Melatonin MT1 and MT2 Receptors Exhibit Distinct Effects in the Modulation of Body Temperature across the Light/Dark Cycle

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
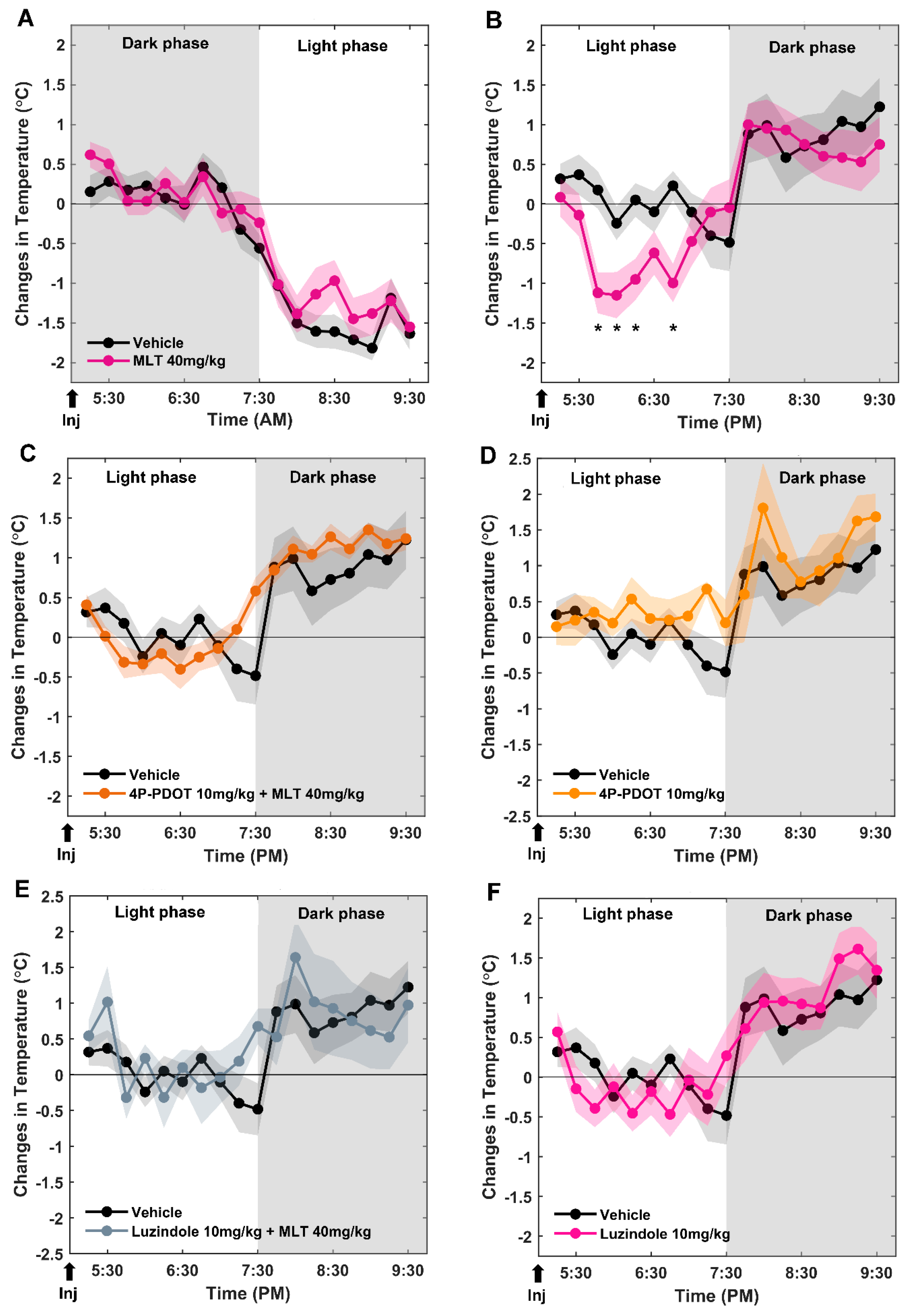
2.1. Effects of MLT Injected at the End of the Dark and of the Light Phases on Tb
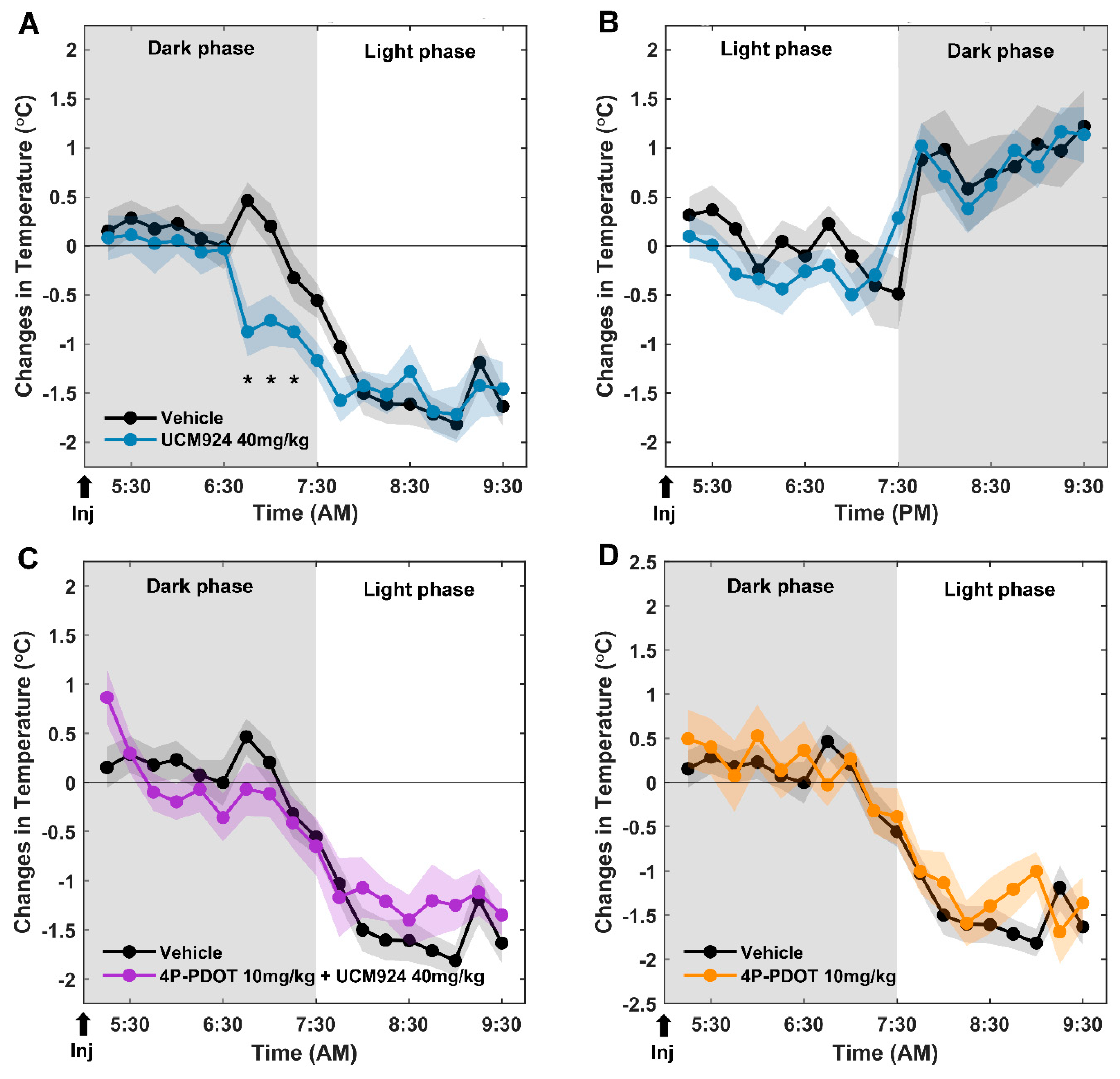
2.2. Effects of the Selective MT2 Partial Agonist UCM924 Injected at the End of the Dark and of the Light Phases on Tb
2.3. Effects of the Selective MT1 Partial Agonist UCM871 Injected at the End of the Dark and of the Light Phases on Tb
2.4. Effects of the Selective MT3 Agonist GR135531 and Prazosin Injected at the End of the Light Phase on Tb
3. Discussion
4. Materials and Methods
4.1. Animals
4.2. Drugs and Pharmacological Treatments
4.3. Assessment of Body Temperature
4.4. Statistical Analysis
Author Contributions
Funding
Conflicts of Interest
References
- Krauchi, K.; Deboer, T. The interrelationship between sleep regulation and thermoregulation. Front. Biosci 2010, 15, 604–625. [Google Scholar] [CrossRef]
- Boulant, J.A. Role of the preoptic-anterior hypothalamus in thermoregulation and fever. Clin. Infect. Dis. 2000, 31, S157–S161. [Google Scholar] [CrossRef] [PubMed]
- Boulant, J.A. Counterpoint: Heat-induced membrane depolarization of hypothalamic neurons: An unlikely mechanism of central thermosensitivity. Am. J. Physiol. Regul. Integr. Comp Physiol. 2006, 290, R1481–R1484. [Google Scholar] [PubMed]
- Zhao, Z.D.; Yang, W.Z.; Gao, C.; Fu, X.; Zhang, W.; Zhou, Q.; Chen, W.; Ni, X.; Lin, J.K.; Yang, J.; et al. A hypothalamic circuit that controls body temperature. Proc. Natl. Acad. Sci. USA 2017, 114, 2042–2047. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liedtke, W.B. Deconstructing mammalian thermoregulation. Proc. Natl. Acad. Sci. USA 2017, 114, 1765–1767. [Google Scholar] [CrossRef] [Green Version]
- Krauchi, K.; Cajochen, C.; Pache, M.; Flammer, J.; Wirz-Justice, A. Thermoregulatory effects of melatonin in relation to sleepiness. Chronobiol. Int. 2006, 23, 475–484. [Google Scholar] [CrossRef] [PubMed]
- Ng, K.Y.; Leong, M.K.; Liang, H.; Paxinos, G. Melatonin receptors: Distribution in mammalian brain and their respective putative functions. Brain Struct. Funct. 2017, 222, 2921–2939. [Google Scholar] [CrossRef]
