Specific Antibody Fragment Ligand Traps Blocking FGF1 Activity

 ,
,
Abstract
:
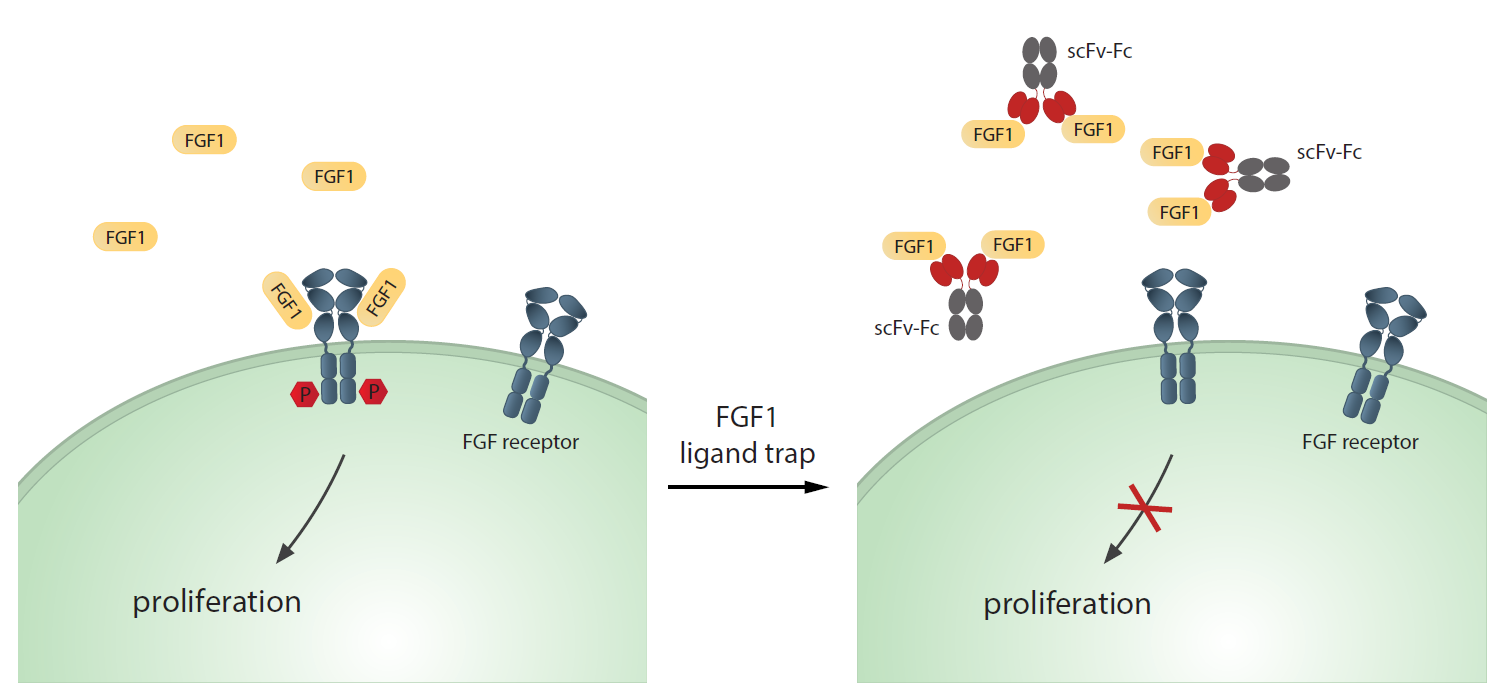
1. Introduction
2. Results
2.1. Selection of Human scFv Antibody Fragments Specific for FGF1
2.2. Purification of the Generated FGF1-Specific scFvs
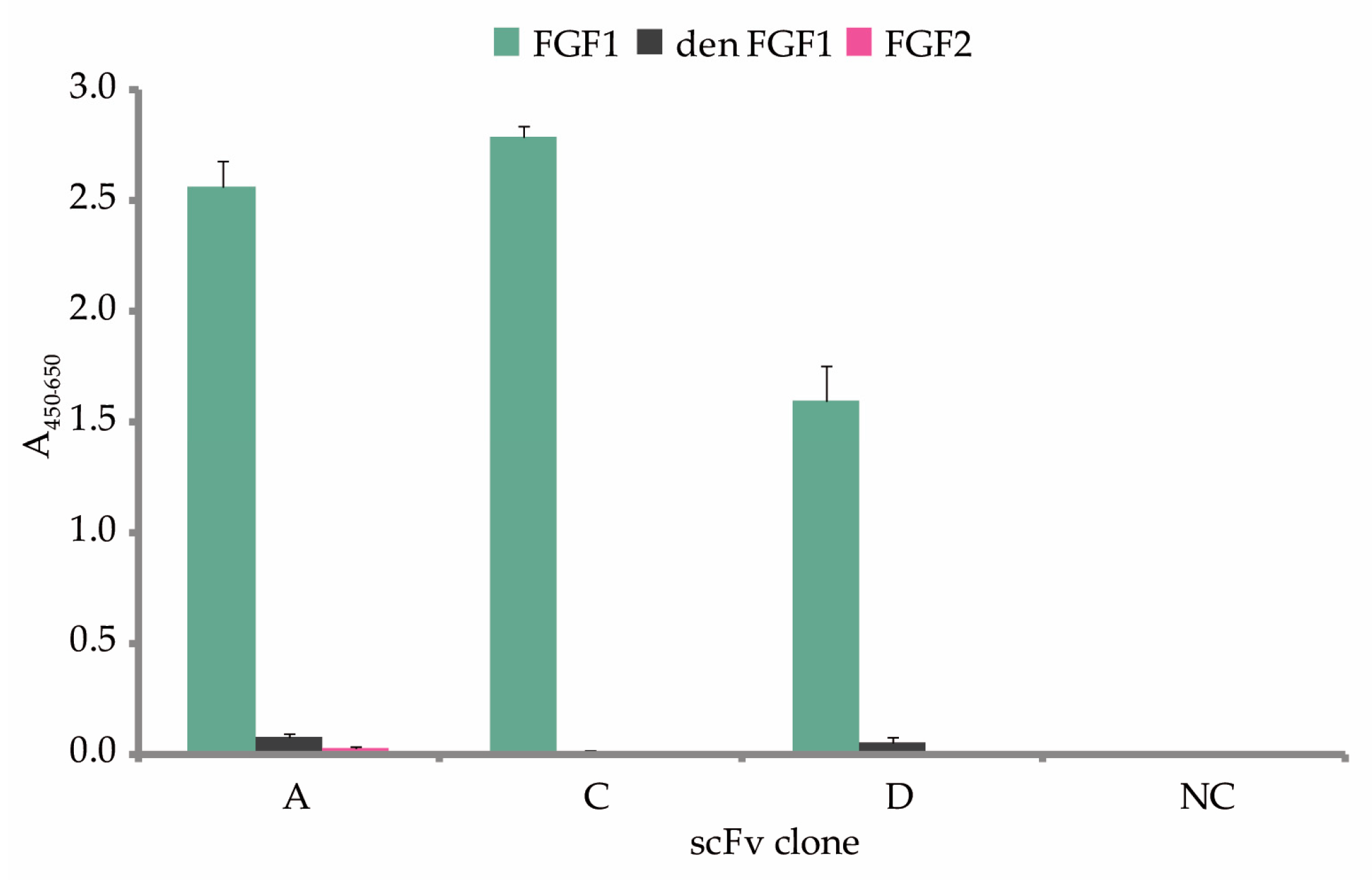
2.3. Characterization, Analysis of Binding to FGF1 and Kinetic Parameters of the Selected scFvs
2.4. Reformatting of the scFv Antibody Fragments to the scFv-Fc Fusions
2.5. Kinetic Parameters of the scFv-Fc Antibody Fragments
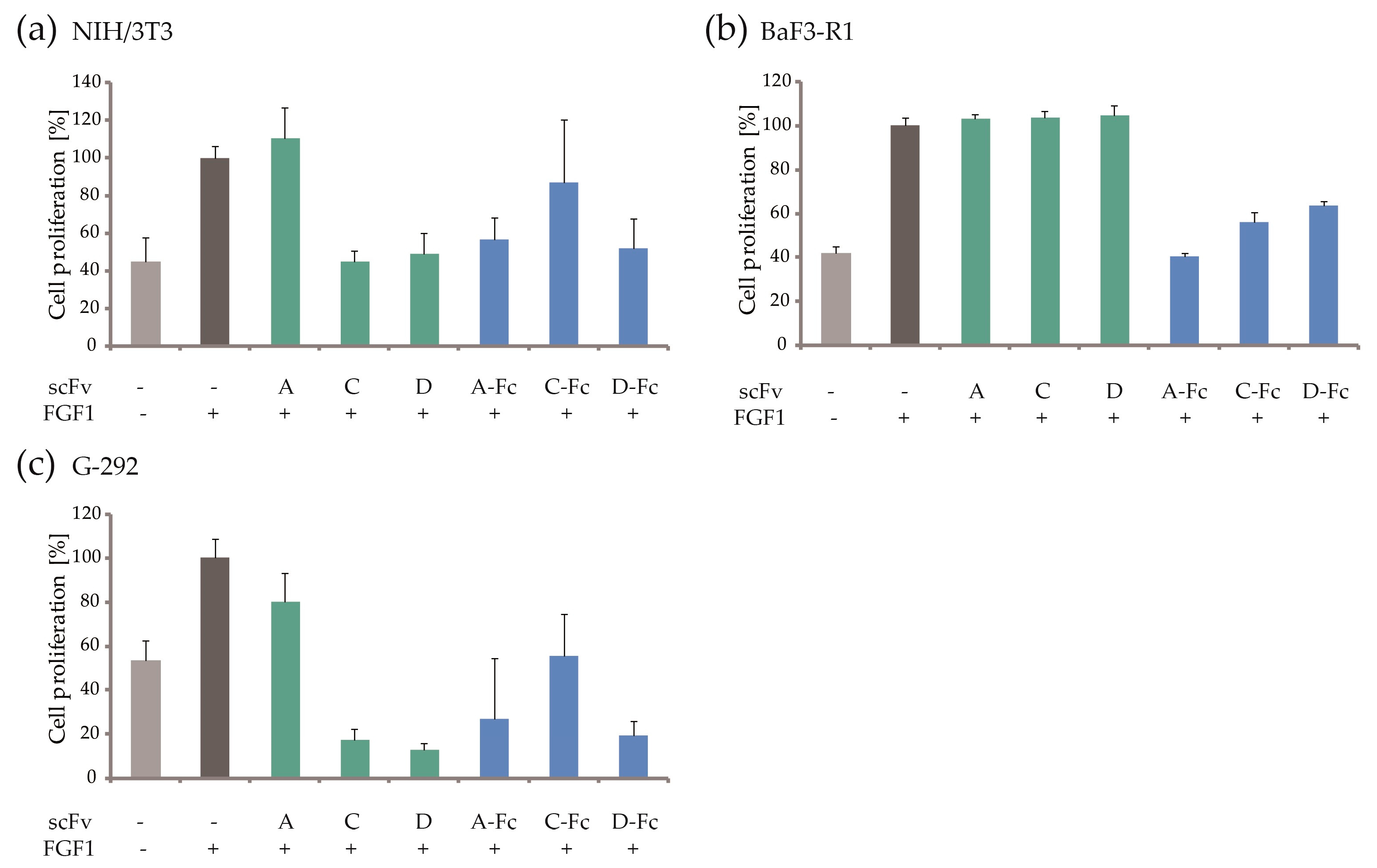
2.6. Blocking of FGF1-dependent Cell Proliferation
3. Discussion
4. Materials and Methods
4.1. Proteins
4.1.1. FGF1 and FGF1 Mutant Y94A/N95A
4.1.2. 15N FGF1
4.1.3. FGF2
4.1.4. ECD_FGFR1-Fc
4.2. Phage Display Selection of scFv Antibody Fragments
4.3. Biolayer Interferometry Screening of scFv Antibody Fragments
4.4. Purification and Analysis of scFv Antibody Fragments
4.5. Nuclear Magnetic Resonance Measurements
4.6. Cross-Reactivity Assay
4.7. Biolayer Interferometry Studies of FGF1 Binding Affinity
4.8. Epitope Binning with Biolayer Interferometry
4.9. scFv-Fc Fusions Preparation, Purification and Analysis of the Proteins
4.10. Cell Proliferation Studies
Supplementary Materials
Author Contributions
Acknowledgments
Conflicts of Interest
Abbreviations
| BLI | Biolayer interferometry |
| CDR | Complementarity determining region |
| CHO | Chinese hamster ovary |
| ECD | Extracellular domain |
| EGFR | Epidermal growth factor receptor |
| ELISA | Enzyme-linked immunosorbent assay |
| Erk1/2 | Extracellular signal-regulated kinases 1/2 |
| Fc | Fragment crystallizable |
| FGF1 | Fibroblast growth factor 1 |
| FGFR | Fibroblast growth factor receptor |
| HMQC | Heteronuclear multiple-quantum correlation |
| HRP | Horseradish peroxidase |
| IB | Immunoblotting |
| KB | Kinetics buffer |
| MALDI | Matrix-assisted laser desorption/ionization |
| MS | Mass spectrometry |
| NMR | Nuclear magnetic resonance |
| RT | Room temperature |
| RTK | Receptor tyrosine kinase |
| SAR | Structure-activity relationships |
| scFv | Single chain Fragment variable |
| SDS-PAGE | Sodium dodecyl sulfate polyacrylamide gel electrophoresis |
| TEA | Triethylamine |
| TKI | Tyrosine kinase inhibitor |
| VEGFR | Vascular endothelial growth factor receptor |
