1. Introduction
Approximately 30% of astronauts on short-term (~2 week) Space Shuttle flights and 60% on long-duration (~6 month) missions to the International Space Station (ISS) are reported to have experienced some impairment in distant or near visual acuity [
1]. Previous studies of rodents also show that environmental conditions associated with spaceflight and simulated microgravity induce alterations in retinal structure and function [
2,
3,
4].
Retinal damage and degeneration can occur as a result of multiple factors, including aging, ischemia, fluctuation in oxygen tension, oxidative stress, and increased intraocular pressure. Our previous data from Space Shuttle studies showed a significant increase of apoptosis in retinal cells of flight mice compared to ground controls [
2], which could lead to morphologic changes and impairment of retinal function. However, the pathophysiological consequences and cellular mechanisms of stress stimuli, especially those associated with a spaceflight environment, in facilitating retinal damage are less studied and remain unclear.
Data on the response of biochemical systems to altered gravity is very limited in animal studies [
5]. Some lines of evidence suggest that one of the mechanisms involved in response to spaceflight, including changes in the gravity vector, is likely due to oxidative stress [
2,
6,
7]. Studies have shown that exposure to microgravity during spaceflights is associated with elevations in oxidative stress and production of lipid peroxidation in both humans and rodents [
8,
9]. However, our knowledge about the duration or dose of microgravity exposure needed to induce oxidative stress or alterations in retinal structure and function is extremely limited. For example, one study has shown that even transient exposure to microgravity produced by parabolic flight can cause alterations in retinal vasculature to occur [
1]. One means to address the impact of microgravity conditions is through the application of artificial gravity (AG) during spaceflight.
Creating AG by centrifugal force makes it possible to have a partial gravity or Earth-like gravity (1
g) control in space, where all other environmental conditions associated with spaceflight are present. A newly developed mouse habitat cage unit (HCU) by Japan Aerospace Exploration Agency (JAXA) has been installed in the Centrifuge-equipped Biological Experiment Facility (CBEF) on board the ISS [
10]. This rodent centrifugation capability provides opportunities to compare the effectiveness of AG prescription under spaceflight conditions. Since the HCU can reproduce Earth-like gravity, it can be used to test the effectiveness of AG to mitigate the effects of microgravity on physiological systems [
11].
The goal of the present study was to investigate the effects of spaceflight on oxidative stress and apoptosis in retinal endothelial cells and to identify spaceflight-induced changes in protein expression profiles in mouse ocular tissue. Additionally, we sought to determine whether the application of 1 g AG during spaceflight could mitigate any detrimental effects of microgravity on the retina. We hypothesized that spaceflight would induce elevations in oxidative stress and apoptosis in retinal endothelial cells, as well as alter ocular proteins associated with apoptosis, cell repair, inflammation and metabolic function. We further hypothesized that the application of 1 g AG would mitigate these changes.
