Relaxin 2/RXFP1 Signaling Induces Cell Invasion via the β-Catenin Pathway in Endometrial Cancer

 ,
,
Abstract
:1. Introduction
2. Results
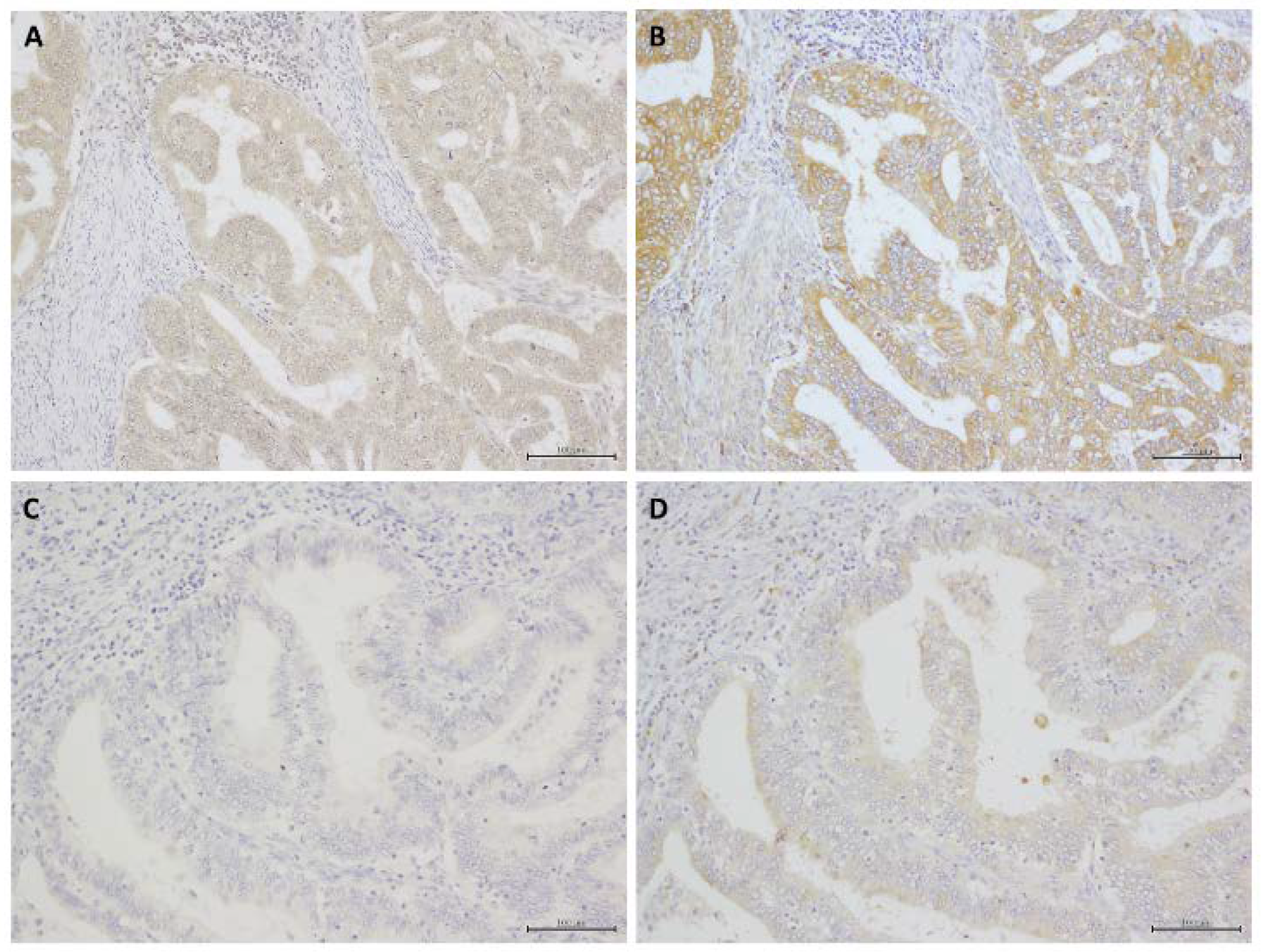
2.1. Immunohistochemistry
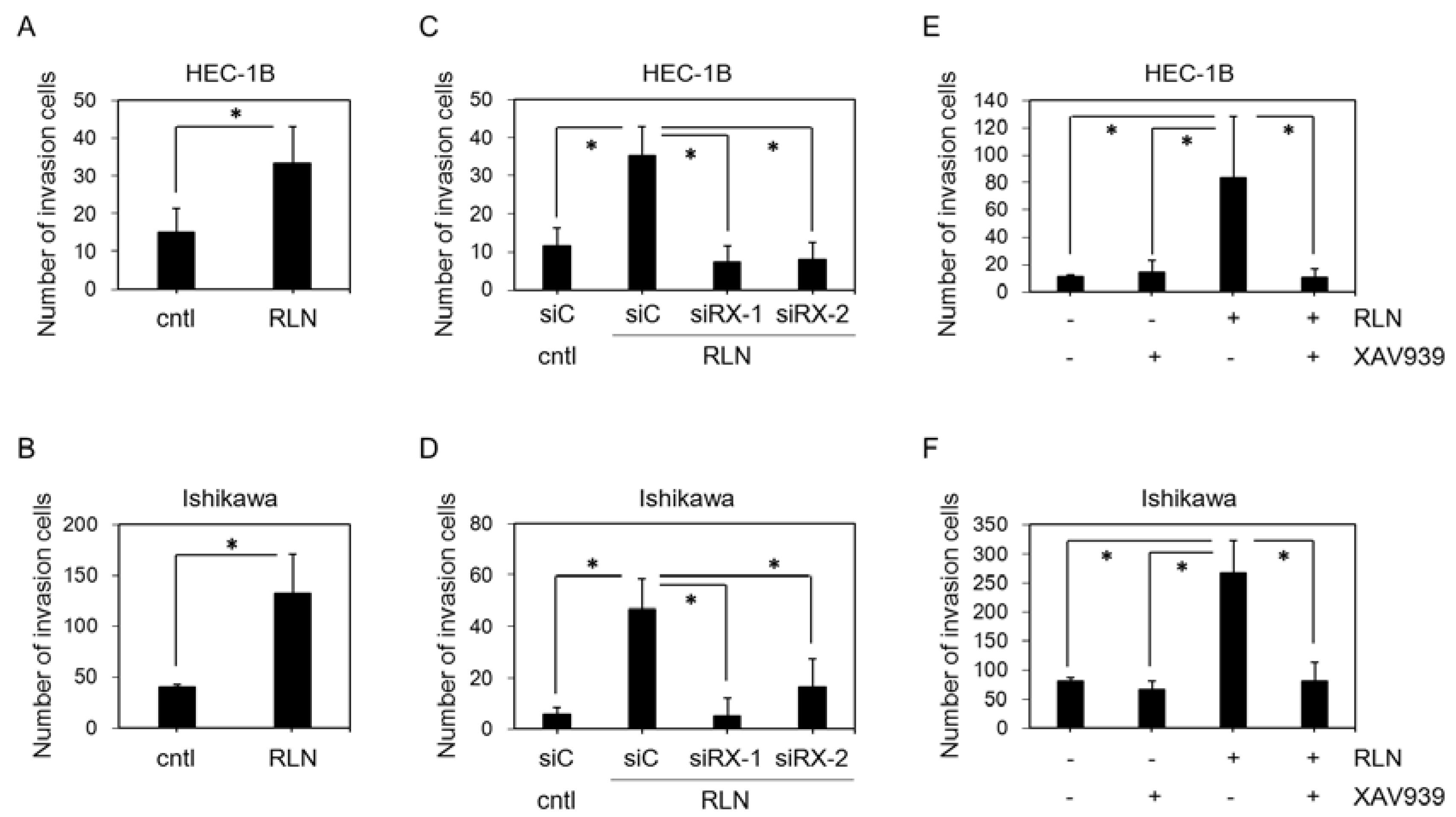
2.2. Effect of RLN2 on HEC-1B and Ishikawa Cell Invasiveness
2.3. Effect of siRXFP1 on HEC-1B and Ishikawa Cell Invasiveness
2.4. Effect of XAV939 on HEC-1B and Ishikawa Cell Invasiveness
2.5. Effect of RLN2 on β-Catenin Phosphorylation in Cancer Cell Lines
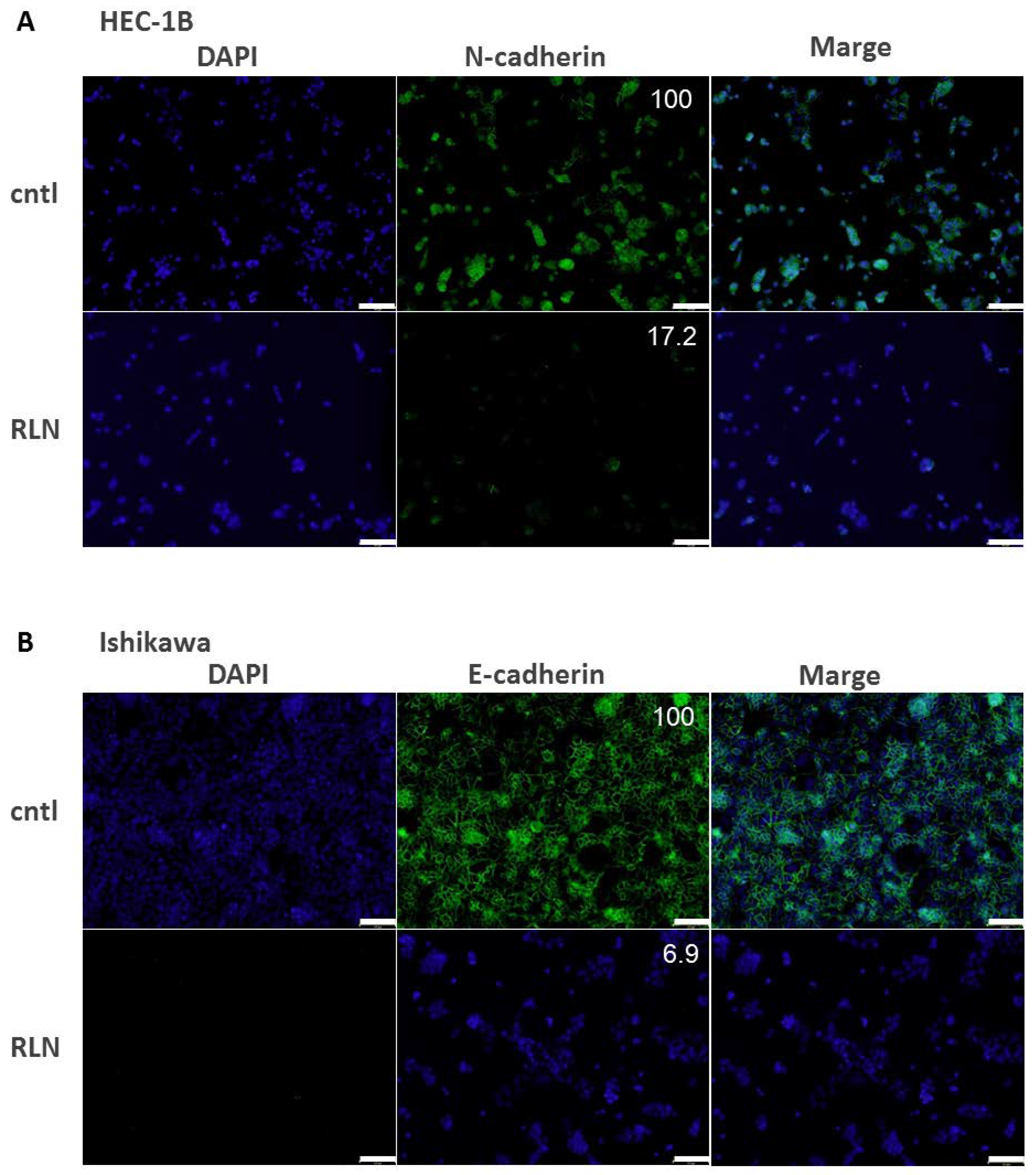
2.6. Effect of RLN2 on Cadherin Expression in HEC-1B and Ishikawa Cells
3. Discussion
4. Materials and Methods
4.1. Patients
4.2. Immunohistochemistry
4.3. Cell Lines and Culture Conditions
4.4. Matrigel Invasion Assays
4.5. Small Interfering RNA Transfection
4.6. Immunoblotting
4.7. Immunofluorescence Cytochemistry
4.8. Statistical Analysis
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
References
- Kitchener, H.C.; Trimble, E.L. Endometrial cancer state of the science meeting. Int. J. Gynecol. Cancer 2009, 19, 134–140. [Google Scholar] [CrossRef] [PubMed]
- AlHilli, M.M.; Podratz, K.C.; Dowdy, S.C.; Bakkum-Gamez, J.N.; Weaver, A.L.; McGree, M.E.; Keeney, G.L.; Cliby, W.A.; Mariani, A. Risk-scoring system for the individualized prediction of lymphatic dissemination in patients with endometrioid endometrial cancer. Gynecol. Oncol. 2013, 131, 103–108. [Google Scholar] [CrossRef] [PubMed]
- Kato, T.; Watari, H.; Endo, D.; Mitamura, T.; Odagiri, T.; Konno, Y.; Hosaka, M.; Kobayashi, N.; Todo, Y.; Sudo, S.; et al. New revised FIGO 2008 staging system for endometrial cancer produces better discrimination in survival compared with the 1988 staging system. J. Surg. Oncol. 2012, 106, 938–941. [Google Scholar] [CrossRef] [PubMed]