References
- Chin, K.; DeVries, S.; Fridlyand, J.; Spellman, P.T.; Roydasgupta, R.; Kuo, W.L.; Lapuk, A.; Neve, R.M.; Qian, Z.; Ryder, T.; et al. Genomic and transcriptional aberrations linked to breast cancer pathophysiologies. Cancer Cell 2006, 10, 529–541. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gelsi-Boyer, V. Comprehensive Profiling of 8p11-12 Amplification in Breast Cancer. Mol. Cancer Res. 2005, 3, 655–667. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Weiss, J.; Sos, M.L.; Seidel, D.; Peifer, M.; Zander, T.; Heuckmann, J.M.; Ullrich, R.T.; Menon, R.; Maier, S.; Soltermann, A.; et al. Frequent and focal FGFR1 amplification associates with therapeutically tractable FGFR1 dependency in squamous cell lung cancer. Sci. Transl. Med. 2010, 2. [Google Scholar] [CrossRef] [PubMed]
- Tsujimoto, H.; Sugihara, H.; Hagiwara, A.; Hattori, T. Amplification of growth factor receptor genes and DNA ploidy pattern in the progression of gastric cancer. Virchows Arch. 1997, 431, 383–389. [Google Scholar] [CrossRef] [PubMed]
- Hanker, A.B.; Garrett, J.T.; Estrada, M.V.; Moore, P.D.; Ericsson, P.G.; Koch, J.P.; Langley, E.; Singh, S.; Kim, P.S.; Frampton, G.M.; et al. HER2-overexpressing breast cancers amplify FGFR signaling upon acquisition of resistance to dual therapeutic blockade of HER2. Clin. Cancer Res. 2017, 23, 4323–4334. [Google Scholar] [CrossRef] [PubMed]
- Hallinan, N.; Finn, S.; Cuffe, S.; Rafee, S.; O’Byrne, K.; Gately, K. Targeting the fibroblast growth factor receptor family in cancer. Cancer Treat. Rev. 2016, 46, 51–62. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eswarakumar, V.P.; Lax, I.; Schlessinger, J. Cellular signaling by fibroblast growth factor receptors. Cytokine Growth Factor Rev. 2005, 16, 139–149. [Google Scholar] [CrossRef] [PubMed]
- Presta, M.; Dell’Era, P.; Mitola, S.; Moroni, E.; Ronca, R.; Rusnati, M. Fibroblast growth factor/fibroblast growth factor receptor system in angiogenesis. Cytokine Growth Factor Rev. 2005, 16, 159–178. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Katoh, M. Therapeutics Targeting FGF Signaling Network in Human Diseases. Trends Pharmacol. Sci. 2016, 37, 1081–1096. [Google Scholar] [CrossRef] [PubMed]
- Hudis, C.A. Trastuzumab—Mechanism of Action and Use in Clinical Practice. N. Engl. J. Med. 2007, 357, 39–51. [Google Scholar] [CrossRef] [PubMed]