3. Discussion
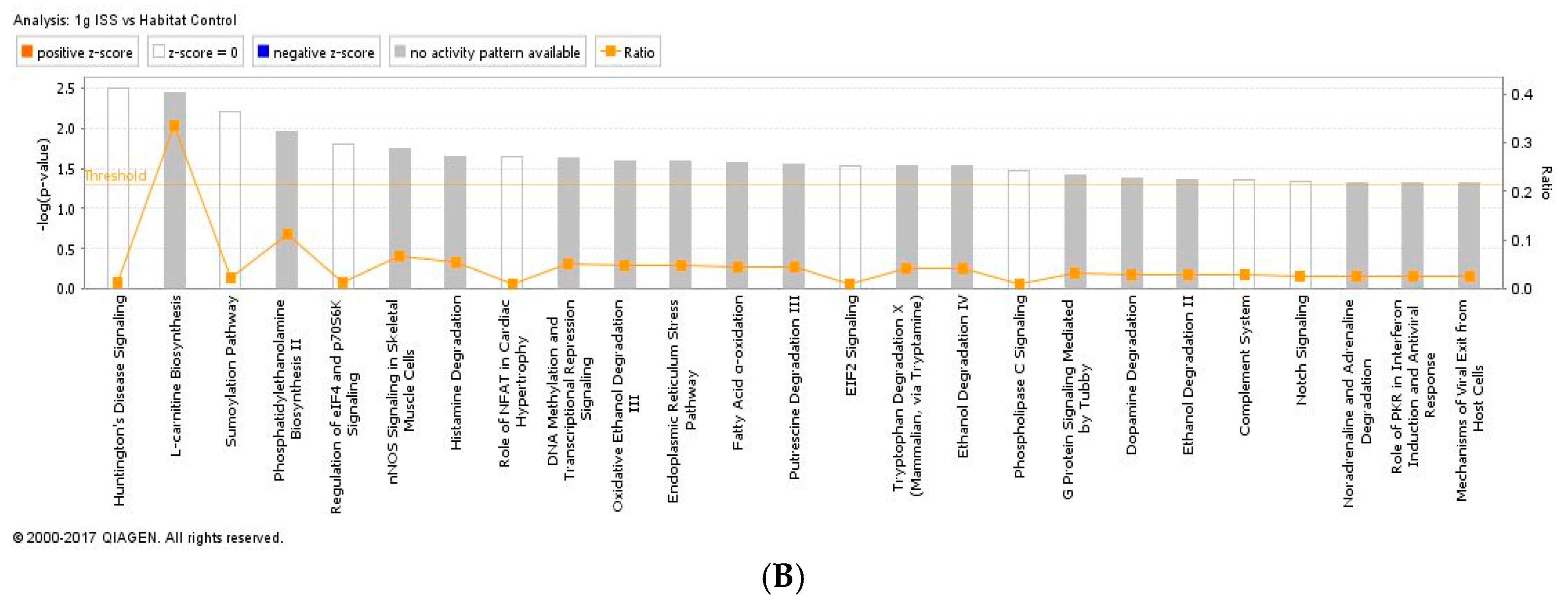
The current study demonstrates that spaceflight induces apoptosis in retinal vascular endothelial cells. We also identified spaceflight-induced changes in proteomic profiles and pathways in the ocular tissue. The results indicate that spaceflight induces changes in neuronal structure, cellular organization, mitochondrial function and oxidative phosphorylation and inflammation which, in turn, may lead to tissue injury and late neurodegeneration. This study is the first to investigate the role of AG provided by centrifugation during spaceflight as a countermeasure for mitigating putative effects of microgravity on ocular structure and function.
In our study, several oxidative stress-related signaling pathways are significantly altered in the µg group, which includes fatty acid β-oxidation I, fatty acid activation and α-adrenergic pathway. However, we did not find a significant change in an oxidative stress marker for lipid peroxidation, 4-HNE when measured two days after splash-down. In our previous space shuttle Atlantis (STS-135) study, the level of HNE protein was significantly elevated in the retina after spaceflight compared to controls [
2]. This is not surprising as these two flight conditions are different. Female C57BL/6 mice were flown in the STS-135 for 13-day mission, and within 3–5 h of landing, eyes were removed for analysis. In the current study, male mice were flown to the ISS for a 35-day mission, and the mice were euthanized at about 40 h after splashing down. Differences in gender, mission duration and recovery time might contribute to their dynamic stress response. The level of oxidative stress can be affected by their stress reaction to a prolonged flight in space and in the period of re-adaptation to the normal gravity after landing [
15]. Another possible explanation for the lack of significant change in 4-HNE in the current study is that spaceflight might elicit protective effects in the central nervous system (CNS) by inhibiting some gravity-initiated stress signaling pathway.