- Creutzberg, C.L.; van Putten, W.L.; Koper, P.C.; Lybeert, M.L.; Jobsen, J.J.; Warlam-Rodenhuis, C.C.; De Winter, K.A.; Lutgens, L.C.; van den Bergh, A.C.; van de Steen-Banasik, E.; et al. Surgery and postoperative radiotherapy versus surgery alone for patients with stage-1 endometrial carcinoma: Multicentre randomised trial. Lancet 2000, 355, 1404–1411. [Google Scholar] [CrossRef]
- Orr, J.W., Jr.; Roland, P.Y.; Leichter, D.; Orr, P.F. Endometrial cancer: Is surgical staging necessary? Curr. Opin. Oncol. 2001, 13, 408–412. [Google Scholar] [CrossRef] [PubMed]
- Keys, H.M.; Roberts, J.A.; Brunetto, V.L.; Zaino, R.J.; Spirtos, N.M.; Bloss, J.D.; Pearlman, A.; Maiman, M.A.; Bell, J.G. A phase III trial of surgery with or without adjunctive external pelvic radiation therapy in intermediate risk endometrial adenocarcinoma: A Gynecologic Oncology Group study. Gynecol. Oncol. 2004, 92, 744–751. [Google Scholar] [CrossRef] [PubMed]
- Ivell, R.; Einspanier, A. Relaxin peptides are new global players. Trends Endocrinol. Metab. 2002, 13, 343–348. [Google Scholar] [CrossRef]
- Bathgate, R.A.; Halls, M.L.; van der Westhuizen, E.T.; Callander, G.E.; Kocan, M.; Summers, R.J. Relaxin family peptides and their receptors. Physiol. Rev. 2013, 93, 405–480. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.Y.; Kudo, M.; Chen, T.; Nakabayashi, K.; Bhalla, A.; van der Spek, P.J.; van Duin, M.; Hsueh, A.J. The three subfamilies of leucine-rich repeat-containing G protein-coupled receptors (LGR): Identification of LGR6 and LGR7 and the signaling mechanism for LGR7. Mol. Endocrinol. 2000, 14, 1257–1271. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.Y.; Nakabayashi, K.; Nishi, S.; Kumagai, J.; Kudo, M.; Sherwood, O.D.; Hsueh, A.J. Activation of orphan receptors by the hormone relaxin. Science 2002, 295, 671–674. [Google Scholar] [CrossRef] [PubMed]
- Kamat, A.A.; Feng, S.; Agoulnik, I.U.; Kheradmand, F.; Bogatcheva, N.V.; Coffey, D.; Sood, A.K.; Agoulnik, A.I. The role of relaxin in endometrial cancer. Cancer Biol. Ther. 2006, 5, 71–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marshall, S.A.; Senadheera, S.N.; Parry, L.J.; Girling, J.E. The Role of Relaxin in Normal and Abnormal Uterine Function during the Menstrual Cycle and Early Pregnancy. Reprod. Sci. 2017, 24, 342–354. [Google Scholar] [CrossRef] [PubMed]
