MiR-125b-2 Knockout in Testis Is Associated with Targeting to the PAP Gene, Mitochondrial Copy Number, and Impaired Sperm Quality


Abstract
1. Introduction
2. Result
2.1. Phenotype of KO Mice
2.2. miR-125b-2 Knockout Causes Infertility
2.3. RNA-Seq Analysis Revealed the Changes in the Mouse Transcriptome between the KO and WT
2.4. Sperm mtDNA Copy and Integrity
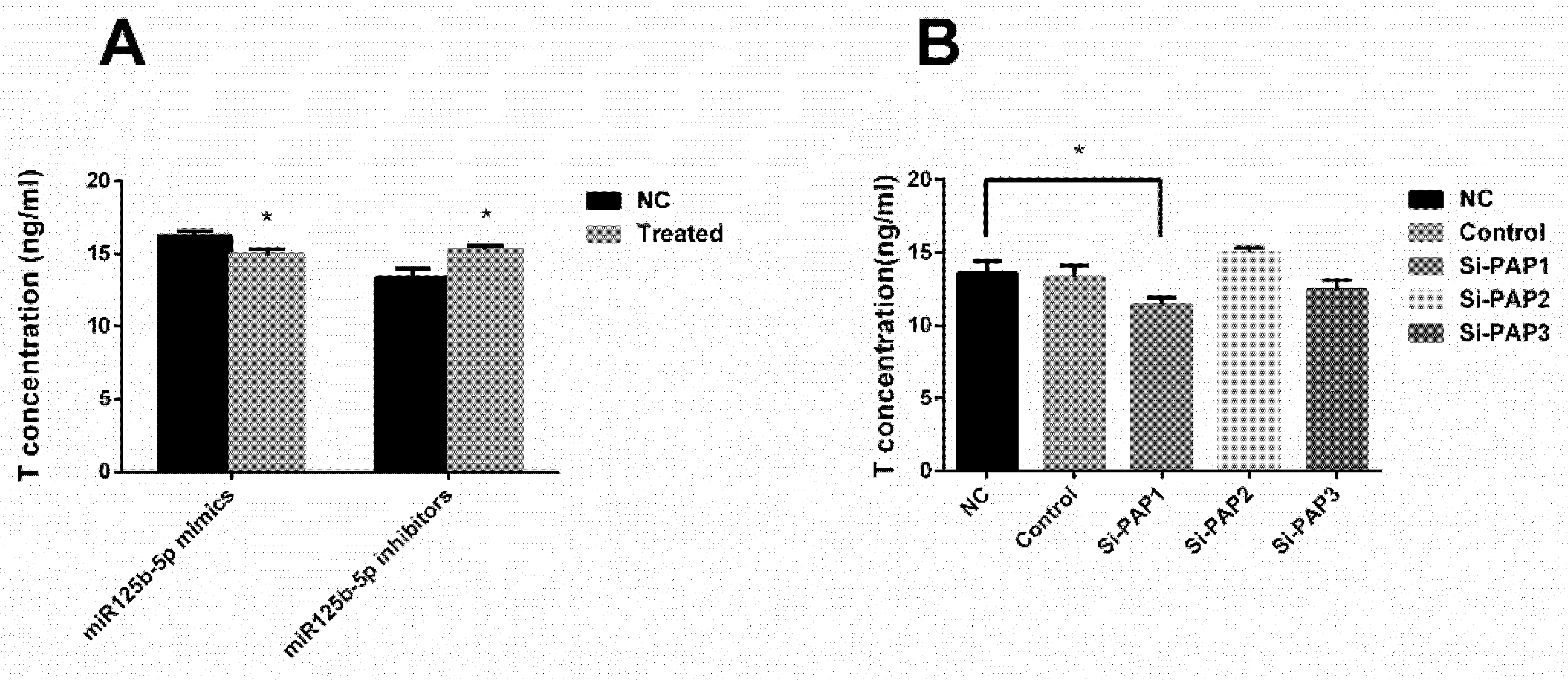
2.5. miR-125b-5p Targeted Papolb (PAP) Gene to Suppress Testosterone Secretion in TM3 Cells
2.5.1. miR-125b-5p Targeted on PAP
2.5.2. Both miR-125b-2 and PAP Affect the Secretion of Testosterone in TM3
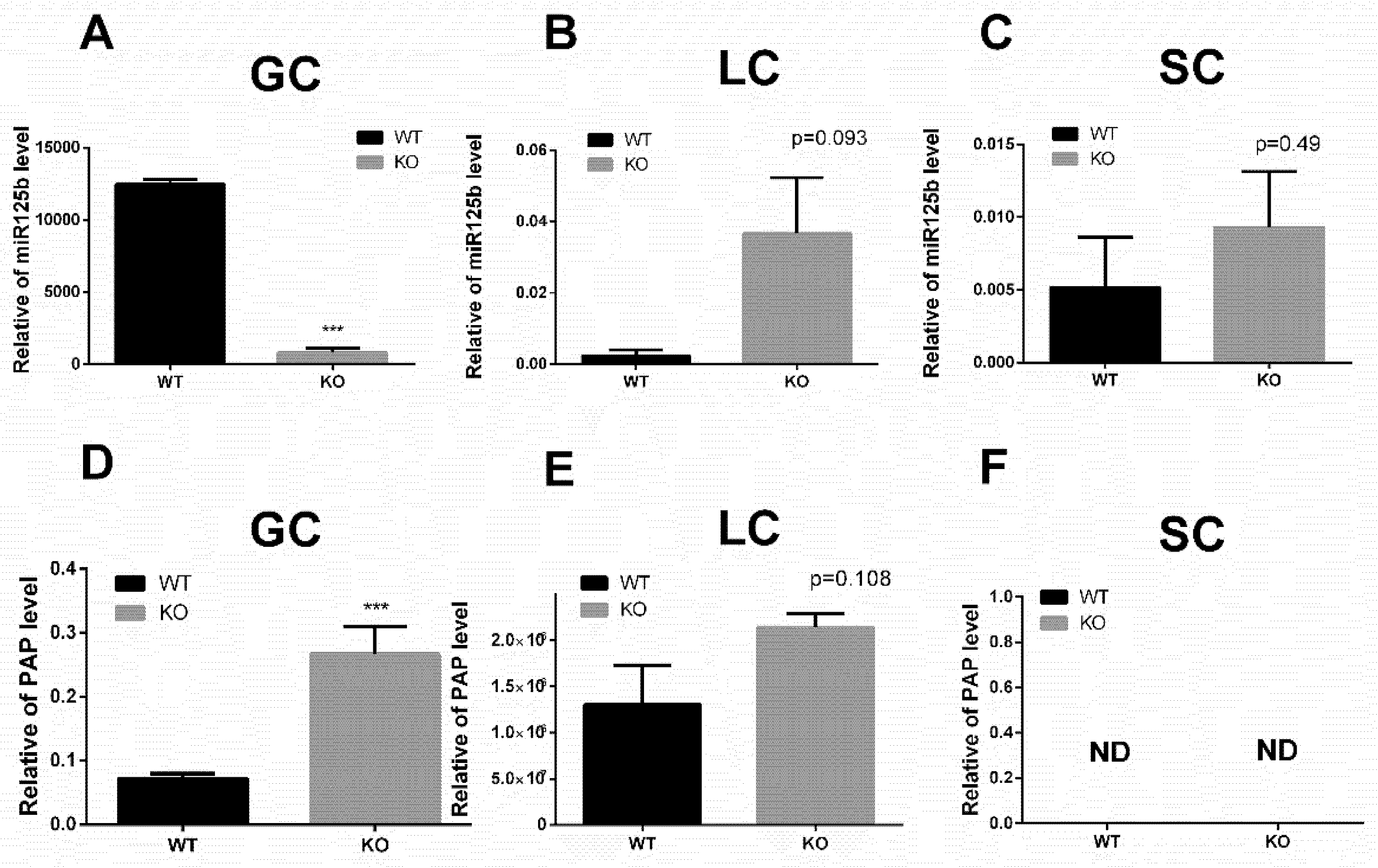
2.5.3. The Localization of miR-125b and PAP in Testicular Cells
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Fertility Test
4.3. Body Weight Measurement
4.4. Morphological Observation of Testis
4.5. Sperm Quality Evaluation
4.6. Target Prediction and Pathway Analysis
4.7. Protein Extraction and Western blot
4.8. Plasmid Construction, Transfection, and Luciferase Assay
4.9. RNA Extraction and qRT-PCR
4.10. Sperm mtDNA Integrity
4.11. Sperm mtDNA Mutations of mt-Cytb and mt-ATP6
4.12. siRNA Transfection
4.13. Cell Isolation and Purification
4.14. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Boivin, J.; Bunting, L.; Collins, J.A.; Nygren, K.G. International estimates of infertility prevalence and treatment-seeking: Potential need and demand for infertility medical care. Hum. Reprod. 2007, 22, 1506–1512. [Google Scholar] [CrossRef] [PubMed]
- Bakloutigargouri, S.; Ghorbel, M.; Ben, M.A.; Mkaouarrebai, E.; Cherif, M.; Chakroun, N.; Sellami, A.; Fakhfakh, F.; Ammarkeskes, L. A novel m.6307A>G mutation in the mitochondrial COXI gene in asthenozoospermic infertile men. Mol. Reprod. Dev. 2013, 80, 581–587. [Google Scholar] [CrossRef] [PubMed]
- Spiropoulos, J.; Turnbull, D.M.; Chinnery, P.F. Can mitochondrial DNA mutations cause sperm dysfunction? Mol. Hum. Reprod. 2002, 8, 719–721. [Google Scholar] [CrossRef] [PubMed]
- Ohta, H.; Tohda, A.; Nishimune, Y. Proliferation and Differentiation of Spermatogonial Stem Cells in the W/Wv Mutant Mouse Testis1. Biol. Reprod. 2003, 69, 1815–1821. [Google Scholar] [CrossRef] [PubMed]
- Russell, L.D.; Ettlin, R.A.; Hikim, A.P.S.; Clegg, E.D. Histological and Histopathological Evaluation of the Testis. Int. J. Androl. 2010, 16, 83. [Google Scholar] [CrossRef]