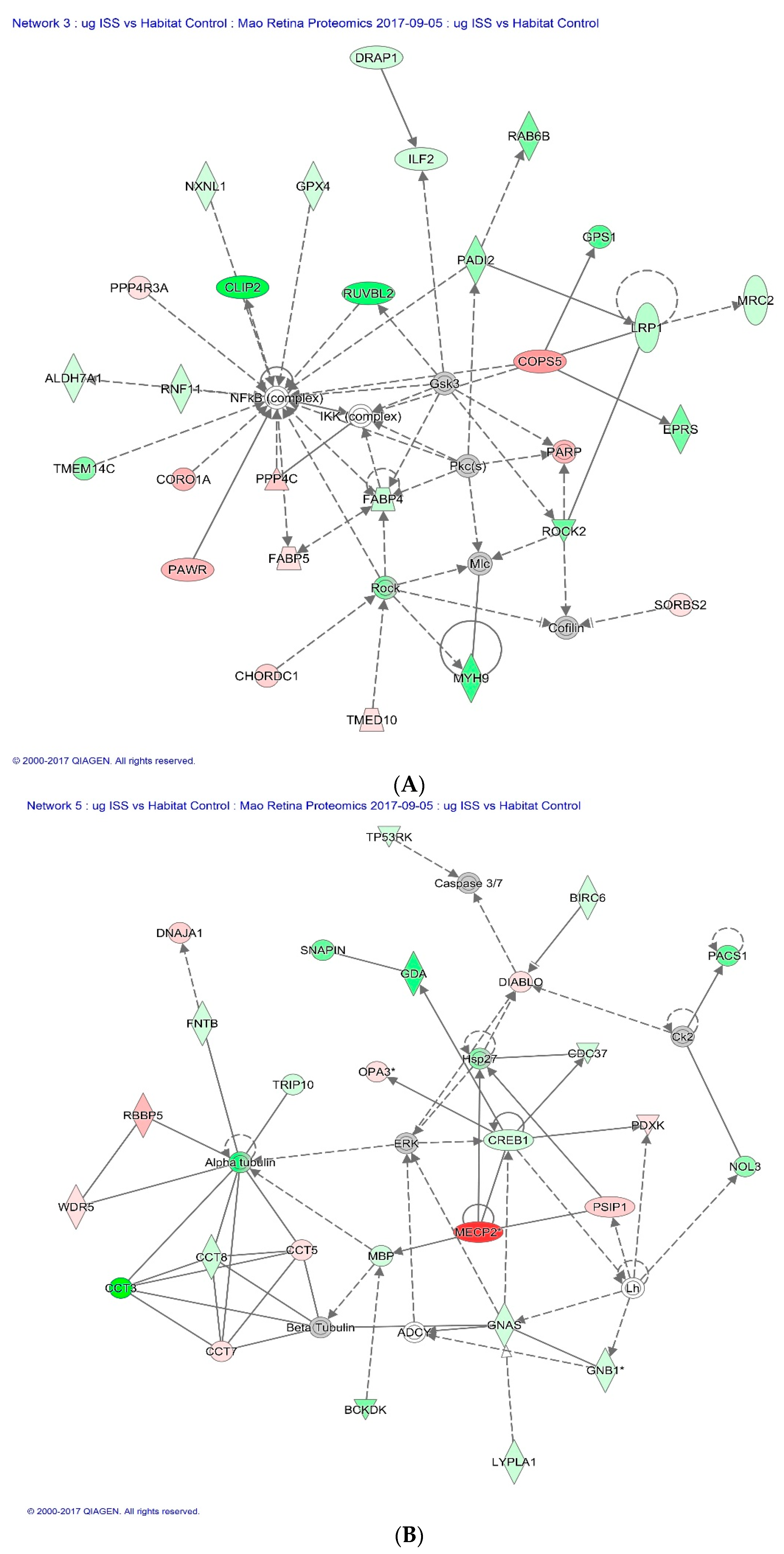
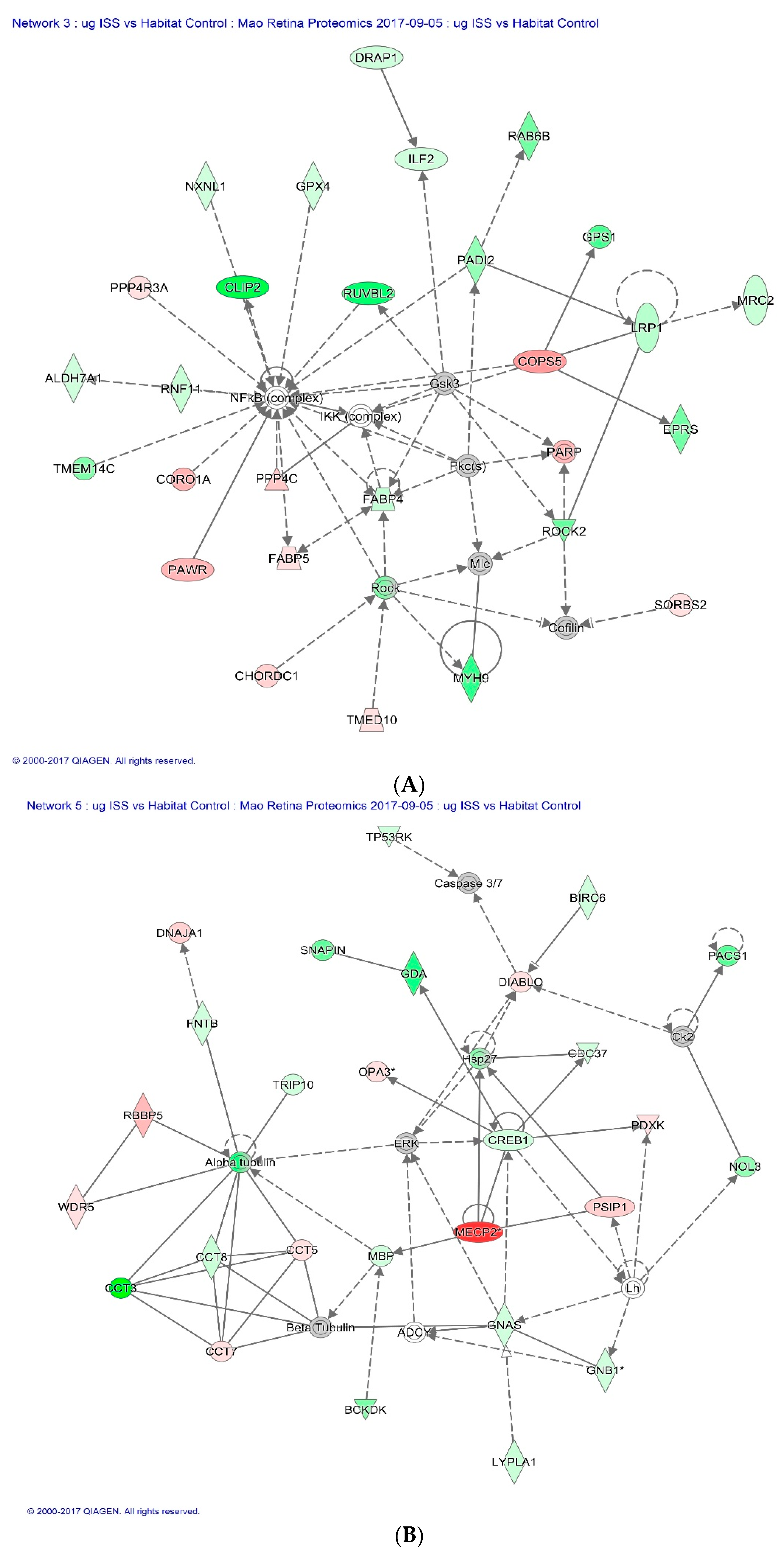
Of all the proteins significantly altered by spaceflight, the Cap-Gly domain containing linker protein 2 (CLIP2), also known as CLIP-115, was significantly down-regulated in both µg and µg + 1
g groups. This protein is found predominantly in the CNS, where it likely plays a role in the normal structure and function of nerve cells. Within cells, this protein is thought to regulate cell cytoskeleton function that helps to determine cell shape, size, and movement [
16,
17,
18,
19,
20].
