 |
| 4 | 0.4265 | 0.1090 |
| 5 | 0.4280 | 0.1395 |
| 6 | 0.3145 | 0.1170 |
| 7 | 0.2860 | 0.1005 |
| Blank | 0.0980 | 0.1380 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, Z.-J.; Zhang, X.; Wang, B.-F.; Rao, M.-F.; Wang, H.; Lei, H.-T.; Liu, H.; Zhang, Y.; Sun, Y.-M.; Xu, Z.-L. Production of Antigen-Binding Fragment against O,O-Diethyl Organophosphorus Pesticides and Molecular Dynamics Simulations of Antibody Recognition. Int. J. Mol. Sci. 2018, 19, 1381. https://doi.org/10.3390/ijms19051381
Chen Z-J, Zhang X, Wang B-F, Rao M-F, Wang H, Lei H-T, Liu H, Zhang Y, Sun Y-M, Xu Z-L. Production of Antigen-Binding Fragment against O,O-Diethyl Organophosphorus Pesticides and Molecular Dynamics Simulations of Antibody Recognition. International Journal of Molecular Sciences. 2018; 19(5):1381. https://doi.org/10.3390/ijms19051381
Chicago/Turabian StyleChen, Zi-Jian, Xuan Zhang, Bing-Feng Wang, Mei-Fang Rao, Hong Wang, Hong-Tao Lei, Hui Liu, Yan Zhang, Yuan-Ming Sun, and Zhen-Lin Xu. 2018. "Production of Antigen-Binding Fragment against O,O-Diethyl Organophosphorus Pesticides and Molecular Dynamics Simulations of Antibody Recognition" International Journal of Molecular Sciences 19, no. 5: 1381. https://doi.org/10.3390/ijms19051381