Chalcone Derivatives 4′-Amino-1-Naphthyl-Chalcone (D14) and 4′-Amino-4-Methyl-1-Naphthyl-Chalcone (D15) Suppress Migration and Invasion of Osteosarcoma Cells Mediated by p53 Regulating EMT-Related Genes


 ,
,
Abstract
:
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
1. Introduction
2. Results
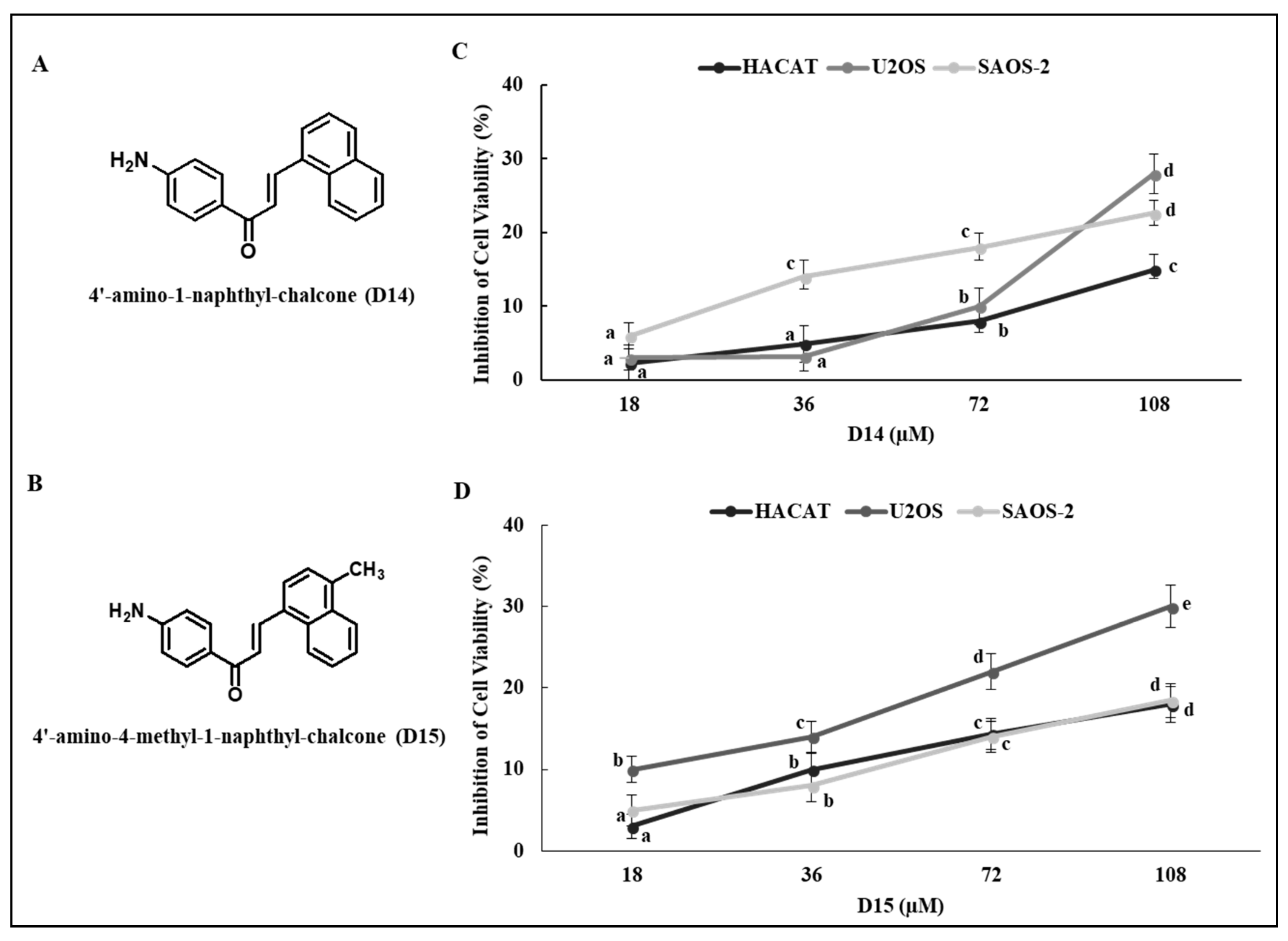
2.1. 4′-Aminochalcones Poorly Inhibit Viability of Osteosarcoma and Normal Cells
2.2. 4′-Aminochalcones Decrease Osteosarcoma Cell Migration and Invasiveness
2.3. 4′-Aminochalcones Attenuate the Expression and Proteolytic Activity of MMP-2 and MMP-9 in Osteosarcoma Cells
2.4. 4′-Aminochalcones Regulate EMT Gene Expression in Osteosarcoma Cells
3. Discussion
4. Materials and Methods
4.1. Cell Culture and Chemicals
4.2. Cell Viability Assay
4.3. In Vitro Cell Migration and Invasion Assay
4.4. Gelatin Zymography
4.5. Reverse Transcription-Polymerase Chain Reaction (RT-PCR)
4.6. Western Blot
4.7. Expression Vector and Transfection
4.8. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Kansara, M.; Teng, M.W.; Smyth, M.J.; Thomas, D.M. Translational biology of osteosarcoma. Nat. Rev. Cancer 2014, 14, 722–735. [Google Scholar] [CrossRef] [PubMed]
- Anderson, M.E. Update on Survival in Osteosarcoma. Orthop. Clin. N. Am. 2016, 47, 283–292. [Google Scholar] [CrossRef] [PubMed]
- Geller, D.S.; Gorlick, R. Osteosarcoma: A review of diagnosis, management, and treatment strategies. Clin. Adv. Hematol. Oncol. 2010, 8, 705–718. [Google Scholar] [PubMed]
- Kargozaran, H.; Yuan, S.Y.; Breslin, J.W.; Watson, K.D.; Gaudreault, N.; Breen, A.; Wu, M.H. A role for endothelial-derived matrix metalloproteinase-2 in breast cancer cell transmigration across the endothelial-basement membrane barrier. Clini. Exp. Metast. 2007, 24, 495–502. [Google Scholar] [CrossRef] [PubMed]
- Taniwaki, K.; Fukamachi, H.; Komori, K.; Ohtake, Y.; Nonaka, T.; Sakamoto, T.; Shiomi, T.; Okada, Y.; Itoh, T.; Itohara, S.; et al. Stroma-derived matrix metalloproteinase (MMP)-2 promotes membrane type 1-MMP-dependent tumor growth in mice. Cancer Res. 2007, 67, 4311–4319. [Google Scholar] [CrossRef] [PubMed]