- Slezak, R.; Sasiadek, M. Chromosome Y microdeletions in the pathogenesis of male infertility. Polski Merkuriusz Lekarski Organ Polskiego Towarzystwa Lekarskiego 2002, 13, 229. [Google Scholar] [PubMed]
- Guo, T.; Qin, Y.; Gao, X.; Chen, H.; Li, G.; Ma, J.; Chen, Z.J. The role of male chromosomal polymorphism played in spermatogenesis and the outcome of IVF/ICSI-ET treatment. Int. J. Androl. 2012, 35, 802–809. [Google Scholar] [CrossRef] [PubMed]
- He, Z.; Kokkinaki, M.; Pant, D.; Gallicano, G.I.; Dym, M. Small RNA molecules in the regulation of spermatogenesis. Reproduction 2009, 137, 901–911. [Google Scholar] [CrossRef]
- Fabian, M.R.; Sonenberg, N.; Filipowicz, W. Regulation of mRNA translation and stability by microRNAs. Annu. Rev. Biochem. 2010, 79, 351–379. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Graham, I.; Hastings, R.; Gunewardena, S.; Brinkmeier, M.L.; Conn, P.M.; Camper, S.A.; Kumar, T.R. Gonadotrope-Specific Deletion of Dicer Results in Severely Suppressed Gonadotropins and Fertility Defects. J. Biol. Chem. 2015, 290, 2699–2714. [Google Scholar] [CrossRef]
- Wang, H.; Hastings, R.; Miller, W.L.; Kumar, T.R. Fshb-iCre mice are efficient and specific Cre deleters for the gonadotrope lineage. Mol. Cell. Endocrinol. 2016, 419, 124–138. [Google Scholar] [CrossRef]
- Ying, X.; Wei, K.; Lin, Z.; Cui, Y.; Ding, J.; Chen, Y.; Xu, B. MicroRNA-125b Suppresses Ovarian Cancer Progression via Suppression of the Epithelial-Mesenchymal Transition Pathway by Targeting the SET Protein. Cell. Physiol. Biochem. 2016, 39, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Solozobova, V.; Blattner, C. Regulation of p53 in embryonic stem cells. Exp. Cell Res. 2010, 316, 2434–2446. [Google Scholar] [CrossRef] [PubMed]
- Cheng, X.; Xi, Q.Y.; Wei, S.; Wu, D.; Ye, R.S.; Chen, T.; Qi, Q.E.; Jiang, Q.Y.; Wang, S.B.; Wang, L.N. Critical role of miR-125b in lipogenesis by targeting stearoyl-CoA desaturase-1 (SCD-1). J. Anim. Sci. 2016, 94, 65. [Google Scholar] [CrossRef] [PubMed]
- Kim, K.H.; Seo, Y.M.; Kim, E.Y.; Lee, S.Y.; Kwon, J.; Ko, J.J.; Lee, K.A. The miR-125 family is an important regulator of the expression and maintenance of maternal effect genes during preimplantational embryo development. Open Biol. 2016, 6. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Wang, C.L. Effects of testosterone on PPARγ and P450arom expression in polycystic ovary syndrome patients and related mechanisms. Eur. Rev. Med. Pharmacol. Sci. 2018, 22, 1549–1553. [Google Scholar] [PubMed]