- Porta, R.; Borea, R.; Coelho, A.; Khan, S.; Araújo, A.; Reclusa, P.; Franchina, T.; Van Der Steen, N.; van Dam, P.; Ferri, J.; et al. FGFR a promising druggable target in cancer: Molecular biology and new drugs. Crit. Rev. Oncol. Hematol. 2017, 113, 256–267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, A.; Meetze, K.; Vo, N.Y.; Kollipara, S.; Mazsa, E.K.; Winston, W.M.; Weiler, S.; Poling, L.L.; Chen, T.; Ismail, N.S.; et al. GP369, an FGFR2-IIIb-specific antibody, exhibits potent antitumor activity against human cancers driven by activated FGFR2 signaling. Cancer Res. 2010, 70, 7630–7639. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Wang, L.; Park, H.; Chhim, S.; Tanphanich, M.; Yashiro, M.; Kim, K.J. Monoclonal Antibodies to Fibroblast Growth Factor Receptor 2 Effectively Inhibit Growth of Gastric Tumor Xenografts. Clin. Cancer Res. 2010, 16, 5750–5758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inokuchi, M.; Fujimori, Y.; Otsuki, S.; Sato, Y.; Nakagawa, M.; Kojima, K. Therapeutic Targeting of Fibroblast Growth Factor Receptors in Gastric Cancer. Gastroenterol. Res. Pract. 2015, 2015. [Google Scholar] [CrossRef] [PubMed]
- ODonnell, P.; Goldman, J.W.; Gordon, M.S.; Shih, K.; Choi, Y.J.; Lu, D.; Kabbarah, O.; Ho, W.; Rooney, I.; Lam, E.T. 621 A Phase I Dose-escalation Study of MFGR1877S, a Human Monoclonal Anti-fibroblast Growth Factor Receptor 3 (FGFR3) Antibody, in Patients (pts) with Advanced Solid Tumors. Eur. J. Cancer 2012, 48, 191–192. [Google Scholar] [CrossRef]
- Tolcher, A.W.; Papadopoulos, K.P.; Patnaik, A.; Wilson, K.; Thayer, S.; Zanghi, J.; Gemo, A.T.; Kavanaugh, W.M.; Keer, H.N.; LoRusso, P.M. A phase I, first in human study of FP-1039 (GSK3052230), a novel FGF ligand trap, in patients with advanced solid tumors. Ann. Oncol. 2016, 27, 526–532. [Google Scholar] [CrossRef] [PubMed]
- Li, D.; Wei, X.; Xie, K.; Chen, K.; Li, J.; Fang, J. A novel decoy receptor fusion protein for FGF-2 potently inhibits tumour growth. Br. J. Cancer 2014, 111, 68–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Taraboletti, G.; Rusnati, M.; Ragona, L.; Colombo, G. Targeting tumor angiogenesis with TSP-1-based compounds: Rational design of antiangiogenic mimetics of endogenous inhibitors. Oncotarget 2010, 1, 662–673. [Google Scholar] [PubMed]