- Feng, S.; Agoulnik, I.U.; Bogatcheva, N.V.; Kamat, A.A.; Kwabi-Addo, B.; Li, R.; Ayala, G.; Ittmann, M.M.; Agoulnik, A.I. Relaxin promotes prostate cancer progression. Clin. Cancer Res. 2007, 13, 1695–1702. [Google Scholar] [CrossRef] [PubMed]
- Shabanpoor, F.; Separovic, F.; Wade, J.D. The human insulin superfamily of polypeptide hormones. Vitam. Horm. 2009, 80, 1–31. [Google Scholar] [PubMed]
- Nair, V.B.; Samuel, C.S.; Separovic, F.; Hossain, M.A.; Wade, J.D. Human relaxin-2: Historical perspectives and role in cancer biology. Amino Acids 2012, 43, 1131–1140. [Google Scholar] [CrossRef] [PubMed]
- Mazoujian, G.; Bryant-Greenwood, G.D. Relaxin in breast tissue. Lancet 1990, 335, 298–299. [Google Scholar] [CrossRef]
- Goldsmith, L.T.; Weiss, G. Relaxin in human pregnancy. Ann. N. Y. Acad. Sci. 2009, 1160, 130–135. [Google Scholar] [CrossRef] [PubMed]
- Mookerjee, I.; Unemori, E.N.; DU, X.J.; Tregear, G.W.; Samuel, C.S. Relaxin modulates fibroblast function, collagen production, and matrix metalloproteinase-2 expression by cardiac fibroblasts. Ann. N. Y. Acad. Sci. 2005, 1041, 190–193. [Google Scholar] [CrossRef] [PubMed]
- Sacchi, T.B.; Bani, D.; Brandi, M.L.; Falchetti, A.; Bigazzi, M. Relaxin influences growth, differentiation and cell-cell adhesion of human breast-cancer cells in culture. Int. J. Cancer 1994, 57, 129–134. [Google Scholar] [CrossRef] [PubMed]
- Bani, D.; Flagiello, D.; Poupon, M.F.; Nistri, S.; Poirson-Bichat, F.; Bigazzi, M.; Bani Sacchi, T. Relaxin promotes differentiation of human breast cancer cells MCF-7 transplanted into nude mice. Virchows Arch. 1999, 435, 509–519. [Google Scholar] [CrossRef] [PubMed]