- Benstaali, C.; Mailloux, A.; Bogdan, A.; Auzeby, A.; Touitou, Y. Circadian rhythms of body temperature and motor activity in rodents their relationships with the light-dark cycle. Life Sci. 2001, 68, 2645–2656. [Google Scholar] [CrossRef]
- Krauchi, K.; Wirz-Justice, A. Circadian rhythm of heat production, heart rate, and skin and core temperature under unmasking conditions in men. Am. J. Physiol. 1994, 267, R819–R829. [Google Scholar] [CrossRef] [PubMed]
- Dubocovich, M.L.; Markowska, M. Functional MT1 and MT2 melatonin receptors in mammals. Endocrine 2005, 27, 101–110. [Google Scholar] [CrossRef]
- Lacoste, B.; Angeloni, D.; Dominguez-Lopez, S.; Calderoni, S.; Mauro, A.; Fraschini, F.; Descarries, L.; Gobbi, G. Anatomical and cellular localization of melatonin MT1 and MT2 receptors in the adult rat brain. J. Pineal. Res. 2015, 58, 397–417. [Google Scholar] [CrossRef] [PubMed]
- Pandi-Perumal, S.R.; Srinivasan, V.; Maestroni, G.J.; Cardinali, D.P.; Poeggeler, B.; Hardeland, R. Melatonin: Nature’s most versatile biological signal? FEBS J. 2006, 273, 2813–2838. [Google Scholar] [CrossRef] [PubMed]
- Wu, Y.H.; Zhou, J.N.; Balesar, R.; Unmehopa, U.; Bao, A.; Jockers, R.; Van Heerikhuize, J.; Swaab, D.F. Distribution of MT1 melatonin receptor immunoreactivity in the human hypothalamus and pituitary gland: Colocalization of MT1 with vasopressin, oxytocin, and corticotropin-releasing hormone. J. Comp. Neurol. 2006, 499, 897–910. [Google Scholar] [CrossRef] [PubMed]
- Claustrat, B.; Leston, J. Melatonin: Physiological effects in humans. Neurochirurgie 2015, 61, 77–84. [Google Scholar] [CrossRef] [PubMed]
- Arendt, J. Melatonin. Clin. Endocrinol. 1988, 29, 205–229. [Google Scholar] [CrossRef]
- Klein, D.C.; Moore, R.Y.; Reppert, S.M. Suprachiasmatic Nucleus: The Mind’s Clock; Oxford University Press: Oxford, UK, 1991. [Google Scholar]
- Hardeland, R.; Poeggeler, B. Melatonin and synthetic melatonergic agonists: Actions and metabolism in the central nervous system. Cent. Nerv. Syst. Agents Med. Chem. 2012, 12, 189–216. [Google Scholar] [CrossRef] [PubMed]
- Comai, S.; Gobbi, G. Unveiling the role of melatonin MT2 receptors in sleep, anxiety and other neuropsychiatric diseases: A novel target in psychopharmacology. J. Psychiatry Neurosci. 2014, 39, 6–21. [Google Scholar] [CrossRef] [PubMed]