- Webb, A.H.; Gao, B.T.; Goldsmith, Z.K.; Irvine, A.S.; Saleh, N.; Lee, R.P.; Lendermon, J.B.; Bheemreddy, R.; Zhang, Q.; Brennan, R.C.; et al. Inhibition of MMP-2 and MMP-9 decreases cellular migration, and angiogenesis in in vitro models of retinoblastoma. BMC Cancer 2017, 17, 434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luo, Y.; Liang, F.; Zhang, Z.-Y. PRL1 promotes cell migration and invasion by increasing MMP2 and MMP9 expression through Src and ERK1/2 pathways. Biochemistry 2009, 48, 1838–1846. [Google Scholar] [CrossRef] [PubMed]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [PubMed]
- Lee, S.-H.; Lee, S.-J.; Jung, Y.S.; Xu, Y.; Kang, H.S.; Ha, N.-C.; Park, B.-J. Blocking of p53-Snail Binding, Promoted by Oncogenic K-Ras, Recovers p53 Expression and Function. Neoplasia 2009, 11, 22–31. [Google Scholar] [CrossRef] [PubMed]
- Shiota, M.; Izumi, H.; Onitsuka, T.; Miyamoto, N.; Kashiwagi, E.; Kidani, A.; Hirano, G.; Takahashi, M.; Naito, S.; Kohno, K. Twist and p53 reciprocally regulate target genes via direct interaction. Oncogene 2008, 27, 5543–5553. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powell, E.; Piwnica-Worms, D.; Piwnica-Worms, H. Contribution of p53 to metastasis. Cancer Discov. 2014, 4, 405–414. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.; Teixeira Lima, F.; Seba, V.; Mendes Lourenco, A.L.; Lucas, T.G.; de Andrade, B.V.; Torrezan, G.S.; Polaquini, C.R.; Garcia, M.E.; Couto, L.B.; et al. Curcumin Analog CH-5 Suppresses the Proliferation, Migration, and Invasion of the Human Gastric Cancer Cell Line HGC-27. Molecules 2018, 23, 279. [Google Scholar] [CrossRef] [PubMed]
- Seba, V.; Silva, G.; Lima, F.T.; Santos, M.B.; Cechinel Filho, V.; Fachin, A.L.; Regasini, L.O.; Marins, M. Antiproliferative and anti-migratory activity of chalcones against osteosarcoma cells. Toxicol. In Vitro. submitted.