- Ronca, R.; Giacomini, A.; Rusnati, M.; Presta, M. The potential of fibroblast growth factor/fibroblast growth factor receptor signaling as a therapeutic target in tumor angiogenesis. Expert Opin. Ther. Targets 2015, 19, 1361–1377. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Park, H.; Chhim, S.; Ding, Y.; Jiang, W.; Queen, C.; Kim, K.J. A Novel Monoclonal Antibody to Fibroblast Growth Factor 2 Effectively Inhibits Growth of Hepatocellular Carcinoma Xenografts. Mol. Cancer Ther. 2012, 11, 864–872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tao, J.; Xiang, J.J.; Li, D.; Deng, N.; Wang, H.; Gong, Y.P. Selection and characterization of a human neutralizing antibody to human fibroblast growth factor-2. Biochem. Biophys. Res. Commun. 2010, 394, 767–773. [Google Scholar] [CrossRef] [PubMed]
- Cai, Y.; Yao, S.; Zhong, J.; Zhang, J.; Jiang, H.; Deng, Y.; Deng, N. Inhibition activity of a disulfide-stabilized diabody against basic fibroblast growth factor in lung cancer. Oncotarget 2017, 8, 20187–20197. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, A.; Furuya, A.; Yamasaki, M.; Hanai, N.; Kuriki, K.; Kamiakito, T.; Kobayashi, Y.; Yoshida, H.; Koike, M.; Fukayama, M. High frequency of fibroblast growth factor (FGF) 8 expression in clinical prostate cancers and breast tissues, immunohistochemically demonstrated by a newly established neutralizing monoclonal antibody against FGF 8. Cancer Res. 1998, 58, 2053–2056. [Google Scholar] [PubMed]
- Shi, H.-L.; Yang, T.; Deffar, K.; Dong, C.-G.; Liu, J.-Y.; Fu, C.-L.; Zheng, D.-X.; Qin, B.; Wang, J.-J.; Wang, X.-Z.; et al. A novel single-chain variable fragment antibody against FGF-1 inhibits the growth of breast carcinoma cells by blocking the intracrine pathway of FGF-1. IUBMB Life 2011, 63, 129–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aono, Y.; Hasegawa, H.; Yamazaki, Y.; Shimada, T.; Fujita, T.; Yamashita, T.; Fukumoto, S. Anti-FGF-23 neutralizing antibodies ameliorate muscle weakness and decreased spontaneous movement of Hyp mice. J. Bone Miner. Res. 2011, 26, 803–810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrzewska, M.; Krowarsch, D.; Wiedlocha, A.; Olsnes, S.; Otlewski, J. Structural Requirements of FGF-1 for Receptor Binding and Translocation into Cells. Biochemistry 2006, 45, 15338–15348. [Google Scholar] [CrossRef] [PubMed]