- Binder, C.; Hagemann, T.; Husen, B.; Schulz, M.; Einspanier, A. Relaxin enhances in-vitro invasiveness of breast cancer cell lines by up-regulation of matrix metalloproteases. Mol. Hum. Reprod. 2002, 8, 789–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hombach-Klonisch, S.; Bialek, J.; Trojanowicz, B.; Weber, E.; Holzhausen, H.J.; Silvertown, J.D.; Summerlee, A.J.; Dralle, H.; Hoang-Vu, C.; Klonisch, T. Relaxin enhances the oncogenic potential of human thyroid carcinoma cells. Am. J. Pathol. 2006, 169, 617–632. [Google Scholar] [CrossRef] [PubMed]
- Thompson, V.C.; Hurtado-Coll, A.; Turbin, D.; Fazli, L.; Lehman, M.L.; Gleave, M.E.; Nelson, C.C. Relaxin drives Wnt signaling through upregulation of PCDHY in prostate cancer. Prostate 2010, 70, 1134–1145. [Google Scholar] [CrossRef] [PubMed]
- Kim, B.G.; Kim, J.W.; Kim, S.M.; Go, R.E.; Hwang, K.A.; Choi, K.C. 3,3′-Diindolylmethane Suppressed Cyprodinil-Induced Epithelial-Mesenchymal Transition and Metastatic-Related Behaviors of Human Endometrial Ishikawa Cells via an Estrogen Receptor-Dependent Pathway. Int. J. Mol. Sci. 2018, 19, 189. [Google Scholar] [CrossRef] [PubMed]
- Guo, J.; Liu, C.; Zhou, X.; Xu, X.; Deng, L.; Li, X.; Guan, F. Conditioned Medium from Malignant Breast Cancer Cells Induces an EMT-Like Phenotype and an Altered N-Glycan Profile in Normal Epithelial MCF10A Cells. Int. J. Mol. Sci. 2017, 18, 1528. [Google Scholar] [CrossRef] [PubMed]
- Schmalhofer, O.; Brabletz, S.; Brabletz, T. E-cadherin, beta-catenin, and ZEB1 in malignant progression of cancer. Cancer Metastasis Rev. 2009, 28, 151–166. [Google Scholar] [CrossRef] [PubMed]
- Fan, H.B.; Zhai, Z.Y.; Li, X.G.; Gao, C.Q.; Yan, H.C.; Chen, Z.S.; Wang, X.Q. CDX2 Stimulates the Proliferation of Porcine Intestinal Epithelial Cells by Activating the mTORC1 and Wnt/β-Catenin Signaling Pathways. Int. J. Mol. Sci. 2017, 18, 2447. [Google Scholar] [CrossRef] [PubMed]