- Gobbi, G.; Comai, S. Differential Function of Melatonin MT1 and MT2 Receptors in REM and NREM Sleep. Front. Endocrinol (Lausanne) 2019, 10, 87. [Google Scholar] [CrossRef]
- Gobbi, G.; Comai, S. Sleep well. Untangling the role of melatonin MT1 and MT2 receptors in sleep. J. Pineal Res. 2019, 66, e12544. [Google Scholar] [CrossRef]
- Comai, S.; Ochoa-Sanchez, R.; Gobbi, G. Sleep-wake characterization of double MT(1)/MT(2) receptor knockout mice and comparison with MT(1) and MT(2) receptor knockout mice. Behav. Brain Res. 2013, 243, 231–238. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Comai, S.; Lacoste, B.; Bambico, F.R.; Dominguez-Lopez, S.; Spadoni, G.; Rivara, S.; Bedini, A.; Angeloni, D.; Fraschini, F.; et al. Promotion of non-rapid eye movement sleep and activation of reticular thalamic neurons by a novel MT2 melatonin receptor ligand. J. Neurosci. 2011, 31, 18439–18452. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Comai, S.; Spadoni, G.; Bedini, A.; Tarzia, G.; Gobbi, G. Melatonin, selective and non-selective MT1/MT2 receptors agonists: Differential effects on the 24-h vigilance states. Neurosci. Lett. 2014, 561, 156–161. [Google Scholar] [CrossRef]
- Ochoa-Sanchez, R.; Rainer, Q.; Comai, S.; Spadoni, G.; Bedini, A.; Rivara, S.; Fraschini, F.; Mor, M.; Tarzia, G.; Gobbi, G. Anxiolytic effects of the melatonin MT(2) receptor partial agonist UCM765: Comparison with melatonin and diazepam. Prog. Neuropsychopharmacol. Biol. Psychiatry 2012, 39, 318–325. [Google Scholar] [CrossRef]
- Lopez-Canul, M.; Comai, S.; Dominguez-Lopez, S.; Granados-Soto, V.; Gobbi, G. Antinociceptive properties of selective MT(2) melatonin receptor partial agonists. Eur. J. Pharmacol 2015, 764, 424–432. [Google Scholar] [CrossRef] [PubMed]
- Lopez-Canul, M.; Palazzo, E.; Dominguez-Lopez, S.; Luongo, L.; Lacoste, B.; Comai, S.; Angeloni, D.; Fraschini, F.; Boccella, S.; Spadoni, G.; et al. Selective melatonin MT2 receptor ligands relieve neuropathic pain through modulation of brainstem descending antinociceptive pathways. Pain 2015, 156, 305–317. [Google Scholar] [CrossRef]
- Pevet, P. Melatonin receptors as therapeutic targets in the suprachiasmatic nucleus. Expert Opin. Ther. Targets 2016, 20, 1209–1218. [Google Scholar] [CrossRef] [PubMed]