- Powers, C.J.; MclEskey, S.W.; Wellstein, A. Fibroblast growth factors, their receptors and signaling. Endocr. Relat. Cancer 2000, 7, 165–197. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beenken, A.; Mohammadi, M. The FGF family: Biology, pathophysiology and therapy. Nat. Rev. Drug Discov. 2009, 8, 235–253. [Google Scholar] [CrossRef] [PubMed]
- Turner, N.; Grose, R. Fibroblast growth factor signalling: From development to cancer. Nat. Rev. Cancer 2010, 10, 116–129. [Google Scholar] [CrossRef] [PubMed]
- Chae, Y.K.; Ranganath, K.; Hammerman, P.S.; Siddik, Z.H.; Mohindra, N.; Kalya, A.; Matsangou, M.; Costa, R.; Carneiro, B.; Villaflor, V.M.; et al. Inhibition of the fibroblast growth factor receptor (FGFR) pathway: The current landscape and barriers to clinical application. Oncotarget 2017, 8, 16052–16074. [Google Scholar] [CrossRef] [PubMed]
- Babina, I.S.; Turner, N.C. Advances and challenges in targeting FGFR signalling in cancer. Nat. Rev. Cancer 2017, 17, 318–332. [Google Scholar] [CrossRef] [PubMed]
- Liang, G.; Liu, Z.; Wu, J.; Cai, Y.; Li, X. Anticancer molecules targeting fibroblast growth factor receptors. Trends Pharmacol. Sci. 2012, 33, 531–541. [Google Scholar] [CrossRef] [PubMed]
- Sommer, A.; Kopitz, C.; Schatz, C.A.; Nising, C.F.; Mahlert, C.; Lerchen, H.G.; Stelte-Ludwig, B.; Hammer, S.; Greven, S.; Schuhmacher, J.; et al. Preclinical efficacy of the auristatin-based antibody-drug conjugate BAY 1187982 for the treatment of FGFR2-positive solid tumors. Cancer Res. 2016, 76, 6331–6339. [Google Scholar] [CrossRef] [PubMed]
- Giacomini, A.; Chiodelli, P.; Matarazzo, S.; Rusnati, M.; Presta, M.; Ronca, R. Blocking the FGF/FGFR system as a “two-compartment” antiangiogenic/antitumor approach in cancer therapy. Pharmacol. Res. 2016, 107, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Harding, T.C.; Long, L.; Palencia, S.; Zhang, H.; Sadra, A.; Hestir, K.; Patil, N.; Levin, A.; Hsu, A.W.; Charych, D.; et al. Blockade of Nonhormonal Fibroblast Growth Factors by FP-1039 Inhibits Growth of Multiple Types of Cancer. Sci. Transl. Med. 2013, 5, 178ra39. [Google Scholar] [CrossRef] [PubMed]