Figure 6A illustrates CLIP2 in the network of proteins responsible for cellular assembly and organization, cell signaling and interaction as defined by IPA in response to spaceflight.
Methyl-CpG-binding protein 2 (MECP2) had the highest fold changes in flight mice (µg group) compared to HC. This protein binds to methylated DNA and regulates chromatin structure and transcription [
21,
22]. MECP2 is most abundantly produced in the brain where it is expressed primarily in post-mitotic neurons [
23]. Previous work has also shown MECP2 mediated epigenetic regulation of senescent endothelial progenitor cells and promoted apoptosis [
24]. Furthermore, a recent study provides evidence that elevated MECP2 in mice causes neurodegeneration [
25].
Figure 6B shows the regulative role of MECP2 in a network of proteins that involves cell death and survival.
The data from this study indicates that adding 1
g on the ISS is effective in reducing the endothelial cell damage and increasing cellular organization and function compared to the µg group. Another study of the same animals used in our present study also showed that 1
g AG served to maintain femur bone density and soleus/gastrocnemius muscle mass similar to that in the ground control group, whereas significant decreases in bone density and muscle mass occurred in the flight mice without centrifuge [
10]. These studies provide evidence that in-flight AG can mitigate some of the effects of weightlessness during spaceflight. Further investigation will be needed to define the relationship between gravitational dose/time and physiological response by assessing retinal physiological endpoints and function. More rodent centrifugation studies on board the ISS are also needed to provide comprehensive information to compare the effectiveness of the AG prescription in other physiological systems during weightless conditions.
4. Material and Methods
4.1. Spaceflight and Mouse Condition
Twelve male 9-week old C57BL/6 male mice, obtained from a US breeding colony, were launched 18 July 2016, at the Kennedy Space Center (KSC) on a SpaceX-9 rocket for the 35-day MHU-1 mission to the ISS. The animals were housed in the mouse HCU located in the JAXA “Kibo” facility on the ISS. The 12 flight mice were subdivided into two groups. The first group of flight mice (
n = 6) were exposed to ambient microgravity conditions (µg group), while the second group of flight mice (
n = 6) were exposed to continuous artificial Earth gravity (µg + 1
g group) while they were in the HCU. AG was achieved through the use of a short-arm centrifuge for the duration of their stay on the ISS. The flight mice were then returned live to Earth and splashed down in the Pacific Ocean on 26 August 2016. It took approximately 40 h for the mice to be recovered in the Pacific Ocean, brought to shore and transported to the testing and processing laboratory located in San Diego, California on 28 August 2016. The spaceflight mice were then euthanized and their eyes were removed and prepared for analysis. Ground control mouse studies were completed in Japan after the return of the flight mice. Control mice (habitat controls,
n = 6; vivarium controls,
n = 6) were acquired on 31 August 2016 from a breeding colony in Japan and shipped to the JAXA animal facility in Tsukuba, Japan. HC mice were acclimated to the water lixit system and the same special food bar diet as the space flown mice were fed. They were first housed in the transportation cage unit (TCU) to simulate launch and flight to the ISS, and then placed in the HCUs to simulate the housing conditions experienced by µg mice on the ISS. They were again placed in the TCU to simulate the return to Earth flight. The control mouse dissections took place on 3 November 2016. Control mouse eye tissue was then shipped to the US for analysis. All mice received the same ad libitum access to food and water. A detailed description of the flight schedule and mouse information has been previously reported [
11].
The Loma Linda University (LLU) Institutional Animal Care and Use Committee (IACUC) was consulted, but no protocol was required since only archived frozen and fixed tissues were obtained after euthanasia. However, animal experiments were approved by the Institutional Animal Care and Use Committee of University of Tsukuba (No. 16-048), Explora Biolabs (Study Number: EB15-010) on 23 May 2016, JAXA (Protocol Number: 016-014B) on 2 June 2016 and NASA (Protocol Number: NAS-15-004-Y1) on 31 May 2016, and the research with vertebrate animals are done in strict accordance with guidelines and applicable laws in Japan and the United States of America.