- Cao, W.H.; Liu, H.M.; Liu, X.; Li, J.G.; Liang, J.; Liu, M.; Niu, Z.H. Relaxin enhances in-vitro invasiveness of breast cancer cell lines by upregulation of S100A4/MMPs signaling. Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 609–617. [Google Scholar] [PubMed]
- MacLennan, A.H.; Green, R.C.; Grant, P.; Nicolson, R. Ripening of the human cervix and induction of labor with intracervical purified porcine relaxin. Obstet. Gynecol. 1986, 68, 598–601. [Google Scholar] [PubMed]
- Kelly, A.J.; Kavanagh, J.; Thomas, J. Relaxin for cervical ripening and induction of labour. Cochrane Database Syst. Rev. 2001, Cd003103. [Google Scholar] [CrossRef] [PubMed]
- Borgna-Pignatti, C.; Cappellini, M.D.; De Stefano, P.; Del Vecchio, G.C.; Forni, G.L.; Gamberini, M.R.; Ghilardi, R.; Origa, R.; Piga, A.; Romeo, M.A.; et al. Survival and complications in thalassemia. Ann. N. Y. Acad. Sci. 2005, 1054, 40–47. [Google Scholar] [CrossRef] [PubMed]
- Bullesbach, E.E.; Schwabe, C. Total synthesis of human relaxin and human relaxin derivatives by solid-phase peptide synthesis and site-directed chain combination. J. Biol. Chem. 1991, 266, 10754–10761. [Google Scholar] [PubMed]
- Yang, X.; Chen, M.W.; Terry, S.; Vacherot, F.; Chopin, D.K.; Bemis, D.L.; Kitajewski, J.; Benson, M.C.; Guo, Y.; Buttyan, R. A human- and male-specific protocadherin that acts through the wnt signaling pathway to induce neuroendocrine transdifferentiation of prostate cancer cells. Cancer Res. 2005, 65, 5263–5271. [Google Scholar] [CrossRef] [PubMed]
- Karim, R.; Tse, G.; Putti, T.; Scolyer, R.; Lee, S. The significance of the Wnt pathway in the pathology of human cancers. Pathology 2004, 36, 120–128. [Google Scholar] [CrossRef] [PubMed]
- Jiang, Y.G.; Luo, Y.; He, D.L.; Li, X.; Zhang, L.L.; Peng, T.; Li, M.C.; Lin, Y.H. Role of Wnt/β-catenin signaling pathway in epithelial-mesenchymal transition of human prostate cancer induced by hypoxia-inducible factor-1α. Int. J. Urol. 2007, 14, 1034–1039. [Google Scholar] [CrossRef] [PubMed]