- Frazee, A.C.; Pertea, G.; Jaffe, A.E.; Langmead, B.; Salzberg, S.L.; Leek, J.T. Flexible isoform-level differential expression analysis with Ballgown. bioRxiv 2014. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling The False Discovery Rate—A Practical And Powerful Approach To Multiple Testing. J. R. Stat. Soc. 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Ekstrand, M.I.; Maria, F.; Anja, R.; Bae, P.C.; Martina, G.; Kjell, H.; Pierre, R.; Gustafsson, C.M.; Larsson, N.G. Mitochondrial transcription factor A regulates mtDNA copy number in mammals. Hum. Mol. Genet. 2004, 13, 935–944. [Google Scholar] [CrossRef]
- Sun, Z.; Xue, X.; Zhang, Y.; Niu, R.; Wang, J. Effect of sodium fluoride on the sperm mitochondrial DNA in mice. Biochem. Biophys. Res. Commun. 2017, 492, 295–299. [Google Scholar] [CrossRef]
- Abhari, A.; Zarghami, N.; Shahnazi, V.; Barzegar, A.; Farzadi, L.; Karami, H.; Zununi, V.S.; Nouri, M. Significance of microRNA targeted estrogen receptor in male fertility. Iran. J. Basic Med. Sci. 2014, 17, 81–86. [Google Scholar]
- Zhang, Q.; Wang, Q.; Zhang, Y.; Cheng, S.; Hu, J.; Ma, Y.; Zhao, X. Comprehensive Analysis of MicroRNA(-)Messenger RNA from White Yak Testis Reveals the Differentially Expressed Molecules Involved in Development and Reproduction. Int. J. Mol. Sci. 2018, 19, 3083. [Google Scholar] [CrossRef] [PubMed]
- Ram Prakash, Y.; Noora, K. Small RNAs in spermatogenesis. Mol. Cell. Endocrinol. 2014, 382, 498–508. [Google Scholar]
- Said, T.M.; Agarwal, A.; Sharma, R.K.; Mascha, E.; Sikka, S.C.; Thomas, T.A., Jr. Human sperm superoxide anion generation and correlation with semen quality in patients with male infertility. Fertil. Steril. 2004, 82, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Peña, F.J.; Martínez, H.R.; Tapia, J.A.; Ferrusola, C.O.; Fernández, L.G.; García, B.M. Mitochondria in Mammalian Sperm Physiology and Pathology: A Review. Reprod. Domest. Anim. 2010, 44, 345–349. [Google Scholar] [CrossRef] [PubMed]
- Darr, C.R.; Moraes, L.E.; Connon, R.E.; Love, C.C.; Teague, S.; Varner, D.D.; Meyers, S.A. The relationship between mitochondrial DNA copy number and stallion sperm function. Theriogenology 2017, 94, 94–99. [Google Scholar] [CrossRef] [PubMed]
- Larsson, N.G.; Wang, J.; Wilhelmsson, H.; Oldfors, A.; Rustin, P.; Lewandoski, M.; Barsh, G.S.; Clayton, D.A. Mitochondrial transcription factor A is necessary for mtDNA maintenance and embryogenesis in mice. Nat. Genet. 1998, 18, 231. [Google Scholar] [CrossRef]