4.2. Dissecting and Preservation of Mouse Eyes after Spaceflight
Within 2 days of splashing down, mice were euthanized and their eyes were harvested. The posterior half of the right eyecup containing the retina layer, optic disc and a short segment of the optic nerve was placed individually in sterile cryovials, snap frozen in liquid nitrogen and kept at −80 °C prior to use. The left eyes were fixed in 4% paraformaldehyde in phosphate buffered saline (PBS) for immunohistochemistry (IHC) assays.
4.3. Immunohistochemistry Assays and Histology
Six µm sections were cut through each eye, and sections were roughly 100 µm apart providing 10 sections per eye for analysis. To characterize apoptosis, 5 sections were subjected to a terminal deoxynucleotidyltransferase dUTP nick-end labeling (TUNEL) staining. Paraffin-embedded sections were deparaffinized in Histo-Clear, then permeabilized in proteinase K solution. Stained sections were evaluated using the DeadEnd™ Fluorometric TUNEL system kit (catalog no. G3250, Promega Corp., Madison, WI, USA). The same sections were then incubated with DyLight 594 Lycopersicon esculantum-Lectin (catalog no: DL-11721, Vector Laboratories, Burlingame, CA, USA) at a 1:100 dilution for 30 min at room temperature to stain the endothelium. Nuclei were counterstained with diamidino-2-phenylindole (DAPI, blue). Sections were examined using a BZ-X710 All-in-One inverted fluorescence microscope with structural illumination (Keyence Corp., Elmwood Park, NJ, USA). TUNEL-positive cells were identified by green fluorescence, vascular endothelium were identified with red fluorescence. TUNEL-positive cells that were laid within red lectin-labeled endothelium were identified as TUNEL-positive endothelial cells.
For quantitative analysis, the total number of TUNEL-positive endothelial cells in the retina were determined in 5 sections of each eye. The area of the retina was measured on digital microphotographs using ImageJ counting plugin 1.41 software (National Institutes of Health, Bethesda, MD, USA;
http://rsbweb.nih.gov/ij/), and the density profiles were expressed as mean number of apoptotic positive cells/mm
2. The mean of the density profile measurements across 5 retina sections per eye was used as a single experimental value.
Immunofluorescence staining for an oxidative damage marker on ocular sections was also performed using an anti-4-HNE antibody. Five sections were incubated with the anti-4-HNE antibody (catalog no. HNE11-S, Alpha Diagnostic International Inc., San Antonio, TX, USA) at 4 °C for 2 h followed by a donkey anti-rabbit IgG fluorescence-conjugated secondary antibody (catalog no. A21206, Invitrogen Corp., Waltham, MA, USA) for 2 h at room temperature and counterstained with DAPI.
To determine 4-HNE immunoreactivity, fluorescence intensity was measured on each section and calculated using ImageJ software. Once the green channel was separated from the red channel in an image, fluorescence intensities from the areas of interest were measured using the integral/density feature in the ImageJ program and data were extracted and averaged within the group. Fluorescence was averaged across 5 retinas per group in retinal nuclear layers. Data were normalized with respect to controls. The values were represented as fold changes over controls.
4.4. Mass Spectrometry
Protein expression profiles of ocular tissue containing the retina were determined in eighteen mice (n = 6/group) from three sample groups: µg, µg + 1 g, and HC, using high resolution mass spectrometry.
