Role of Ttca of Citrobacter Werkmanii in Bacterial Growth, Biocides Resistance, Biofilm Formation and Swimming Motility

Abstract
:1. Introduction
2. Results
2.1. Identification of the Insertion Sites of Tn5
2.2. Characterization of ttcA Gene
2.3. Effect of BIT on Bacterial Growth
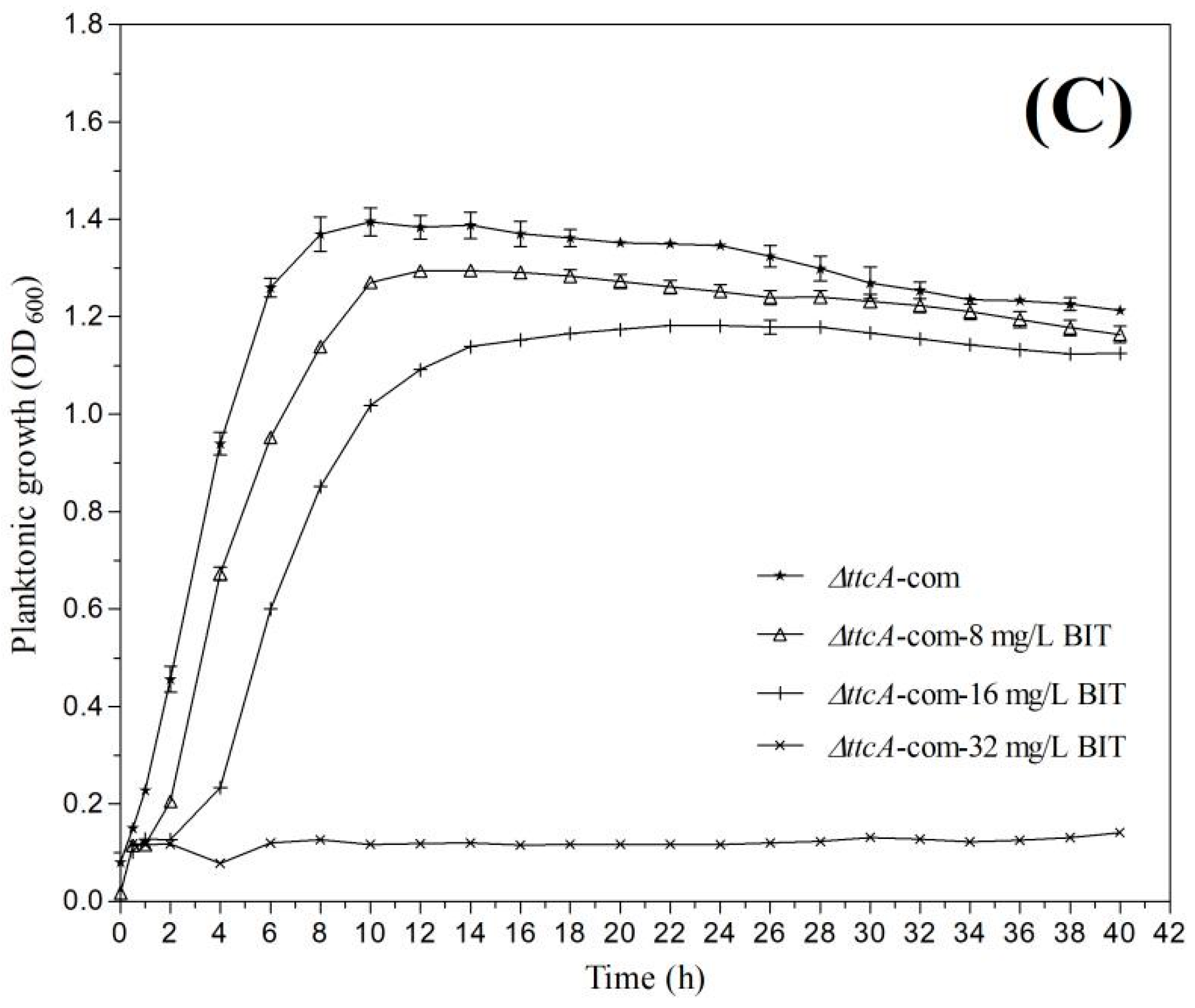
2.4. Effect of BIT on Planktonic Growth and Initial Biofilm Formation
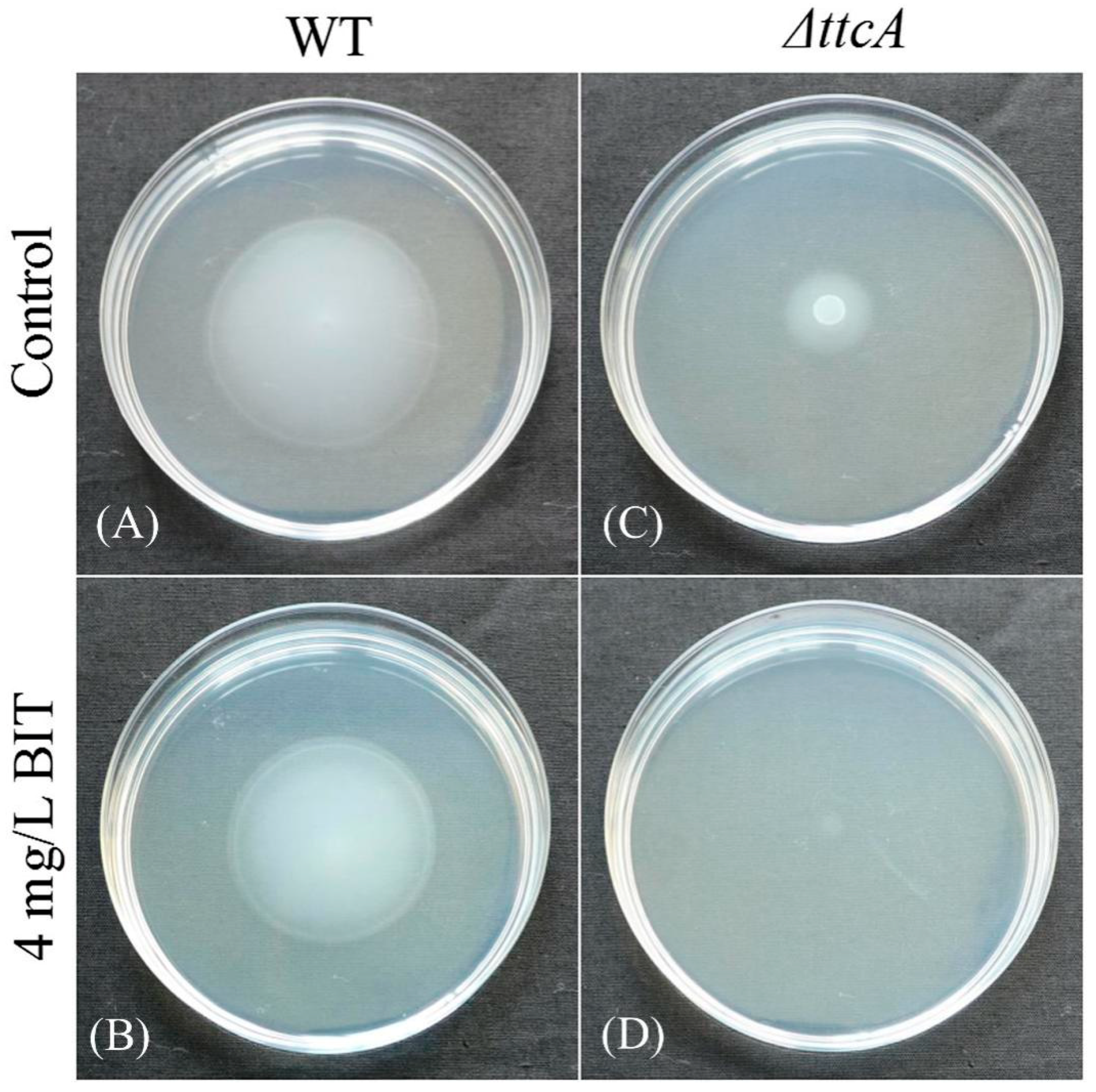
2.5. Role of ttcA in Bacterial Motility
2.6. Effect of BIT on Biofilm Architecture
3. Discussion
4. Materials and Methods
4.1. Bacterial Strains and Chemicals
4.2. Construction of Transposon for Mutagenesis
4.3. Identification of Transposon Insertion Site
4.4. Construction of Gene Knockout Mutants and Complementary Strains
4.5. Determination of the Bacterial Growth Curves
4.6. In Vitro Inhibitory Effect of BIT on Initial Biofilm Formation
4.7. Motility Assay
4.8. Observation of Biofilm Structures with Confocal Laser Scanning Microscopy (CLSM)
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| BIT | 1,2-benzisothiazolin-3-one |
| LB | Luria Bertani |
| CLSM | Confocal Laser Scanning Microscopy |
References
- Pucci, M.J.; Podos, S.D.; Thanassi, J.A.; Leggio, M.J.; Bradbury, B.J.; Deshpande, M. In vitro and in vivo profiles of ACH-702, an isothiazoloquinolone, against bacterial pathogens. Antimicrob. Agents Chemother. 2011, 55, 2860–2871. [Google Scholar] [CrossRef] [PubMed]
- Vicentini, C.B.; Romagnoli, C.; Manfredini, S.; Rossi, D.; Mares, D. Pyrazolo [3,4-c] isothiazole and isothiazolo [4,3-d] isoxazole derivatives as antifungal agents. Pharm. Biol. 2011, 49, 545–552. [Google Scholar] [CrossRef] [PubMed]
- Williams, T.M. The mechanism of action of isothiazolone biocide. Power Plant Chem. 2007, 9, 14–22. [Google Scholar]
- Zhou, G.; Shi, Q.S.; Huang, X.M.; Xie, X.B. Comparison of transcriptomes of wild-type and isothiazolone-resistant Pseudomonas aeruginosa by using RNA-seq. Mol. Biol. Rep. 2016, 43, 527–540. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Shi, Q.S.; Ouyang, Y.S.; Chen, Y.B. Involvement of outer membrane proteins and peroxide-sensor genes in Burkholderia cepacia resistance to isothiazolone. World J. Microbiol. Biotechnol. 2014, 30, 1251–1260. [Google Scholar] [CrossRef] [PubMed]