- Bortolotto, L.F.; Barbosa, F.R.; Silva, G.; Bitencourt, T.A.; Beleboni, R.O.; Baek, S.J.; Marins, M.; Fachin, A.L. Cytotoxicity of trans-chalcone and licochalcone A against breast cancer cells is due to apoptosis induction and cell cycle arrest. Biomed. Pharmacother. 2017, 85, 425–433. [Google Scholar] [CrossRef] [PubMed]
- Sikander, M.; Malik, S.; Yadav, D.; Biswas, S.; Katare, D.P.; Jain, S.K. Cytoprotective activity of a trans-chalcone against hydrogen peroxide induced toxicity in hepatocellular carcinoma (HepG2) cells. Asian Pac. J. Cancer Prev. 2011, 12, 2513–2516. [Google Scholar] [PubMed]
- Silva, G.; Fachin, A.L.; Beleboni, R.O.; Franca, S.C.; Marins, M. In vitro action of flavonoids in the canine malignant histiocytic cell line DH82. Molecules 2013, 18, 15448–15463. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.B.; Pinhanelli, V.C.; Garcia, M.A.R.; Silva, G.; Baek, S.J.; Franca, S.C.; Fachin, A.L.; Marins, M.; Regasini, L.O. Antiproliferative and pro-apoptotic activities of 2′- and 4′-aminochalcones against tumor canine cells. Eur. J. Med. Chem. 2017, 138, 884–889. [Google Scholar] [CrossRef] [PubMed]
- Silva, G.; Marins, M.; Fachin, A.L.; Lee, S.H.; Baek, S.J. Anti-cancer activity of trans-chalcone in osteosarcoma: Involvement of Sp1 and p53. Mol. Carcinog. 2016, 55, 1438–1448. [Google Scholar] [CrossRef] [PubMed]
- Lei, P.; He, H.; Hu, Y.; Liao, Z. Small interfering RNA-induced silencing of galectin-3 inhibits the malignant phenotypes of osteosarcoma in vitro. Mol. Med. Rep. 2015, 12, 6316–6322. [Google Scholar] [CrossRef] [PubMed]
- Jin, J.; Cai, L.; Liu, Z.M.; Zhou, X.S. miRNA-218 inhibits osteosarcoma cell migration and invasion by down-regulating of TIAM1, MMP2 and MMP9. Asian Pac. J. Cancer Prev. 2013, 14, 3681–3684. [Google Scholar] [CrossRef] [PubMed]
- Bjornland, K.; Flatmark, K.; Pettersen, S.; Aaasen, A.O.; Fodstad, O.; Maelandsmo, G.M. Matrix metalloproteinases participate in osteosarcoma invasion. J. Surg. Res. 2005, 127, 151–156. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zhang, X. Association of MMP-2 expression and prognosis in osteosarcoma patients. Int. J. Clin. Exp. Pathol. 2015, 8, 14965–14970. [Google Scholar] [PubMed]
- Li, H.; Zhang, K.; Liu, L.H.; Ouyang, Y.; Bu, J.; Guo, H.B.; Xiao, T. A systematic review of matrix metalloproteinase 9 as a biomarker of survival in patients with osteosarcoma. Tumour Biol. 2014, 35, 5487–5491. [Google Scholar] [CrossRef] [PubMed]
- Yang, G.; Yuan, J.; Li, K. EMT transcription factors: Implication in osteosarcoma. Med. Oncol. 2013, 30, 697. [Google Scholar] [CrossRef] [PubMed]
- Zhang, X.; Zhang, Y.; Mao, Y.; Ma, X. The lncRNA PCAT1 is correlated with poor prognosis and promotes cell proliferation, invasion, migration and EMT in osteosarcoma. Onco. Targets Ther. 2018, 11, 629–638. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.; Zhang, D.; Han, S.; Gao, P.; Liu, C.; Li, J.; Pan, X. Fibulin-3 promotes osteosarcoma invasion and metastasis by inducing epithelial to mesenchymal transition and activating the Wnt/β-catenin signaling pathway. Sci. Rep. 2017, 7, 6215. [Google Scholar] [CrossRef] [PubMed]