- Muller, T.; Choidas, A.; Reichmann, E.; Ullrich, A. Phosphorylation and free pool of beta-catenin are regulated by tyrosine kinases and tyrosine phosphatases during epithelial cell migration. J. Biol. Chem. 1999, 274, 10173–10183. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.; Kemler, R. Molecular organization of the uvomorulin-catenin complex. J. Cell Biol. 1992, 116, 989–996. [Google Scholar] [CrossRef] [PubMed]
- Ozawa, M.; Ringwald, M.; Kemler, R. Uvomorulin-catenin complex formation is regulated by a specific domain in the cytoplasmic region of the cell adhesion molecule. Proc. Natl. Acad. Sci. USA 1990, 87, 4246–4250. [Google Scholar] [CrossRef] [PubMed]
- McCrea, PD.; Gumbiner, B.M. Purification of a 92-kDa cytoplasmic protein tightly associated with the cell-cell adhesion molecule E-cadherin (uvomorulin). Characterization and extractability of the protein complex from the cell cytostructure. J. Biol. Chem. 1991, 266, 4514–4520. [Google Scholar] [PubMed]
- Kawanishi, J.; Kato, J.; Sasaki, K.; Fujii, S.; Watanabe, N.; Niitsu, Y. Loss of E-cadherin-dependent cell-cell adhesion due to mutation of the beta-catenin gene in a human cancer cell line, HSC-39. Mol. Cell Biol. 1995, 15, 1175–1181. [Google Scholar] [CrossRef] [PubMed]
- Nishimura, M.; Saito, T.; Yamasaki, H.; Kudo, R. Suppression of gap junctional intercellular communication via 5′ CpG island methylation in promoter region of E-cadherin gene in endometrial cancer cells. Carcinogenesis 2003, 24, 1615–1623. [Google Scholar] [CrossRef] [PubMed]
- Halls, M.L.; Bathgate, R.A.; Summers, R.J. Comparison of signaling pathways activated by the relaxin family peptide receptors, RXFP1 and RXFP2, using reporter genes. J. Pharmacol. Exp. Ther. 2007, 320, 281–290. [Google Scholar] [CrossRef] [PubMed]
- Kumagai, J.; Hsu, S.Y.; Matsumi, H.; Roh, J.S.; Fu, P.; Wade, J.D.; Bathgate, R.A.; Hsueh, A.J. INSL3/Leydig insulin-like peptide activates the LGR8 receptor important in testis descent. J. Biol. Chem. 2002, 277, 31283–31286. [Google Scholar] [CrossRef] [PubMed]