- Rusnati, M.; Oreste, P.; Zoppetti, G.; Presta, M. Biotechnological engineering of heparin/heparan sulphate: A novel area of multi-target drug discovery. Curr. Pharm. Des. 2005, 11, 2489–2499. [Google Scholar] [CrossRef] [PubMed]
- Blackwell, C.; Sherk, C.; Fricko, M.; Ganji, G.; Barnette, M.; Hoang, B.; Tunstead, J.; Skedzielewski, T.; Alsaid, H.; Jucker, B.M.; et al. Inhibition of FGF/FGFR autocrine signaling in mesothelioma with the FGF ligand trap, FP-1039/GSK3052230. Oncotarget 2016, 7, 39861–39871. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- He, X.-X.; Du, S.; Gao, S.-Q.; Chen, J.-Y.; Cao, R.-J.; Xing, Z.-K.; Kazim, A.R.S.; Yu, H.-L.; Zheng, Q.-C.; Zhu, X.-J. Humanization of fibroblast growth factor 1 single-chain antibody and validation for its antitumorigenic efficacy in breast cancer and glioma cells. J. Cell. Mol. Med. 2018, 22, 3259–3263. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, H.; Fu, C.; Wang, W.; Li, Y.; Du, S.; Cao, R.; Chen, J.; Sun, D.; Zhang, Z.; Wang, X.; et al. The FGF-1-specific single-chain antibody scFv1C9 effectively inhibits breast cancer tumour growth and metastasis. J. Cell. Mol. Med. 2014, 18, 2061–2070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Holzmann, K.; Grunt, T.; Heinzle, C.; Sampl, S.; Steinhoff, H.; Reichmann, N.; Kleiter, M.; Hauck, M.; Marian, B. Alternative splicing of fibroblast growth factor receptor IgIII loops in cancer. J. Nucleic Acids 2012, 2012. [Google Scholar] [CrossRef] [PubMed]
- Lee, C.M.Y.; Iorno, N.; Sierro, F.; Christ, D. Selection of human antibody fragments by phage display. Nat. Protoc. 2007, 2, 3001–3008. [Google Scholar] [CrossRef] [PubMed]
- Vaks, L.; Benhar, I. Production of stabilized scFv antibody fragments in the E. coli bacterial cytoplasm. Methods Mol. Biol. 2014, 1060, 171–184. [Google Scholar] [CrossRef] [PubMed]
- Shuker, S.B.; Hajduk, P.J.; Meadows, R.P.; Fesik, S.W. Discovering high-affinity ligands for proteins: SAR by NMR. Science 1996, 274, 1531–1534. [Google Scholar] [CrossRef] [PubMed]
- Barile, E.; Pellecchia, M. NMR-based approaches for the identification and optimization of inhibitors of protein-protein interactions. Chem. Rev. 2014, 114, 4749–4763. [Google Scholar] [CrossRef] [PubMed]
- Singh, M.; Krajewski, M.; Mikolajka, A.; Holak, T.A. Molecular determinants for the complex formation between the retinoblastoma protein and LXCXE sequences. J. Biol. Chem. 2005, 280, 37868–37876. [Google Scholar] [CrossRef] [PubMed]