- Comai, S.; Ochoa-Sanchez, R.; Dominguez-Lopez, S.; Bambico, F.R.; Gobbi, G. Melancholic-Like behaviors and circadian neurobiological abnormalities in melatonin MT1 receptor knockout mice. Int. J. Neuropsychopharmacol. 2015, 18, pyu075. [Google Scholar] [CrossRef]
- Nosjean, O.; Ferro, M.; Coge, F.; Beauverger, P.; Henlin, J.M.; Lefoulon, F.; Fauchere, J.L.; Delagrange, P.; Canet, E.; Boutin, J.A. Identification of the melatonin-binding site MT3 as the quinone reductase 2. J. Biol Chem 2000, 275, 31311–31317. [Google Scholar] [CrossRef]
- Slominski, R.M.; Reiter, R.J.; Schlabritz-Loutsevitch, N.; Ostrom, R.S.; Slominski, A.T. Melatonin membrane receptors in peripheral tissues: Distribution and functions. Mol. Cell Endocrinol. 2012, 351, 152–166. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardeland, R. Melatonin: Signaling mechanisms of a pleiotropic agent. Biofactors 2009, 35, 183–192. [Google Scholar] [CrossRef]
- Cagnacci, A.; Elliott, J.A.; Yen, S.S. Melatonin: A major regulator of the circadian rhythm of core temperature in humans. J. Clin. Endocrinol Metab 1992, 75, 447–452. [Google Scholar]
- Hughes, R.J.; Badia, P. Sleep-promoting and hypothermic effects of daytime melatonin administration in humans. Sleep 1997, 20, 124–131. [Google Scholar] [CrossRef]
- Arendt, J.; Skene, D.J. Melatonin as a chronobiotic. Sleep Med. Rev. 2005, 9, 25–39. [Google Scholar] [CrossRef]
- Cagnacci, A. Melatonin in relation to physiology in adult humans. J. Pineal Res. 1996, 21, 200–213. [Google Scholar] [CrossRef]
- Cagnacci, A.; Soldani, R.; Romagnolo, C.; Yen, S.S. Melatonin-induced decrease of body temperature in women: A threshold event. Neuroendocrinology 1994, 60, 549–552. [Google Scholar] [CrossRef]
- Gilbert, S.S.; van den Heuvel, C.J.; Dawson, D. Daytime melatonin and temazepam in young adult humans: Equivalent effects on sleep latency and body temperatures. J. Physiol. 1999, 514, 905–914. [Google Scholar] [CrossRef]
- Marrin, K.; Drust, B.; Gregson, W.; Atkinson, G. A meta-analytic approach to quantify the dose-response relationship between melatonin and core temperature. Eur. J. Appl. Physiol. 2013, 113, 2323–2329. [Google Scholar] [CrossRef]
- Schwimmer, H.; Mursu, N.; Haim, A. Effects of light and melatonin treatment on body temperature and melatonin secretion daily rhythms in a diurnal rodent, the fat sand rat. Chronobiol. Int. 2010, 27, 1401–1419. [Google Scholar] [CrossRef]