- Patil, N.A.; Rosengren, K.J.; Separovic, F.; Wade, J.D.; Bathgate, R.A.D.; Hossain, M.A. Relaxin family peptides: Structure-activity relationship studies. Br. J. Pharmacol. 2017, 174, 950–961, Erratum in 2017, 174, 4836. [Google Scholar] [CrossRef] [PubMed]
- Xu, Y.; Yu, Q.; Liu, Y. Serum relaxin-2 as a novel biomarker for prostate cancer. Br. J. Biomed. Sci. 2018, 75, 145–148. [Google Scholar] [CrossRef] [PubMed]
- Guo, X.; Liu, Y.; Huang, X.; Wang, Y.; Qu, J.; Lv, Y. Serum relaxin as a diagnostic and prognostic marker in patients with epithelial ovarian cancer. Cancer Biomark. 2017, 21, 81–87. [Google Scholar] [CrossRef] [PubMed]
- Ren, P.; Yu, Z.T.; Xiu, L.; Wang, M.; Liu, H.M. Elevated serum levels of human relaxin-2 in patients with esophageal squamous cell carcinoma. World J. Gastroenterol. 2013, 19, 2412–2418. [Google Scholar] [CrossRef] [PubMed]
- Binder, C.; Simon, A.; Binder, L.; Hagemann, T.; Schulz, M.; Emons, G.; Trümper, L.; Einspanier, A. Elevated concentrations of serum relaxin are associated with metastatic disease in breast cancer patients. Breast Cancer Res. Treat. 2004, 87, 157–166. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Niu, M.; Yang, W.; Zang, L.; Xi, Y. Role of relaxin-2 in human primary osteosarcoma. Cancer Cell Int. 2013, 13, 59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashimoto, C.; Miki, Y.; Tanaka, S.; Takagi, K.; Fue, M.; Doe, Z.; Li, B.; Yaegashi, N.; Suzuki, T.; Ito, K. 17β-Hydroxysteroid Dehydrogenase Type 2 Expression Is Induced by Androgen Signaling in Endometrial Cancer. Int. J. Mol. Sci. 2018, 19, 1139. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, S.; Miki, Y.; Hashimoto, C.; Takagi, K.; Doe, Z.; Li, B.; Yaegashi, N.; Suzuki, T.; Ito, K. The role of 5α-reductase type 1 associated with intratumoral dihydrotestosterone concentrations in human endometrial carcinoma. Mol. Cell Endocrinol. 2015, 401, 56–64. [Google Scholar] [CrossRef] [PubMed]