- Lipsky, B.A.; Hook, E.W.; Smith, A.A.; Plorde, J.J. Citrobacter infections in humans: Experience at the Seattle Veterans Administration Medical Center and a review of the literature. Rev. Infect. Dis. 1980, 2, 746–760. [Google Scholar] [CrossRef] [PubMed]
- Arens, S.; Verhaegen, J.; Verbist, L. Differentiation and susceptibility of Citrobacter isolates from patients in a university hospital. Clin. Microbiol. Infect. 1997, 3, 53–57. [Google Scholar] [CrossRef] [PubMed]
- Badger, J.L.; Stins, M.F.; Kim, K.S. Citrobacter freundii invades and replicates in human brain microvascular endothelial cells. Infect. Immun. 1999, 67, 4208–4215. [Google Scholar] [PubMed]
- Doran, T.I. The role of Citrobacter in clinical disease of children: Review. Clin. Infect. Dis. 1999, 28, 384–394. [Google Scholar] [CrossRef] [PubMed]
- Macaskie, L.E.; Empson, R.M.; Lin, F.; Tolley, M.R. Enzymatically-mediated uranium accumulation and uranium recovery using a Citrobacter sp. Immobilised as a biofilm within a plug-flow reactor. J. Chem. Technol. Biotechnol. 1995, 63, 1–16. [Google Scholar] [CrossRef]
- Finlay, J.A.; Allan, V.J.M.; Conner, A.; Callow, M.E.; Basnakova, G.; Macaskie, L.E. Phosphate release and heavy metal accumulation by biofilm-immobilized and chemically-coupled cells of a citrobacter sp. pre-grown in continuous culture. Biotechnol. Bioeng. 1999, 63, 87–97. [Google Scholar] [CrossRef]
- Zhou, G.; Li, L.J.; Shi, Q.S.; Ouyang, Y.S.; Chen, Y.B.; Hu, W.F. Effects of nutritional and environmental conditions on planktonic growth and biofilm formation for Citrobacter werkmanii BF-6. J. Microbiol. Biotechnol. 2013, 23, 1673–1682. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Peng, H.; Wang, Y.S.; Huang, X.M.; Xie, X.B.; Shi, Q.S. Complete genome sequence of Citrobacter werkmanii strain BF-6 isolated from industrial putrefaction. BMC Genom. 2017, 18, 765. [Google Scholar] [CrossRef] [PubMed]
- Collier, P.J.; Ramsey, A.; Waigh, R.D.; Douglas, K.T.; Austin, P.; Gilbert, P. Chemical reactivity of some isothiazolone biocides. J. Appl. Microbiol. 1990, 69, 578–584. [Google Scholar] [CrossRef]
- Zhou, G.; Shi, Q.S.; Huang, X.M.; Xie, X.B.; Chen, Y.B. Insights into Pseudomonas aeruginosa ATCC9027 resistance to isothiazolones through proteomics. Microb. Drug Resist. 2015, 21, 140–148. [Google Scholar] [CrossRef] [PubMed]