- Liu, D.D.; Kang, Y. Ets2 anchors the prometastatic function of mutant p53 in osteosarcoma. Genes Dev. 2017, 31, 1823–1824. [Google Scholar] [CrossRef] [PubMed]
- Ginat, D.; Schulte, J.; Gooi, Z.; Cipriani, N. High-Grade Conventional Osteosarcoma of the Mandible Associated With P53 Germline Mutation. J. Craniofac. Surg. 2018, 29, 738–740. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.; McDonnell, T.J.; Montes de Oca Luna, R.; Kapoor, M.; Mims, B.; El-Naggar, A.K.; Lozano, G. High metastatic potential in mice inheriting a targeted p53 missense mutation. Proc. Natl. Acad. Sci. USA 2000, 97, 4174–4179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, R.; Tian, K.; Wang, W.; Wang, L. p53 suppresses cell proliferation, metastasis, and angiogenesis of osteosarcoma through inhibition of the PI3K/AKT/mTOR pathway. Int. J. Surg. 2015, 20, 80–87. [Google Scholar] [CrossRef] [PubMed]
- Densmore, C.L.; Kleinerman, E.S.; Gautam, A.; Jia, S.F.; Xu, B.; Worth, L.L.; Waldrep, J.C.; Fung, Y.K.; T’Ang, A.; Knight, V. Growth suppression of established human osteosarcoma lung metastases in mice by aerosol gene therapy with PEI-p53 complexes. Cancer Gene Ther. 2001, 8, 619–627. [Google Scholar] [CrossRef] [PubMed]
- Nakase, M.; Inui, M.; Okumura, K.; Kamei, T.; Nakamura, S.; Tagawa, T. p53 gene therapy of human osteosarcoma using a transferrin-modified cationic liposome. Mol. Cancer Ther. 2005, 4, 625–631. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, T.; Veronese, A.; Pichiorri, F.; Lee, T.J.; Jeon, Y.J.; Volinia, S.; Pineau, P.; Marchio, A.; Palatini, J.; Suh, S.S.; et al. p53 regulates epithelial-mesenchymal transition through microRNAs targeting ZEB1 and ZEB2. J. Exp. Med. 2011, 208, 875–883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Lei, Y.; Gao, X.; Liang, Q.; Li, L.; Feng, J.; Hou, P.; Han, L.; Zhang, Y.; Huang, B.; et al. p53 Attenuates the oncogenic Ras-induced epithelial-mesenchymal transition in human mammary epithelial cells. Biochem. Biophys. Res. Commun. 2013, 434, 606–613. [Google Scholar] [CrossRef] [PubMed]
- Mahapatra, D.K.; Bharti, S.K.; Asati, V. Anti-cancer chalcones: Structural and molecular target perspectives. Eur. J. Med. Chem. 2015, 98, 69–114. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J.; Zhu, X.; Li, H.; Li, B.; Sun, L.; Xie, T.; Zhu, T.; Zhou, H.; Ye, Z. Piperine inhibits proliferation of human osteosarcoma cells via G2/M phase arrest and metastasis by suppressing MMP-2/-9 expression. Int. Immunopharmacol. 2015, 24, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.J.; Lee, T.S.; Park, J.B.; Park, K.K.; Choe, J.Y.; Sin, D.I.; Park, Y.Y.; Moon, Y.S.; Lee, K.G.; Yeo, J.H.; et al. Disulfiram suppresses invasive ability of osteosarcoma cells via the inhibition of MMP-2 and MMP-9 expression. J. Biochem. Mol. Biol. 2007, 40, 1069–1076. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.; Zhang, Z.; Liao, G.; Liu, S.; Zhang, Y.; Wen, J.; Zeng, L. Positive expression of LSD1 and negative expression of E-cadherin correlate with metastasis and poor prognosis of colon cancer. Dig. Dis. Sci. 2013, 58, 1581–1589. [Google Scholar]