- Saito, R.; Miki, Y.; Ishida, N.; Inoue, C.; Kobayashi, M.; Hata, S.; Yamada-Okabe, H.; Okada, Y.; Sasano, H. The Significance of MMP-1 in EGFR-TKI-Resistant Lung Adenocarcinoma: Potential for Therapeutic Targeting. Int. J. Mol. Sci. 2018, 19, 609. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Total (n = 80) | RLN2 | ||
|---|---|---|---|---|
| + (n = 54) | − (n = 26) | p-Value | ||
| Age | 57 (29–85) | 57 (30–85) | 53.5 (29–76) | 0.5153 |
| Grade 1 (G1) | 40 | 23 | 17 | |
| 2 (G2) | 25 | 19 | 6 | |
| 3 (G3) | 15 | 13 | 2 | 0.0856 |
| Stage | ||||
| 1 & 2 | 60 | 42 | 18 | |
| 3 & 4 | 20 | 12 | 8 | 0.3081 |
| RXFP1 | ||||
| Positive | 43 | 33 | 10 | |
| Negative | 37 | 21 | 16 | 0.0570 |
| Estrogen receptor | ||||
| Positive | 51 | 32 | 19 | |
| Negative | 29 | 22 | 7 | 0.2225 |
| Progesterone receptor | ||||
| Positive | 24 | 13 | 11 | |
| Negative | 56 | 41 | 15 | 0.1001 |
| Myometrial invasion * | ||||
| Positive | 29 | 19 | 10 | |
| Negative | 48 | 33 | 15 | 0.7691 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fue, M.; Miki, Y.; Takagi, K.; Hashimoto, C.; Yaegashi, N.; Suzuki, T.; Ito, K. Relaxin 2/RXFP1 Signaling Induces Cell Invasion via the β-Catenin Pathway in Endometrial Cancer. Int. J. Mol. Sci. 2018, 19, 2438. https://doi.org/10.3390/ijms19082438
Fue M, Miki Y, Takagi K, Hashimoto C, Yaegashi N, Suzuki T, Ito K. Relaxin 2/RXFP1 Signaling Induces Cell Invasion via the β-Catenin Pathway in Endometrial Cancer. International Journal of Molecular Sciences. 2018; 19(8):2438. https://doi.org/10.3390/ijms19082438
Chicago/Turabian StyleFue, Misaki, Yasuhiro Miki, Kiyoshi Takagi, Chiaki Hashimoto, Nobuo Yaegashi, Takashi Suzuki, and Kiyoshi Ito. 2018. "Relaxin 2/RXFP1 Signaling Induces Cell Invasion via the β-Catenin Pathway in Endometrial Cancer" International Journal of Molecular Sciences 19, no. 8: 2438. https://doi.org/10.3390/ijms19082438