- Bouvier, D.; Labessan, N.; Clémancey, M.; Latour, J.-M.; Ravanat, J.-L.; Fontecave, M.; Atta, M. TtcA a new tRNA-thioltransferase with an Fe-S cluster. Nucleic Acids Res. 2014, 42, 7960–7970. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jäger, G.; Leipuviene, R.; Pollard, M.G.; Qian, Q.; Björk, G.R. The conserved Cys-X1-X2-Cys motif present in the TtcA protein is required for the thiolation of cytidine in position 32 of tRNA from Salmonella enterica serovar Typhimurium. J. Bacteriol. 2004, 186, 750–757. [Google Scholar] [CrossRef] [PubMed]
- Flemming, H.C.; Wingender, J. The biofilm matrix. Nat. Rev. Microbiol. 2010, 8, 623–633. [Google Scholar] [CrossRef] [PubMed]
- Zhou, G.; Shi, Q.S.; Huang, X.M.; Xie, X.B. The three bacterial lines of defense against antimicrobial agents. Int. J. Mol. Sci. 2015, 16, 21711–21733. [Google Scholar] [CrossRef] [PubMed]
- Billings, N.; Millan, M.; Caldara, M.; Rusconi, R.; Tarasova, Y.; Stocker, R.; Ribbeck, K. The extracellular matrix component Psl provides fast-acting antibiotic defense in Pseudomonas aeruginosa biofilms. PLoS Pathog. 2013, 9, e1003526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mah, T.F. Biofilm-specific antibiotic resistance. Future Microbiol. 2012, 7, 1061–1072. [Google Scholar] [CrossRef] [PubMed]
- Venkatesan, N.; Perumal, G.; Doble, M. Bacterial resistance in biofilm-associated bacteria. Future Microbiol. 2015, 10, 1743–1750. [Google Scholar] [CrossRef] [PubMed]
- Ha, D.G.; Kuchma, S.L.; O’Toole, G.A. Plate-based assay for swimming motility in Pseudomonas aeruginosa. Methods Mol. Biol. 2014, 1149, 59–65. [Google Scholar] [PubMed]
- Rashid, M.H.; Kornberg, A. Inorganic polyphosphate is needed for swimming, swarming and twitching motilities of Pseudomonas aeruginosa. Proc. Natl. Acad. Sci. USA 2000, 97, 4885–4890. [Google Scholar] [CrossRef] [PubMed]
- Murray, T.S.; Kazmierczak, B.I. FlhF is required for swimming and swarming in Pseudomonas aeruginosa. J. Bacteriol. 2006, 188, 6995–7004. [Google Scholar] [CrossRef] [PubMed]
- Li, L.J.; Zhou, G.; Shi, Q.S.; Chen, Y.B.; Ouyang, Y.S.; Hu, W.F. Identification and biofilm formation characterization of Citrobacter werkmanii isolated from industrial spoilage. Microbiol. China 2014, 41, 2–7. [Google Scholar]