- Sinkalu, V.O.; Ayo, J.O.; Adelaiye, A.B.; Hambolu, J.O. Ameliorative effects of melatonin administration and photoperiods on diurnal fluctuations in cloacal temperature of Marshall broiler chickens during the hot dry season. Int. J. Biometeorol. 2015, 59, 79–87. [Google Scholar] [CrossRef]
- Rivara, S.; Vacondio, F.; Fioni, A.; Silva, C.; Carmi, C.; Mor, M.; Lucini, V.; Pannacci, M.; Caronno, A.; Scaglione, F.; et al. N-(Anilinoethyl)amides: Design and synthesis of metabolically stable, selective melatonin receptor ligands. ChemMedChem 2009, 4, 1746–1755. [Google Scholar] [CrossRef]
- Rivara, S.; Pala, D.; Lodola, A.; Mor, M.; Lucini, V.; Dugnani, S.; Scaglione, F.; Bedini, A.; Lucarini, S.; Tarzia, G.; et al. MT1-selective melatonin receptor ligands: Synthesis, pharmacological evaluation, and molecular dynamics investigation of N-{[(3-O-substituted)anilino]alkyl}amides. ChemMedChem 2012, 7, 1954–1964. [Google Scholar] [CrossRef]
- De Vries, J.; Strubbe, J.H.; Wildering, W.C.; Gorter, J.A.; Prins, A.J. Patterns of body temperature during feeding in rats under varying ambient temperatures. Physiol. Behav. 1993, 53, 229–235. [Google Scholar] [CrossRef]
- Briese, E. Normal body temperature of rats: The setpoint controversy. Neurosci. Biobehav. Rev. 1998, 22, 427–436. [Google Scholar] [CrossRef]
- McElhinny, T.L.; Smale, L.; Holekamp, K.E. Patterns of body temperature, activity, and reproductive behavior in a tropical murid rodent, Arvicanthis niloticus. Physiol. behav. 1997, 62, 91–96. [Google Scholar] [CrossRef]
- Karasek, M. Melatonin, human aging, and age-related diseases. Exp. Gerontol. 2004, 39, 1723–1729. [Google Scholar] [CrossRef]
- Pinato, L.; Ramos, D.; Hataka, A.; Rossignoli, P.S.; Granado, M.D.J.; Mazzetto, M.C.; Campos, L.M.G. Day/night expression of MT1 and MT2 receptors in hypothalamic nuclei of the primate Sapajus apella. J. Chem. Neuroanat. 2017, 81, 10–17. [Google Scholar] [CrossRef]
- Waly, N.E.; Hallworth, R. Circadian Pattern of Melatonin MT1 and MT2 Receptor Localization in the Rat Suprachiasmatic Nucleus. J. Circad. Rhythms 2015, 13, 1. [Google Scholar] [CrossRef]
- Masana, M.I.; Benloucif, S.; Dubocovich, M.L. Circadian rhythm of mt1 melatonin receptor expression in the suprachiasmatic nucleus of the C3H/HeN mouse. J. Pineal Res. 2000, 28, 185–192. [Google Scholar] [CrossRef]