Five micrograms of protein lysate were resolved by a 4–20% Tris-Glycine SDS-PAGE (Life Technologies Corp., Waltham, MA, USA). The gel was sliced into 24 gel slices that were 2 mm thick for each sample. Gel slices were destained in 50% methanol (Thermo Fisher Scientific, Hampton, NH, USA), 100 mM ammonium bicarbonate (Sigma-Aldrich, St. Louis, MO, USA), followed by reduction in 10 mM Tris (2-carboxyethyl) phosphine (Pierce, Dallas, TX, USA) and alkylation in 50 mM iodoacetamide (Sigma-Aldrich). Gel slices were then dehydrated in acetonitrile (Thermo Fisher Scientific), followed by addition of 100 ng porcine sequencing grade modified trypsin (Promega Corp.) in 100 mM ammonium bicarbonate (Sigma-Aldrich) and incubation at 37 °C for 12–16 h. Peptide products were then acidified in 0.1% formic acid (Pierce). Tryptic peptides were separated by reverse phase Jupiter Proteo resin (Phenomenex, Torrance, CA, USA) on a 100 × 0.075 mm column using a nanoAcquity Ultra Performance LC™ (UPLC™) system (Waters Corp., Milford, MA, USA). Peptides were eluted using a 30 min gradient from 97:3 to 60:40 buffer A:B ratio. (Buffer A = 0.1% formic acid, 0.5% acetonitrile; buffer B = 0.1% formic acid, 90% acetonitrile). Eluted peptides were ionized by electrospray (1.9 kV) followed by MS/MS analysis using collision-induced dissociation on an LTQ Orbitrap Velos mass spectrometer (Thermo Fisher Scientific). MS data were acquired using the Fourier Transform ion cyclotron resonance spectrometry (FTMS) analyzer in profile mode at a resolution of 60,000 over a range of 375 to 1500 m/z. MS/MS data were acquired for the top 15 peaks from each MS scan using the ion trap analyzer in centroid mode and normal mass range with a normalized collision energy of 35.0.
4.5. Data Analysis
Proteins were identified by searching the UniProtKB database (2017-03 release; restricted to Mus musculus; 81,557 entries) using the Andromeda search engine in MaxQuant (version 1.5.7.4, Max Planck Institute of biochemistry, Martinsried, Germany). Search parameters were as follows: trypsin digestion with up to three missed cleavages; fixed modification of carbamidomethyl of cysteine; variable modifications of oxidation on methionine and acetyl on N-terminus; first search set to 5 ppm precursor ion tolerance and the main search was set to 3 ppm; selected label-free quantitation with iBAQ with a minimum ratio of 1. A contaminants database was used for the first search to identify commonly identified contaminants.