- Nijkamp, M.M.; Span, P.N.; Hoogsteen, I.J.; van der Kogel, A.J.; Kaanders, J.H.; Bussink, J. Expression of E-cadherin and vimentin correlates with metastasis formation in head and neck squamous cell carcinoma patients. Radiother. Oncol. 2011, 99, 344–348. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Zhang, X.; Li, Z.; Chai, J.; Zhang, G.; Yu, Z.; Cheng, Y.; Hu, S. Overexpression of nuclear β-catenin at invasive front in rectal carcinoma is associated with lymph node metastasis and poor prognosis. Clin. Transl. Oncol. 2014, 16, 488–494. [Google Scholar] [CrossRef] [PubMed]
- Toiyama, Y.; Yasuda, H.; Saigusa, S.; Tanaka, K.; Inoue, Y.; Goel, A.; Kusunoki, M. Increased expression of Slug and Vimentin as novel predictive biomarkers for lymph node metastasis and poor prognosis in colorectal cancer. Carcinogenesis 2013, 34, 2548–2557. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nguyen, P.T.; Kudo, Y.; Yoshida, M.; Iizuka, S.; Ogawa, I.; Takata, T. N-cadherin expression is correlated with metastasis of spindle cell carcinoma of head and neck region. J. Oral. Pathol. Med. 2011, 40, 77–82. [Google Scholar] [CrossRef] [PubMed]
- Luo, Y.; Yu, T.; Zhang, Q.; Fu, Q.; Hu, Y.; Xiang, M.; Peng, H.; Zheng, T.; Lu, L.; Shi, H. Upregulated N-cadherin expression is associated with poor prognosis in epithelial-derived solid tumours: A meta-analysis. Eur. J. Clin. Investig. 2018, 48, e12903. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ferrari-Amorotti, G.; Chiodoni, C.; Shen, F.; Cattelani, S.; Soliera, A.R.; Manzotti, G.; Grisendi, G.; Dominici, M.; Rivasi, F.; Colombo, M.P.; et al. Suppression of invasion and metastasis of triple-negative breast cancer lines by pharmacological or genetic inhibition of slug activity. Neoplasia 2014, 16, 1047–1058. [Google Scholar] [CrossRef] [PubMed]
- Lai, T.Y.; Su, C.C.; Kuo, W.W.; Yeh, Y.L.; Kuo, W.H.; Tsai, F.J.; Tsai, C.H.; Weng, Y.J.; Huang, C.Y.; Chen, L.M. β-catenin plays a key role in metastasis of human hepatocellular carcinoma. Oncol. Rep. 2011, 26, 415–422. [Google Scholar] [PubMed]
- Tanaka, H.; Kono, E.; Tran, C.P.; Miyazaki, H.; Yamashiro, J.; Shimomura, T.; Fazli, L.; Wada, R.; Huang, J.; Vessella, R.L.; et al. Monoclonal antibody targeting of N-cadherin inhibits prostate cancer growth, metastasis and castration resistance. Nat. Med. 2010, 16, 1414–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, Y.C.; Bilen, M.A.; Yu, G.; Lin, S.C.; Huang, C.F.; Ortiz, A.; Cho, H.; Song, J.H.; Satcher, R.L.; Kuang, J.; et al. Inhibition of cell adhesion by a cadherin-11 antibody thwarts bone metastasis. Mol. Cancer Res. 2013, 11, 1401–1411. [Google Scholar] [CrossRef] [PubMed]
- Sharili, A.S.; Allen, S.; Smith, K.; Hargreaves, J.; Price, J.; McGonnell, I. Expression of Snail2 in long bone osteosarcomas correlates with tumour malignancy. Tumour Biol. 2011, 32, 515–526. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Walker, A.; Frei, R.; Lawson, K.R. The cytoplasmic domain of N-cadherin modulates MMP9 induction in oral squamous carcinoma cells. Int. J. Oncol. 2014, 45, 1699–1706. [Google Scholar] [CrossRef] [PubMed]