- Odo, M.; Koh, K.; Takada, T.; Yamashita, A.; Narita, M.; Kuzumaki, N.; Ikegami, D.; Sakai, H.; Iseki, M.; Inada, E.; et al. Changes in circadian rhythm for mRNA expression of melatonin 1A and 1B receptors in the hypothalamus under a neuropathic pain-like state. Synapse 2014, 68, 153–158. [Google Scholar] [CrossRef]
- Poirel, V.J.; Masson-Pevet, M.; Pevet, P.; Gauer, F. MT1 melatonin receptor mRNA expression exhibits a circadian variation in the rat suprachiasmatic nuclei. Brain Res. 2002, 946, 64–71. [Google Scholar] [CrossRef]
- Cagnacci, A.; Krauchi, K.; Wirz-Justice, A.; Volpe, A. Homeostatic versus circadian effects of melatonin on core body temperature in humans. J. Biol. Rhythms 1997, 12, 509–517. [Google Scholar] [CrossRef]
- Yeleswaram, K.; McLaughlin, L.G.; Knipe, J.O.; Schabdach, D. Pharmacokinetics and oral bioavailability of exogenous melatonin in preclinical animal models and clinical implications. J. Pineal Res. 1997, 22, 45–51. [Google Scholar] [CrossRef]
- Alam, M.N.; Mallick, B.N. Role of lateral preoptic area alpha-1 and alpha-2 adrenoceptors in sleep-wakefulness and body temperature regulation. Brain Res. Bull. 1994, 35, 171–177. [Google Scholar] [CrossRef]
- Oishi, A.; Cecon, E.; Jockers, R. Melatonin Receptor Signaling: Impact of Receptor Oligomerization on Receptor Function. Int. Rev. Cell Mol. Biol. 2018, 338, 59–77. [Google Scholar] [PubMed]
- Kamal, M.; Gbahou, F.; Guillaume, J.L.; Daulat, A.M.; Benleulmi-Chaachoua, A.; Luka, M.; Chen, P.; Kalbasi Anaraki, D.; Baroncini, M.; Mannoury la Cour, C.; et al. Convergence of melatonin and serotonin (5-HT) signaling at MT2/5-HT2C receptor heteromers. J. Biol. Chem. 2015, 290, 11537–11546. [Google Scholar] [CrossRef]
- Posa, L.; De Gregorio, D.; Gobbi, G.; Comai, S. Targeting Melatonin MT2 Receptors: A Novel Pharmacological Avenue for Inflammatory and Neuropathic Pain. Curr. Med. Chem. 2018, 25, 3866–3882. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Clough, S.J.; Dubocovich, M.L. Role of the MT1 and MT2 melatonin receptors in mediating depressive- and anxiety-like behaviors in C3H/HeN mice. Genes Brain Behav. 2017, 16, 546–553. [Google Scholar] [CrossRef]
- Comai, S.; Lopez-Canul, M.; De Gregorio, D.; Posner, A.; Ettaoussi, M.; Guarnieri, F.; Gobbi, G. Melatonin MT1 receptor as a novel target in neuropsychopharmacology: MT1 ligands, pathophysiological and therapeutic implications and perspectives. Pharmacol. Res. 2019. [Google Scholar] [CrossRef]