- Larsen, R.A.; Wilson, M.M.; Guss, A.M.; Metcalf, W.W. Genetic analysis of pigment biosynthesis in Xanthobacter autotrophicus Py2 using a new, highly efficient transposon mutagenesis system that is functional in a wide variety of bacteria. Arch. Microbiol. 2002, 178, 193–201. [Google Scholar] [CrossRef] [PubMed]
- Cong, Y.; Wang, J.; Chen, Z.; Xiong, K.; Xu, Q.; Hu, F. Characterization of swarming motility in Citrobacter freundii. FEMS Microbiol. Lett. 2011, 317, 160–171. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.C.; Wu, Y.Q.; Cao, D.; Zhang, W.B.; Wang, H.H. Only acyl carrier protein 1 (AcpP1) functions in Pseudomonas aeruginosa fatty acid synthesis. Front. Microbiol. 2017, 8, 2186. [Google Scholar] [CrossRef] [PubMed]
- Stepanović, S.; Vuković, D.; Dakić, I.; Savić, B.; Švabić-Vlahović, M. A modified microtiter-plate test for quantification of staphylococcal biofilm formation. J. Microbiol. Methods 2000, 40, 175–179. [Google Scholar] [CrossRef]
- Huang, Y.H.; Lin, J.S.; Ma, J.C.; Wang, H.H. Functional characterization of triclosan-resistant enoyl-acyl-carrier protein reductase (FabV) in Pseudomonas aeruginosa. Front. Microbiol. 2016, 7, 1903. [Google Scholar] [CrossRef] [PubMed]
- Shukla, S.K.; Rao, T.S. Effect of calcium on Staphylococcus aureus biofilm architecture: A confocal laser scanning microscopic study. Colloid Surf. B Biointerfaces 2013, 103, 448–454. [Google Scholar] [CrossRef] [PubMed]
- Heydorn, A.; Ersbøll, B.K.; Hentzer, M.; Parsek, M.R.; Givskov, M.; Molin, S. Experimental reproducibility in flow-chamber biofilms. Microbiology 2000, 146, 2409–2415. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heydorn, A.; Nielsen, A.T.; Hentzer, M.; Sternberg, C.; Givskov, M.; Ersbøll, B.K.; Molin, S. Quantification of biofilm structures by the novel computer program COMSTAT. Microbiology 2000, 146, 2395–2407. [Google Scholar] [CrossRef] [PubMed]
- Tang, Q.Y.; Feng, M.G. DPS Data Processing System: Experimental Design, Statistical Analysis and Data Mining; Science Press: Beijing, China, 2007. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Number | Locus Tag | Gene Name | Description |
|---|---|---|---|
| 64 | B2G73_RS05960 | ttcA | tRNA 2-thiocytidine(32) synthetase |
| 94 | B2G73_RS07535 | - | LysR family transcriptional regulator |
| 111 | B2G73_RS05205 | - | HNH endonuclease |
| Primer Names | Forward Primer Sequences (5′-3′) | Reverse Primer Sequences (5′-3′) | Application |
|---|---|---|---|
| TnI-F/R | GGAGAGGCTATTCGGCTATG | GTAAGGTGATCCGGTGGATG | Identification of Tn5 inserted sites |
| ttcA-WJ-F/R | ATTATCGCTTCTCACCGACCTG | AACGTCCAGCCACGGTAAAATC | Identification of ttcA deletion |
| ttcAP-F/R | GCTCTAGAGTGATGCATACTCTTCTGAT | CCCAAGCTTTTACTTCACTTCCACAACGT | ttcA complementation |
| ttcA-BS-F/R | CTTCAATATGATCGAAGACGGC | TTGGGTATCATCCTCTTCTGGC | Partial sequence of ttcA gene |
| Parameters * | WT (Mean ± SD) | ΔttcA (Mean ± SD) | ||||
|---|---|---|---|---|---|---|
| 0 mg/L BIT | 4 mg/L BIT | 16 mg/L BIT | 0 mg/L BIT | 4 mg/L BIT | 16 mg/L BIT | |
| Maximum thickness (μm) | 12.00 ± 2.00 | 11.33 ± 1.15 | 8.00 ± 0.00 | 20.77 ± 2.69 | 18.00 ± 4.00 | 16.67 ± 2.31 |
| Average thickness (μm) | 8.26 ± 1.40 | 5.86 ± 1.17 | 3.54 ± 1.29 | 17.33 ± 2.31 | 9.58 ± 1.72 | 5.79 ± 1.27 |
| Total biomass (μm3/μm2) | 12.06 ± 0.98 | 5.58 ± 1.20 | 3.42 ± 1.06 | 14.52 ± 0.14 | 9.78 ± 1.63 | 6.14 ± 1.27 |
| Average colony size (μm2) | 195.67 ± 17.65 | 182.18 ± 12.12 | 59.00 ± 6.58 | 213.75 ± 7.93 | 192.55 ± 18.54 | 155.90 ± 17.86 |
| Roughness coefficient | 0.22 ± 0.02 | 0.51 ± 0.02 | 0.82 ± 0.21 | 0.08 ± 0.02 | 0.11 ± 0.01 | 0.36 ± 0.06 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhou, G.; Wang, Y.-S.; Peng, H.; Huang, X.-M.; Xie, X.-B.; Shi, Q.-S. Role of Ttca of Citrobacter Werkmanii in Bacterial Growth, Biocides Resistance, Biofilm Formation and Swimming Motility. Int. J. Mol. Sci. 2018, 19, 2644. https://doi.org/10.3390/ijms19092644
Zhou G, Wang Y-S, Peng H, Huang X-M, Xie X-B, Shi Q-S. Role of Ttca of Citrobacter Werkmanii in Bacterial Growth, Biocides Resistance, Biofilm Formation and Swimming Motility. International Journal of Molecular Sciences. 2018; 19(9):2644. https://doi.org/10.3390/ijms19092644
Chicago/Turabian StyleZhou, Gang, Ying-Si Wang, Hong Peng, Xiao-Mo Huang, Xiao-Bao Xie, and Qing-Shan Shi. 2018. "Role of Ttca of Citrobacter Werkmanii in Bacterial Growth, Biocides Resistance, Biofilm Formation and Swimming Motility" International Journal of Molecular Sciences 19, no. 9: 2644. https://doi.org/10.3390/ijms19092644