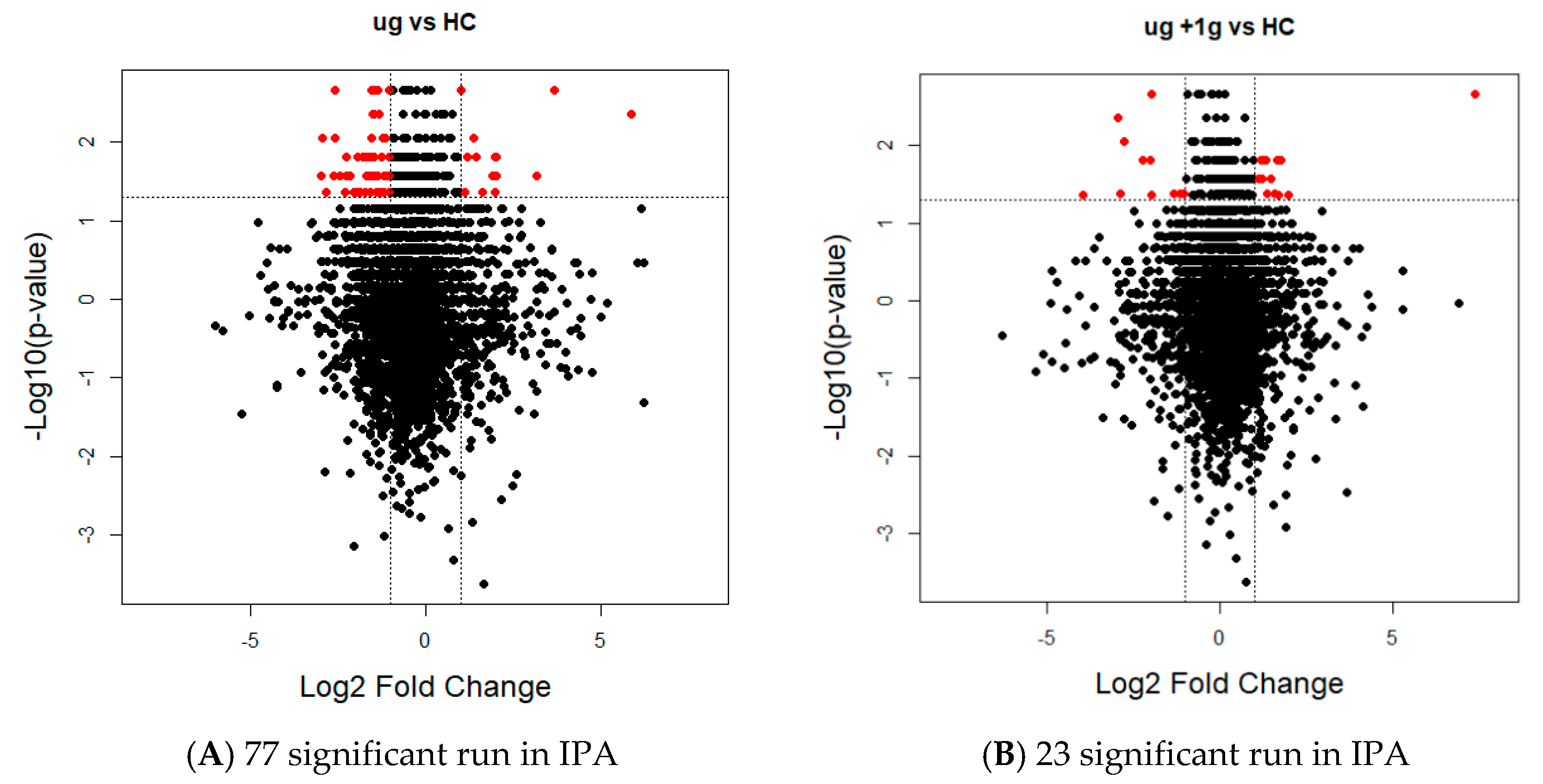
The MaxQuant iBAQ values for each sample were converted to relative iBAQ (riBAQ) intensities by dividing the iBAQ intensity for each protein by the sum of all iBAQ intensities for all proteins identified in the sample. The median of all relative iBAQ intensities for each sample was then used to normalize between the samples. The adjusted riBAQ intensities were then imported into Perseus (version 1.5.6.0, Max Planck Institute of biochemistry, Martinsried, Germany). Perseus was used to log2 transform the data and impute the missing values using a normal distribution with a width of 0.3 and a downshift of 2 standard deviations. Non-parametric Mann–Whitney U tests were used to compare habitat control with the µg and µg + 1 g groups separately. The p-values were corrected by Benjamini–Hochberg to account for multiple testing. Fold changes were also calculated for each comparison. Proteins that were significant by the FDR-adjusted p-value < 0.05 and that had a fold change greater than 2 were considered to be differentially expressed. The heat maps were generated using the Euclidean distance metric and the data was standardized by the mean and standard deviation before performing hierarchical clustering with pheatmap R package (maintained by Raivo Kolde, Massachusetts General Hospital, Boston, MA, USA). Volcano plots were generated using R. These proteins were then analyzed using the IPA pathway analysis.
4.6. Statistical Analysis
The results obtained from all experiments were analyzed by one-way analysis of variance (ANOVA) followed by Tukey’s post hoc multiple-comparison test (Sigma Plot for Windows, version 13.0; Systat Software, Inc., Point Richmond, CA, USA). The significance level was set at p < 0.05. Data are shown as mean ± standard error (SEM).

 ,
, 








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}