- Doolen, S.; Krause, D.N.; Dubocovich, M.L.; Duckles, S.P. Melatonin mediates two distinct responses in vascular smooth muscle. Eur. J. Pharmacol. 1998, 345, 67–69. [Google Scholar] [CrossRef]
- Pandi-Perumal, S.R.; BaHammam, A.S.; Ojike, N.I.; Akinseye, O.A.; Kendzerska, T.; Buttoo, K.; Dhandapany, P.S.; Brown, G.M.; Cardinali, D.P. Melatonin and Human Cardiovascular Disease. J. Cardiovasc. Pharmacol. Ther. 2017, 22, 122–132. [Google Scholar] [CrossRef] [PubMed]
- Comai, S.; Posa, L.; Ochoa-Sanchez, R.; Spadoni, S.; Gobbi, G. Neuropsychopharmacological properties of novel melatonin MT1 receptor ligands. Eur. Neuropsychopharmacol. 2017, 27. [Google Scholar] [CrossRef]
- Pintor, J.; Pelaez, T.; Hoyle, C.H.; Peral, A. Ocular hypotensive effects of melatonin receptor agonists in the rabbit: Further evidence for an MT3 receptor. Br. J. Pharmacol. 2003, 138, 831–836. [Google Scholar] [CrossRef] [PubMed]
- Zurowski, D.; Nowak, L.; Machowska, A.; Wordliczek, J.; Thor, P.J. Exogenous melatonin abolishes mechanical allodynia but not thermal hyperalgesia in neuropathic pain. The role of the opioid system and benzodiazepine-gabaergic mechanism. J. Physiol. Pharmacol. 2012, 63, 641–647. [Google Scholar]
- Molinari, E.J.; North, P.C.; Dubocovich, M.L. 2-[125I]iodo-5-methoxycarbonylamino-N-acetyltryptamine: A selective radioligand for the characterization of melatonin ML2 binding sites. Eur. J. Pharmacol. 1996, 301, 159–168. [Google Scholar] [CrossRef]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
López-Canul, M.; Min, S.H.; Posa, L.; De Gregorio, D.; Bedini, A.; Spadoni, G.; Gobbi, G.; Comai, S. Melatonin MT1 and MT2 Receptors Exhibit Distinct Effects in the Modulation of Body Temperature across the Light/Dark Cycle. Int. J. Mol. Sci. 2019, 20, 2452. https://doi.org/10.3390/ijms20102452
López-Canul M, Min SH, Posa L, De Gregorio D, Bedini A, Spadoni G, Gobbi G, Comai S. Melatonin MT1 and MT2 Receptors Exhibit Distinct Effects in the Modulation of Body Temperature across the Light/Dark Cycle. International Journal of Molecular Sciences. 2019; 20(10):2452. https://doi.org/10.3390/ijms20102452
Chicago/Turabian StyleLópez-Canul, Martha, Seung Hyun Min, Luca Posa, