HDAC-Linked “Proliferative” miRNA Expression Pattern in Pancreatic Neuroendocrine Tumors

 , ,
, ,
Abstract
:
1. Introduction
2. Results
2.1. Clinical Characteristics of pNET Patients
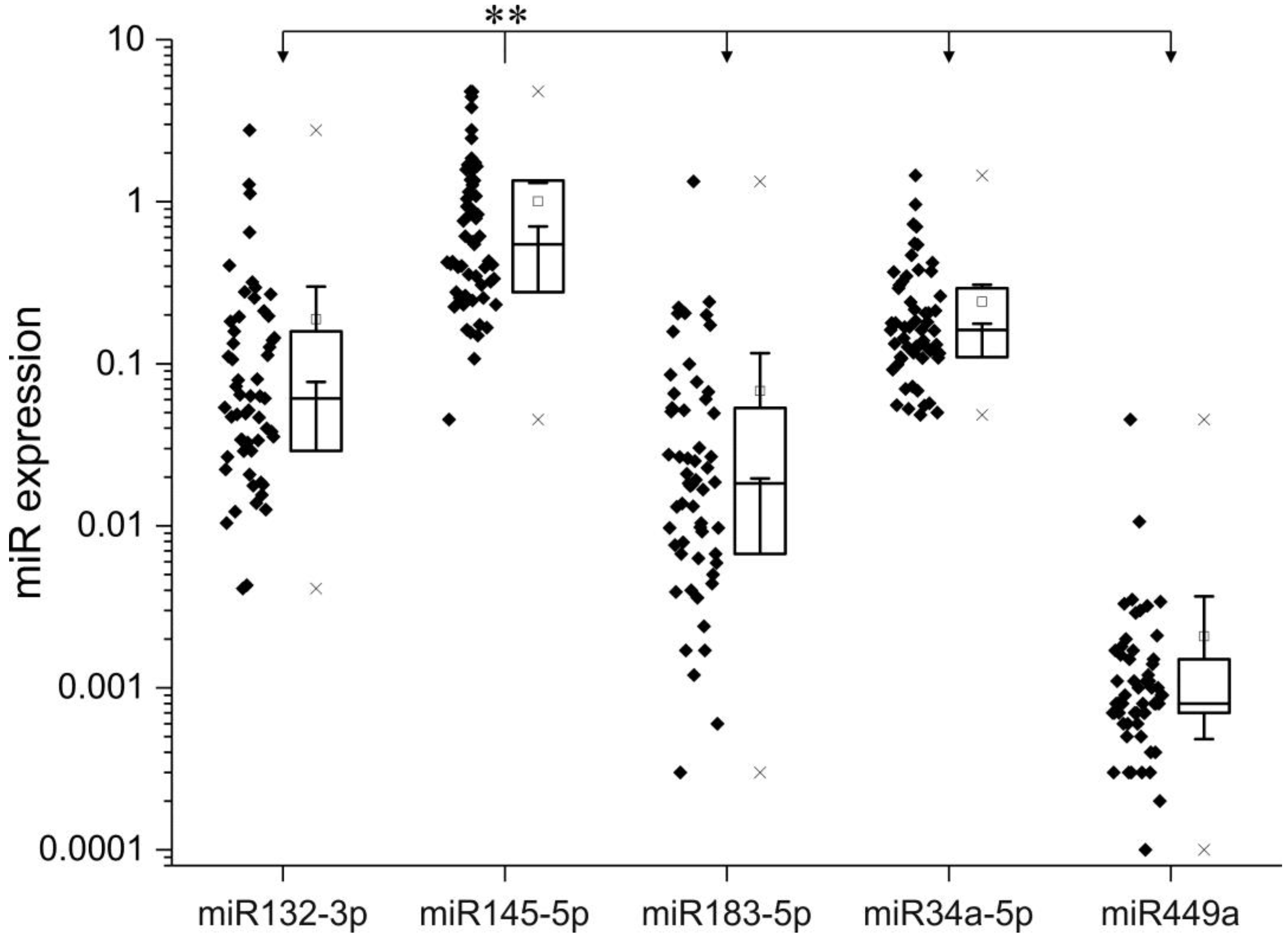
2.2. Expression of miRNA in pNETs
2.3. Correlation Analysis of miRNA and HDAC Expression
2.4. Association of miRNA Expression with Grading and Mitotic/Proliferative Status
2.5. miRNA Expression and Clinicopathological Characteristics
2.6. Impact of miRNA Expression on DFS and OS
3. Discussion
4. Materials and Methods
4.1. Clinical and Pathological Characterization of pNET Cases
4.2. Isolation of Total miRNA from FFPE Samples and Expression Analysis by RT-PCR
4.3. Immunohistochemistry and Processing for Markers of HDACs (1-6, 8-11 and Sirt1)
4.4. Interpretation and Scoring of the IHC
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| Cyt | Cytoplasmatic |
| DFS | Disease-free survival |
| EMT | Epithelial to mesenchymal transition |
| FFPE | Formalin-fixed paraffin-embedded |
| HDAC | Histone deacetylases |
| HPF | High power fields |
| IHC | Immunohistochemistry |
| miRNA | microRNA |
| nuc/ncl | Nuclear |
| OS | Overall survival |
| pNET | Pancreatic neuroendocrine tumor |
| SD | Standard deviation |
| TMA | Tissue microarray |
References
- Alexandraki, K.I.; Kaltsas, G. Gastroenteropancreatic neuroendocrine tumors: New insights in the diagnosis and therapy. Endocrine 2012, 41, 40–52. [Google Scholar] [CrossRef] [PubMed]
- Schimmack, S.; Svejda, B.; Lawrence, B.; Kidd, M.; Modlin, I.M. The diversity and commonalities of gastroenteropancreatic neuroendocrine tumors. Langenbecks Arch. Surg. 2011, 396, 273–298. [Google Scholar] [CrossRef] [PubMed]
- Lawrence, B.; Gustafsson, B.I.; Chan, A.; Svejda, B.; Kidd, M.; Modlin, I.M. The epidemiology of gastroenteropancreatic neuroendocrine tumors. Endocrinol. Metab. Clin. N. Am. 2011, 40, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Ehehalt, F.; Saeger, H.D.; Schmidt, C.M.; Grutzmann, R. Neuroendocrine tumors of the pancreas. Oncologist 2009, 14, 456–467. [Google Scholar] [CrossRef] [PubMed]
- Alexandraki, K.I.; Karapanagioti, A.; Karoumpalis, I.; Boutzios, G.; Kaltsas, G.A. Advances and current concepts in the medical management of gastroenteropancreatic neuroendocrine neoplasms. BioMed Res. Int. 2017, 2017, 9856140. [Google Scholar] [CrossRef] [PubMed]
- Virani, S.; Colacino, J.A.; Kim, J.H.; Rozek, L.S. Cancer epigenetics: A brief review. ILAR J. 2012, 53, 359–369. [Google Scholar] [CrossRef] [PubMed]
- Kanwal, R.; Gupta, K.; Gupta, S. Cancer epigenetics: An introduction. Methods Mol. Biol. 2015, 1238, 3–25. [Google Scholar] [PubMed]
- Felsenfeld, G. A brief history of epigenetics. Cold Spring Harb. Perspect. Biol. 2014, 6, a018200. [Google Scholar] [CrossRef] [PubMed]
- He, L.; Hannon, G.J. MicroRNAs: Small RNAs with a big role in gene regulation. Nat. Rev. Genet. 2004, 5, 522–531. [Google Scholar] [CrossRef] [PubMed]
- Ropero, S.; Esteller, M. The role of histone deacetylases (HDACs) in human cancer. Mol. Oncol. 2007, 1, 19–25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, M.; Jia, Y.; Yu, Z.; House, M.G.; Esteller, M.; Brock, M.V.; Herman, J.G. Epigenetic changes associated with neoplasms of the exocrine and endocrine pancreas. Discov. Med. 2014, 17, 67–73. [Google Scholar] [PubMed]
- Klieser, E.; Swierczynski, S.; Mayr, C.; Schmidt, J.; Neureiter, D.; Kiesslich, T.; Illig, R. Role of histone deacetylases in pancreas: Implications for pathogenesis and therapy. World J. Gastrointest. Oncol. 2015, 7, 473–483. [Google Scholar] [CrossRef] [PubMed]
- Croce, C.M. Causes and consequences of microRNA dysregulation in cancer. Nat. Rev. Genet. 2009, 10, 704–714. [Google Scholar] [CrossRef] [PubMed]
- Fiaschetti, G.; Abela, L.; Nonoguchi, N.; Dubuc, A.M.; Remke, M.; Boro, A.; Grunder, E.; Siler, U.; Ohgaki, H.; Taylor, M.D.; et al. Epigenetic silencing of miRNA-9 is associated with HES1 oncogenic activity and poor prognosis of medulloblastoma. Br. J. Cancer 2014, 110, 636–647. [Google Scholar] [CrossRef] [PubMed]
- Holubekova, V.; Mendelova, A.; Jasek, K.; Mersakova, S.; Zubor, P.; Lasabova, Z. Epigenetic regulation by DNA methylation and miRNA molecules in cancer. Future Oncol. 2017, 13, 2217–2222. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kwon, H.; Song, K.; Han, C.; Zhang, J.; Lu, L.; Chen, W.; Wu, T. Epigenetic silencing of miRNA-34a in human cholangiocarcinoma via EZH2 and DNA methylation: Impact on regulation of notch pathway. Am. J. Pathol. 2017, 187, 2288–2299. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Xu, Z.; Li, B.; Zhang, Z.; Luo, H.; Wang, Y.; Lu, Z.; Wu, X. Epigenetic silencing of miRNA-9 is correlated with promoter-proximal CpG island hypermethylation in gastric cancer in vitro and in vivo. Int. J. Oncol. 2014, 45, 2576–2586. [Google Scholar] [CrossRef] [PubMed]
- Mayr, C.; Beyreis, M.; Wagner, A.; Pichler, M.; Neureiter, D.; Kiesslich, T. Deregulated microRNAs in biliary tract cancer: Functional targets and potential biomarkers. BioMed Res. Int. 2016, 2016, 4805270. [Google Scholar] [CrossRef] [PubMed]
- Noonan, E.J.; Place, R.F.; Pookot, D.; Basak, S.; Whitson, J.M.; Hirata, H.; Giardina, C.; Dahiya, R. miR-449a targets HDAC-1 and induces growth arrest in prostate cancer. Oncogene 2009, 28, 1714–1724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sampath, D.; Liu, C.; Vasan, K.; Sulda, M.; Puduvalli, V.K.; Wierda, W.G.; Keating, M.J. Histone deacetylases mediate the silencing of miR-15a, miR-16, and miR-29b in chronic lymphocytic leukemia. Blood 2012, 119, 1162–1172. [Google Scholar] [CrossRef] [PubMed]
- Swierczynski, S.; Klieser, E.; Illig, R.; Alinger-Scharinger, B.; Kiesslich, T.; Neureiter, D. Histone deacetylation meets miRNA: Epigenetics and post-transcriptional regulation in cancer and chronic diseases. Expert Opin. Biol. Ther. 2015, 15, 651–664. [Google Scholar] [CrossRef] [PubMed]
- Klieser, E.; Urbas, R.; Stattner, S.; Primavesi, F.; Jager, T.; Dinnewitzer, A.; Mayr, C.; Kiesslich, T.; Holzmann, K.; Di Fazio, P.; et al. Comprehensive immunohistochemical analysis of histone deacetylases in pancreatic neuroendocrine tumors: HDAC5 as a predictor of poor clinical outcome. Hum. Pathol. 2017, 65, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Govindan, R.; Wang, L.; Liu, P.Y.; Goodgame, B.; Wen, W.; Sezhiyan, A.; Pfeifer, J.; Li, Y.F.; Hua, X.; et al. MicroRNA profiling and prediction of recurrence/relapse-free survival in stage I lung cancer. Carcinogenesis 2012, 33, 1046–1054. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Han, Q.; Zhang, H.Y.; Zhong, B.L.; Wang, X.J.; Zhang, B.; Chen, H. MicroRNA-145 inhibits cell migration and invasion and regulates epithelial-mesenchymal transition (EMT) by targeting connective tissue growth factor (CTGF) in esophageal squamous cell carcinoma. Med. Sci. Monit. 2016, 22, 3925–3934. [Google Scholar] [CrossRef] [PubMed]
- Doberstein, K.; Steinmeyer, N.; Hartmetz, A.K.; Eberhardt, W.; Mittelbronn, M.; Harter, P.N.; Juengel, E.; Blaheta, R.; Pfeilschifter, J.; Gutwein, P. MicroRNA-145 targets the metalloprotease ADAM17 and is suppressed in renal cell carcinoma patients. Neoplasia 2013, 15, 218–230. [Google Scholar] [CrossRef] [PubMed]
- Larne, O.; Hagman, Z.; Lilja, H.; Bjartell, A.; Edsjo, A.; Ceder, Y. MiR-145 suppress the androgen receptor in prostate cancer cells and correlates to prostate cancer prognosis. Carcinogenesis 2015, 36, 858–866. [Google Scholar] [CrossRef] [PubMed]
- Chiyomaru, T.; Enokida, H.; Tatarano, S.; Kawahara, K.; Uchida, Y.; Nishiyama, K.; Fujimura, L.; Kikkawa, N.; Seki, N.; Nakagawa, M. MiR-145 and miR-133a function as tumour suppressors and directly regulate FSCN1 expression in bladder cancer. Br. J. Cancer 2010, 102, 883–891. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Yin, L.; Jiang, N.; Guo, W.J.; Gu, J.J.; Chen, M.; Xia, Y.Y.; Wu, J.Z.; Chen, D.; Wu, J.F.; et al. MiR-145, a microRNA targeting ADAM17, inhibits the invasion and migration of nasopharyngeal carcinoma cells. Exp. Cell Res. 2015, 338, 232–238. [Google Scholar] [CrossRef] [PubMed]
- Buurman, R.; Gurlevik, E.; Schaffer, V.; Eilers, M.; Sandbothe, M.; Kreipe, H.; Wilkens, L.; Schlegelberger, B.; Kuhnel, F.; Skawran, B. Histone deacetylases activate hepatocyte growth factor signaling by repressing microRNA-449 in hepatocellular carcinoma cells. Gastroenterology 2012, 143, 811–820. [Google Scholar] [CrossRef] [PubMed]
- Jeon, H.S.; Lee, S.Y.; Lee, E.J.; Yun, S.C.; Cha, E.J.; Choi, E.; Na, M.J.; Park, J.Y.; Kang, J.; Son, J.W. Combining microRNA-449a/b with a HDAC inhibitor has a synergistic effect on growth arrest in lung cancer. Lung Cancer 2012, 76, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Li, F.; Liang, J.; Bai, L. MicroRNA-449a functions as a tumor suppressor in pancreatic cancer by the epigenetic regulation of ATDC expression. Biomed. Pharmacother. 2018, 103, 782–789. [Google Scholar] [CrossRef] [PubMed]
- Bou Kheir, T.; Futoma-Kazmierczak, E.; Jacobsen, A.; Krogh, A.; Bardram, L.; Hother, C.; Gronbaek, K.; Federspiel, B.; Lund, A.H.; Friis-Hansen, L. MiR-449 inhibits cell proliferation and is down-regulated in gastric cancer. Mol. Cancer 2011, 10, 29. [Google Scholar] [CrossRef] [PubMed]
- Kang, Z.H.; Wang, C.Y.; Zhang, W.L.; Zhang, J.T.; Yuan, C.H.; Zhao, P.W.; Lin, Y.Y.; Hong, S.; Li, C.Y.; Wang, L. Histone deacetylase HDAC4 promotes gastric cancer SGC-7901 cells progression via p21 repression. PLoS ONE 2014, 9, e98894. [Google Scholar] [CrossRef] [PubMed]
- Yong-Ming, H.; Ai-Jun, J.; Xiao-Yue, X.; Jian-Wei, L.; Chen, Y.; Ye, C. miR-449a: A potential therapeutic agent for cancer. Anti-Cancer Drugs 2017, 28, 1067–1078. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Belaguli, N.; Berger, D.H. MicroRNA and colorectal cancer. World J. Surg. 2009, 33, 638–646. [Google Scholar] [CrossRef] [PubMed]
- Anand, S.; Majeti, B.K.; Acevedo, L.M.; Murphy, E.A.; Mukthavaram, R.; Scheppke, L.; Huang, M.; Shields, D.J.; Lindquist, J.N.; Lapinski, P.E.; et al. MicroRNA-132-mediated loss of p120RasGAP activates the endothelium to facilitate pathological angiogenesis. Nat. Med. 2010, 16, 909–914. [Google Scholar] [CrossRef] [PubMed]
- Formosa, A.; Lena, A.M.; Markert, E.K.; Cortelli, S.; Miano, R.; Mauriello, A.; Croce, N.; Vandesompele, J.; Mestdagh, P.; Finazzi-Agro, E.; et al. DNA methylation silences miR-132 in prostate cancer. Oncogene 2013, 32, 127–134. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Gao, T.; Tang, J.; Cai, H.; Lin, L.; Fu, S. Loss of microRNA-132 predicts poor prognosis in patients with primary osteosarcoma. Mol. Cell. Biochem. 2013, 381, 9–15. [Google Scholar] [CrossRef] [PubMed]
- Bommer, G.T.; Gerin, I.; Feng, Y.; Kaczorowski, A.J.; Kuick, R.; Love, R.E.; Zhai, Y.; Giordano, T.J.; Qin, Z.S.; Moore, B.B.; et al. p53-mediated activation of miRNA34 candidate tumor-suppressor genes. Curr. Biol. 2007, 17, 1298–1307. [Google Scholar] [CrossRef] [PubMed]
- Yang, C.L.; Zheng, X.L.; Ye, K.; Ge, H.; Sun, Y.N.; Lu, Y.F.; Fan, Q.X. MicroRNA-183 acts as a tumor suppressor in human non-small cell lung cancer by down-regulating MTA1. Cell. Physiol. Biochem. 2018, 46, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Xiong, H.; Chen, R.; Liu, S.; Lin, Q.; Chen, H.; Jiang, Q. MicroRNA-183 induces epithelial-mesenchymal transition and promotes endometrial cancer cell migration and invasion in by targeting CPEB1. J. Cell. Biochem. 2018. [Google Scholar] [CrossRef] [PubMed]
- Gao, J.M.; Huang, L.Z.; Huang, Z.G.; He, R.Q. Clinical value and potential pathways of miR-183-5p in bladder cancer: A study based on miRNA-seq data and bioinformatics analysis. Oncol. Lett. 2018, 15, 5056–5070. [Google Scholar] [CrossRef] [PubMed]
- Gu, W.; Gao, T.; Shen, J.; Sun, Y.; Zheng, X.; Wang, J.; Ma, J.; Hu, X.Y.; Li, J.; Hu, M.J. MicroRNA-183 inhibits apoptosis and promotes proliferation and invasion of gastric cancer cells by targeting PDCD4. Int. J. Clin. Exp. Med. 2014, 7, 2519–2529. [Google Scholar] [PubMed]
- Ren, L.H.; Chen, W.X.; Li, S.; He, X.Y.; Zhang, Z.M.; Li, M.; Cao, R.S.; Hao, B.; Zhang, H.J.; Qiu, H.Q.; et al. MicroRNA-183 promotes proliferation and invasion in oesophageal squamous cell carcinoma by targeting programmed cell death 4. Br. J. Cancer 2014, 111, 2003–2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, M.; Liu, R.; Li, X.; Liao, J.; Pu, Y.; Pan, E.; Yin, L.; Wang, Y. miRNA-183 suppresses apoptosis and promotes proliferation in esophageal cancer by targeting PDCD4. Mol. Cell. 2014, 37, 873–880. [Google Scholar] [CrossRef] [PubMed]
- Lin, X.; Zheng, L.; Song, H.; Xiao, J.; Pan, B.; Chen, H.; Jin, X.; Yu, H. Effects of microRNA-183 on epithelial-mesenchymal transition, proliferation, migration, invasion and apoptosis in human pancreatic cancer SW1900 cells by targeting MTA1. Exp. Mol. Pathol. 2017, 102, 522–532. [Google Scholar] [CrossRef] [PubMed]
- Shi, W.; Bruce, J.; Lee, M.; Yue, S.; Rowe, M.; Pintilie, M.; Kogo, R.; Bissey, P.A.; Fyles, A.; Yip, K.W.; et al. MiR-449a promotes breast cancer progression by targeting CRIP2. Oncotarget 2016, 7, 18906–18918. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, H.L.; Xiao, Y.; Wu, L.; Ma, D.C. Effects of miR-449a on proliferation and migration of human breast cancer cell line MCF-7. Zhongguo Ying Yong Sheng Li Xue Za Zhi 2017, 33, 508–513. (In Chinese) [Google Scholar] [PubMed]
- Bosman, F.T.; Carneiro, F.; Hruban, R.H.; Theise, N.D. WHO Classification of Tumours of the Digestive System, 4th ed.; World Health Organization: Geneva, Switzerland, 2010. [Google Scholar]
- Kemmerling, R.; Weyland, D.; Kiesslich, T.; Illig, R.; Klieser, E.; Jager, T.; Dietze, O.; Neureiter, D. Robust linear regression model of Ki-67 for mitotic rate in gastrointestinal stromal tumors. Oncol. Lett. 2014, 7, 745–749. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klieser, E.; Illig, R.; Stattner, S.; Primavesi, F.; Jager, T.; Swierczynski, S.; Kiesslich, T.; Kemmerling, R.; Bollmann, C.; Di Fazio, P.; et al. Endoplasmic reticulum stress in pancreatic neuroendocrine tumors is linked to clinicopathological parameters and possible epigenetic regulations. Anticancer Res. 2015, 35, 6127–6136. [Google Scholar] [PubMed]
- Kemmerling, R.; Alinger, B.; Dietze, O.; Bosmuller, H.C.; Ocker, M.; Wolkersdorfer, G.W.; Berr, F.; Neureiter, D.; Kiesslich, T. Association of stem cell marker expression pattern and survival in human biliary tract cancer. Int. J. Oncol. 2012, 41, 511–522. [Google Scholar] [CrossRef] [PubMed]
- Kiesslich, T.; Alinger, B.; Wolkersdorfer, G.W.; Ocker, M.; Neureiter, D.; Berr, F. Active Wnt signalling is associated with low differentiation and high proliferation in human biliary tract cancer in vitro and in vivo and is sensitive to pharmacological inhibition. Int. J. Oncol. 2010, 36, 49–58. [Google Scholar] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameters | Cases | p Value a |
|---|---|---|
| Gender, n (%) | ||
| Male | 32 (56.1%) | 0.354 |
| Female | 25 (43.9%) | |
| Age (mean ± SD) | 60.9 ± 14.4 | n.a. |
| Size (mean (cm) ± SD) | 2.69 ± 2.34 | n.a. |
| Localization, n (%) b | ||
| Head | 25 (44.6) | 0.067 |
| Corpus | 11 (19.6) | |
| Cauda | 20 (35.7) | |
| TNM, n c | ||
| T1/2/3/4 | 24/13/17/3 | ** 0.001 |
| N0/1 | 38/19 | * 0.012 |
| M0/1 | 46/11 | ** 0.000 |
| Grading, n (%) | ||
| G1 | 32 (56.1) | n.a. |
| G2 | 17 (29.8) | |
| G3 | 8 (14.0) | |
| R status, n (%) | ||
| R0 | 51 (89.5) | ** 0.000 |
| R1 | 5 (8.8) | |
| R2 | 1 (1.8) | |
| OS, n (%) | ||
| Yes | 47 (82.5) | ** 0.000 |
| No | 10 (17.5) | |
| DFS, n (%) | ||
| Yes | 34 (73.9) | ** 0.001 |
| No | 12 (26.1) | |
| Proliferation/Mitosis | ||
| H&E d | 12.8 ± 27.1 | n.a. |
| pHH3 e | 15.0 ± 31.4 | |
| Ki-67 (%) f | 10.8 ± 21.3 | |
| Hormone activity n (%) g | ||
| No | 30 (52.6) | 0.691 |
| Yes | 27 (47.4) | |
| miRNA- | Mean ± SD | Cut-Off-Value (ROC, Youden-Index) | |
|---|---|---|---|
| Overall-Survival | Disease-Free-Survival | ||
| 132-3p | 0.18826 ± 0.41903 | 0.26125 | 0.06320 |
| 145-5p | 1.0057 ± 1.14076 | 0.58855 | 0.39820 |
| 183-5p | 0.06794 ± 0.18228 | 0.05250 | 0.05250 |
| 34a-5p | 0.24186 ± 0.24647 | 0.08215 | 0.29910 |
| 449a | 0.00207 ± 0.00600 | 0.00310 | 0.00265 |
© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Klieser, E.; Urbas, R.; Swierczynski, S.; Stättner, S.; Primavesi, F.; Jäger, T.; Mayr, C.; Kiesslich, T.; Di Fazio, P.; Helm, K.; et al. HDAC-Linked “Proliferative” miRNA Expression Pattern in Pancreatic Neuroendocrine Tumors. Int. J. Mol. Sci. 2018, 19, 2781. https://doi.org/10.3390/ijms19092781
Klieser E, Urbas R, Swierczynski S, Stättner S, Primavesi F, Jäger T, Mayr C, Kiesslich T, Di Fazio P, Helm K, et al. HDAC-Linked “Proliferative” miRNA Expression Pattern in Pancreatic Neuroendocrine Tumors. International Journal of Molecular Sciences. 2018; 19(9):2781. https://doi.org/10.3390/ijms19092781
Chicago/Turabian StyleKlieser, Eckhard, Romana Urbas, Stefan Swierczynski, Stefan Stättner, Florian Primavesi, Tarkan Jäger, Christian Mayr, Tobias Kiesslich, Pietro Di Fazio, Katharina Helm, and et al. 2018. "HDAC-Linked “Proliferative” miRNA Expression Pattern in Pancreatic Neuroendocrine Tumors" International Journal of Molecular Sciences 19, no. 9: 2781. https://doi.org/10.3390/ijms19092781