- Chan, C.C.; Liu, V.W.; Lau, E.Y.; Yeung, W.S.; Ng, E.H.; Ho, P.C. Mitochondrial DNA content and 4977 bp deletion in unfertilized oocytes. Mol. Hum. Reprod. 2006, 11, 843–846. [Google Scholar] [CrossRef]
- He, L.; Chinnery, P.F.; Durham, S.E.; Blakely, E.L.; Wardell, T.M.; Borthwick, G.M.; Taylor, R.W.; Turnbull, D.M. Detection and quantification of mitochondrial DNA deletions in individual cells by real-time PCR. Nucleic Acids Res. 2002, 30, e68. [Google Scholar] [CrossRef]
- Kashiwabara, S.; Noguchi, J.; Zhuang, T.; Ohmura, K.; Honda, A.; Sugiura, S.; Miyamoto, K.; Takahashi, S.; Inoue, K.; Ogura, A. Regulation of spermatogenesis by testis-specific, cytoplasmic poly(A) polymerase TPAP. Science 2002, 298, 1999–2002. [Google Scholar] [CrossRef]
- Zhuang, T.; Kashiwabara, S.; Noguchi, J.; Baba, T. Transgenic expression of testis-specific poly (A) polymerase TPAP in wild-type and TPAP-deficient mice. J. Reprod. Dev. 2004, 50, 207–213. [Google Scholar] [CrossRef] [PubMed]
- Kashiwabara, S.I.; Tsuruta, S.; Okada, K.; Yamaoka, Y.; Baba, T. Adenylation by testis-specific cytoplasmic poly(A) polymerase, PAPOLB/TPAP, is essential for spermatogenesis. J. Reprod. Dev. 2016, 62, 607–614. [Google Scholar] [CrossRef] [PubMed]
- Kashiwabara, S.; Yamagata, K.N.J.; Fukamizu, A.; Baba, T.; Zhuang, T.G. Identification of a novel isoform of poly (A) polymerase, TPAP, specifically present in the cytoplasm of spermatogenic cells. Dev. Biol. 2000, 228, 106–115. [Google Scholar] [CrossRef] [PubMed]
- Zhao, W.; Manley, J.L. Deregulation of poly(A) polymerase interferes with cell growth. Mol. Cell. Biol. 1998, 18, 5010–5020. [Google Scholar] [CrossRef] [PubMed]
- Juge, F.; Zaessinger, S.; Temme, C.; Wahle, E.; Simonelig, M. Control of poly(A) polymerase level is essential to cytoplasmic polyadenylation and early development in Drosophila. EMBO J. 2014, 21, 6603–6613. [Google Scholar] [CrossRef]
- Wdowiak, A.; Raczkiewicz, D.; Stasiak, M.; Bojar, I. Levels of FSH, LH and testosterone, and sperm DNA fragmentation. Neuro Endocrinol. Lett. 2014, 35, 73. [Google Scholar] [PubMed]
- Renato, F.; Suslik, Z.D.; C, E.S. Hypogonadotropic Hypogonadism Revisited. Clinics 2013, 68 (Suppl. 1), 81–88. [Google Scholar]
- Bhasin, S.; Cunningham, G.R.; Hayes, F.J.; Matsumoto, A.M.; Snyder, P.J.; Swerdloff, R.S.; Montori, V.M. Testosterone therapy in adult men with androgen deficiency syndromes: An endocrine society clinical practice guideline. J. Clin. Endocrinol. Metab. 2006, 91, 2536–2559. [Google Scholar] [CrossRef]
- Veeramachaneni, D.N.; Ott, R.S.; Heath, E.H.; Mcentee, K.; Bolt, D.J.; Hixon, J.E. Pathophysiology of small testes in beef bulls: Relationship between scrotal circumference, histopathologic features of testes and epididymides, seminal characteristics, and endocrine profiles. Am. J. Vet. Res. 1986, 47, 1988–1999. [Google Scholar] [PubMed]
- Babu, S.R.; Sadhnani, M.D.; Swarna, M.; Padmavathi, P.; Reddy, P.P. Evaluation of FSH, LH and testosterone levels in different subgroups of infertile males. Indian J. Clin. Biochem. 2004, 19, 45–49. [Google Scholar] [CrossRef]
- Ferlin, A.; Foresta, C. Infertility, Male; Springer: Berlin/Heidelberg, Germany, 2009. [Google Scholar]
- Maričić, A.; Štifter, S.; Valenčić, M.; Ðorđević, G.; Markić, D.; Španjol, J.; Sotošek, S.; Fučkar, Ž. Primary testicular necrotizing vasculitis clinically presented as neoplasm of the testicle: A case report. World J. Surg. Oncol. 2011, 9, 63. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Sood, S.; Arora, B.; Singh, M.; Beena; Roy, P.S. To Study the Effect of Vitamin D and E on Sodium-Fluoride-induced Toxicity in Reproductive Functions of Male Rabbits. Toxicol. Int. 2012, 19, 182–187. [Google Scholar] [CrossRef] [PubMed]
- Lewis, B.P.; Shih, I.-H.; Jones-Rhoades, M.W.; Bartel, D.P.; Burge, C.B. Prediction of Mammalian MicroRNA Targets. Cell 2003, 115, 787–798. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Song, G.J.; Lewis, V. Mitochondrial DNA integrity and copy number in sperm from infertile men. Fertil. Steril. 2008, 90, 2238–2244. [Google Scholar] [CrossRef] [PubMed]
- Mao, G.H.; Wang, Y.N.; Xu, M.; Wang, W.L.; Tan, L.; Tao, S.B. Polymorphisms in the MT-ATP6 and MT-CYB genes in in vitro fertilization failure. Mitochondrial DNA 2015, 26, 20–24. [Google Scholar] [CrossRef] [PubMed]
- Shang, T.; Zhang, X.; Wang, T.; Sun, B.; Deng, T.; Han, D. Toll-like receptor-initiated testicular innate immune responses in mouse Leydig cells. Endocrinology 2011, 152, 2827. [Google Scholar] [CrossRef]
- Yu, W.; Zheng, H.; Lin, W.; Tajima, A.; Zhang, Y.; Zhang, X.; Zhang, H.; Wu, J.; Han, D.; Rahman, N.A. Estrogen promotes Leydig cell engulfment by macrophages in male infertility. J. Clin. Investig. 2014, 124, 2709–2721. [Google Scholar] [CrossRef]