- Zhao, C.; Zhang, M.; Liu, W.; Wang, C.; Zhang, Q.; Li, W. β-Catenin knockdown inhibits pituitary adenoma cell proliferation and invasion via interfering with AKT and gelatinases expression. Int. J. Oncol. 2015, 46, 1643–1650. [Google Scholar] [CrossRef] [PubMed]
- Kim, N.H.; Kim, H.S.; Li, X.Y.; Lee, I.; Choi, H.S.; Kang, S.E.; Cha, S.Y.; Ryu, J.K.; Yoon, D.; Fearon, E.R.; et al. A p53/miRNA-34 axis regulates Snail1-dependent cancer cell epithelial-mesenchymal transition. J. Cell Biol. 2011, 195, 417–433. [Google Scholar] [CrossRef] [PubMed]
- Tseng, R.C.; Lin, R.K.; Wen, C.K.; Tseng, C.; Hsu, H.S.; Hsu, W.H.; Wang, Y.C. Epigenetic silencing of AXIN2/βTrCP and deregulation of p53-mediated control lead to wild-type β-catenin nuclear accumulation in lung tumorigenesis. Oncogene 2008, 27, 4488–4496. [Google Scholar] [CrossRef] [PubMed]
- Levina, E.; Oren, M.; Ben-Ze’ev, A. Downregulation of β-catenin by p53 involves changes in the rate of β-catenin phosphorylation and Axin dynamics. Oncogene 2004, 23, 4444–4453. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, D.; Wang, M.; Guo, W.; Zhao, X.; Tu, X.; Huang, S.; Zou, X.; Peng, X. Wild-type p53 suppresses the epithelial-mesenchymal transition and stemness in PC-3 prostate cancer cells by modulating miR145. Int. J. Oncol. 2013, 42, 1473–1481. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Ge, Q.; Zhang, Q.; Chen, Z.; Hu, J.; Li, F.; Ye, Z. Targeted p53 activation by saRNA suppresses human bladder cancer cells growth and metastasis. J. Exp. Clin. Cancer Res. 2016, 35, 53. [Google Scholar] [CrossRef] [PubMed]
- Kim, J.; Bae, S.; An, S.; Park, J.K.; Kim, E.M.; Hwang, S.G.; Kim, W.J.; Um, H.D. Cooperative actions of p21WAF1 and p53 induce Slug protein degradation and suppress cell invasion. EMBO Rep. 2014, 15, 1062–1068. [Google Scholar] [CrossRef] [PubMed] [Green Version]





© 2018 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Seba, V.; Silva, G.; Santos, M.B.d.; Baek, S.J.; França, S.D.C.; Fachin, A.L.; Regasini, L.O.; Marins, M. Chalcone Derivatives 4′-Amino-1-Naphthyl-Chalcone (D14) and 4′-Amino-4-Methyl-1-Naphthyl-Chalcone (D15) Suppress Migration and Invasion of Osteosarcoma Cells Mediated by p53 Regulating EMT-Related Genes. Int. J. Mol. Sci. 2018, 19, 2838. https://doi.org/10.3390/ijms19092838
Seba V, Silva G, Santos MBd, Baek SJ, França SDC, Fachin AL, Regasini LO, Marins M. Chalcone Derivatives 4′-Amino-1-Naphthyl-Chalcone (D14) and 4′-Amino-4-Methyl-1-Naphthyl-Chalcone (D15) Suppress Migration and Invasion of Osteosarcoma Cells Mediated by p53 Regulating EMT-Related Genes. International Journal of Molecular Sciences. 2018; 19(9):2838. https://doi.org/10.3390/ijms19092838
Chicago/Turabian StyleSeba, Viviane, Gabriel Silva, Mariana Bastos dos Santos, Seung Joon Baek, Suzelei De Castro França, Ana Lúcia Fachin, Luis Octavio Regasini, and Mozart Marins. 2018. "Chalcone Derivatives 4′-Amino-1-Naphthyl-Chalcone (D14) and 4′-Amino-4-Methyl-1-Naphthyl-Chalcone (D15) Suppress Migration and Invasion of Osteosarcoma Cells Mediated by p53 Regulating EMT-Related Genes" International Journal of Molecular Sciences 19, no. 9: 2838. https://doi.org/10.3390/ijms19092838