- Williamson, M.P. Using chemical shift perturbation to characterise ligand binding. Prog. Nucl. Magn. Reson. Spectrosc. 2013, 73, 1–16. [Google Scholar] [CrossRef] [PubMed]
- Bujak, E.; Matasci, M.; Neri, D.; Wulhfard, S. Reformatting of scFv Antibodies into the scFv-Fc Format and Their Downstream Purification. In Methods in Molecular Biology; Springer: New York, USA, 2014; Volume 1131, pp. 315–334. ISBN 9781627039918. [Google Scholar]
- Ornitz, D.M.; Xu, J.; Colvin, J.S.; McEwen, D.G.; MacArthur, C.A.; Coulier, F.; Gao, G.; Goldfarb, M. Receptor specificity of the fibroblast growth factor family. J. Biol. Chem. 1996, 271, 15292–15297. [Google Scholar] [CrossRef] [PubMed]
- Zakrzewska, M.; Krowarsch, D.; Wiedlocha, A.; Otlewski, J. Design of fully active FGF-1 variants with increased stability. Protein Eng. Des. Sel. 2004, 17, 603–611. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zakrzewska, M.; Krowarsch, D.; Wiedlocha, A.; Olsnes, S.; Otlewski, J. Highly Stable Mutants of Human Fibroblast Growth Factor-1 Exhibit Prolonged Biological Action. J. Mol. Biol. 2005, 352, 860–875. [Google Scholar] [CrossRef] [PubMed]
- Swiderska, K.W.; Szlachcic, A.; Czyrek, A.; Zakrzewska, M.; Otlewski, J. Site-specific conjugation of fibroblast growth factor 2 (FGF2) based on incorporation of alkyne-reactive unnatural amino acid. Bioorg. Med. Chem. 2017, 25, 3685–3693. [Google Scholar] [CrossRef] [PubMed]
- Sokolowska-Wedzina, A.; Borek, A.; Chudzian, J.; Jakimowicz, P.; Zakrzewska, M.; Otlewski, J. Efficient production and purification of extracellular domain of human FGFR-Fc fusion proteins from Chinese hamster ovary cells. Protein Expr. Purif. 2014, 99, 50–57. [Google Scholar] [CrossRef] [PubMed]
- Viti, F.; Nilsson, F.; Demartis, S.; Huber, A.; Neri, D. Design and use of phage display libraries for the selection of antibodies and enzymes. Methods Enzymol. 2000, 326, 480–505. [Google Scholar] [CrossRef] [PubMed]
- Sokolowska-Wedzina, A.; Chodaczek, G.; Chudzian, J.; Borek, A.; Zakrzewska, M.; Otlewski, J. High-Affinity Internalizing Human scFv-Fc Antibody for Targeting FGFR1-Overexpressing Lung Cancer. Mol. Cancer Res. 2017, 15, 1040–1051. [Google Scholar] [CrossRef] [PubMed]