- Kim, G.J.; Georg, I.; Scherthan, H.; Merkenschlager, M.; Guillou, F.; Scherer, G.; Barrionuevo, F. Dicer is required for Sertoli cell function and survival. Int. J. Dev. Biol. 2015, 54, 867–875. [Google Scholar] [CrossRef]
- Chang, Y.F.; Lee-Chang, J.S.; Panneerdoss, S.; Ii, M.L.; Rao, M.K. Isolation of Sertoli, Leydig, and spermatogenic cells from the mouse testis. Biotechniques 2011, 51, 341. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | BW (g) | TW (mg) |
|---|---|---|
| WT | 29.12 ± 0.78 | 166.50 ± 8.43 |
| KO | 28.09 ± 0.89 | 167.50 ± 6.38 |
| Gene | Primer Sequences | Accession No. | Product Sizes (bp) |
|---|---|---|---|
| mtDNA-1 | F:GTTAATGTAGCTTAATAACAAAGCAAAGC | NC_005089.1 | 8220 |
| R:TAGTTGGGTAGTAGGTGTAAATGTATGTG | |||
| mtDNA-2 | F:ATTGGATCAACAAATCTCCTAGG | NC_005089.1 | 8140 |
| R:TTGTTAATGTTTATTGCGTAATAGAGTATG |
| Gene | Primer Sequences | Gene ID | Product Sizes (bp) |
|---|---|---|---|
| mt-Cytb | F:ATGACAAACATACGAAAAACACA | R17711 | 1144 |
| R:ATGGATATAATTTTAGTATTTTGTCTTCGA | |||
| mt-ATP6 | F: ATGAACGAAAATCTATTTGCCTC | 17705 | 681 |
| R:TTATGTATTATCATGTAGATATAGGCTTACTAGGA |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, L.; Zhu, Y.; Chen, T.; Sun, J.; Luo, J.; Shu, G.; Wang, S.; Zhu, X.; Jiang, Q.; Zhang, Y.; et al. MiR-125b-2 Knockout in Testis Is Associated with Targeting to the PAP Gene, Mitochondrial Copy Number, and Impaired Sperm Quality. Int. J. Mol. Sci. 2019, 20, 148. https://doi.org/10.3390/ijms20010148
Li L, Zhu Y, Chen T, Sun J, Luo J, Shu G, Wang S, Zhu X, Jiang Q, Zhang Y, et al. MiR-125b-2 Knockout in Testis Is Associated with Targeting to the PAP Gene, Mitochondrial Copy Number, and Impaired Sperm Quality. International Journal of Molecular Sciences. 2019; 20(1):148. https://doi.org/10.3390/ijms20010148
Chicago/Turabian StyleLi, Longlong, Yanling Zhu, Ting Chen, Jiajie Sun, Junyi Luo, Gang Shu, Songbo Wang, Xiaotong Zhu, Qingyan Jiang, Yongliang Zhang, and et al. 2019. "MiR-125b-2 Knockout in Testis Is Associated with Targeting to the PAP Gene, Mitochondrial Copy Number, and Impaired Sperm Quality" International Journal of Molecular Sciences 20, no. 1: 148. https://doi.org/10.3390/ijms20010148
APA StyleLi, L., Zhu, Y., Chen, T., Sun, J., Luo, J., Shu, G., Wang, S., Zhu, X., Jiang, Q., Zhang, Y., & Xi, Q. (2019). MiR-125b-2 Knockout in Testis Is Associated with Targeting to the PAP Gene, Mitochondrial Copy Number, and Impaired Sperm Quality. International Journal of Molecular Sciences, 20(1), 148. https://doi.org/10.3390/ijms20010148