- Schanda, P.; Brutscher, B. Very fast two-dimensional NMR spectroscopy for real-time investigation of dynamic events in proteins on the time scale of seconds. J. Am. Chem. Soc. 2005, 127, 8014–8015. [Google Scholar] [CrossRef] [PubMed]
- Flores-Flores, C.; Nissim, A.; Shochat, S.; Soreq, H. Development of human antibody fragments directed towards synaptic acetylcholinesterase using a semi-synthetic phage display library. J. Neural Transm. Suppl. 2002, 165–179. [Google Scholar]
- Nissim, A.; Hoogenboom, H.R.; Tomlinson, I.M.; Flynn, G.; Midgley, C.; Lane, D.; Winter, G. Antibody fragments from a “single pot” phage display library as immunochemical reagents. EMBO J. 1994, 13, 692–698. [Google Scholar] [PubMed]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein | KD1 [M] | ka1 [M−1s−1] | kd1 [s−1] | KD2 [M] | ka2 [M−1s−1] | kd2 [s−1] |
|---|---|---|---|---|---|---|
| scFvA | 1.41 × 10−6 | 2.98 × 104 | 8.32 × 10−2 | 3.32 × 10−5 | 1.35 × 101 | 4.49 × 10−4 |
| scFvC | 9.21 × 10−6 | 9.60 × 103 | 4.19 × 10−2 | 2.05 × 10−6 | 2.13 × 102 | 4.36 × 10−4 |
| scFvD | 1.21 × 10−6 | 2.14 × 104 | 2.58 × 10−2 | 1.35 × 10−5 | 6.93 × 101 | 1.35 × 10−5 |
| Protein | KD1 [M] | ka1 [M−1s−1] | kd1 [s−1] | KD2 [M] | ka2 [M−1s−1] | kd2 [s−1] |
|---|---|---|---|---|---|---|
| scFvA-Fc | 4.24 × 10−7 | 8.81 × 104 | 3.72 × 10−2 | 7.27 × 10−6 | 1.64 × 102 | 1.19 × 10−3 |
| scFvC-Fc | 5.22 × 10−7 | 7.24 × 104 | 3.78 × 10−2 | 8.81 × 10−6 | 8.12 × 101 | 7.16 × 10−4 |
| scFvD-Fc | 3.46 × 10−7 | 1.32 × 105 | 4.56 × 10−2 | 6.52 × 10−5 | 5.13 × 101 | 3.34 × 10−3 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chudzian, J.; Szlachcic, A.; Zakrzewska, M.; Czub, M.; Pustula, M.; Holak, T.A.; Otlewski, J. Specific Antibody Fragment Ligand Traps Blocking FGF1 Activity. Int. J. Mol. Sci. 2018, 19, 2470. https://doi.org/10.3390/ijms19092470
Chudzian J, Szlachcic A, Zakrzewska M, Czub M, Pustula M, Holak TA, Otlewski J. Specific Antibody Fragment Ligand Traps Blocking FGF1 Activity. International Journal of Molecular Sciences. 2018; 19(9):2470. https://doi.org/10.3390/ijms19092470
Chicago/Turabian StyleChudzian, Julia, Anna Szlachcic, Malgorzata Zakrzewska, Miroslawa Czub, Marcin Pustula, Tad A. Holak, and Jacek Otlewski. 2018. "Specific Antibody Fragment Ligand Traps Blocking FGF1 Activity" International Journal of Molecular Sciences 19, no. 9: 2470. https://doi.org/10.3390/ijms19092470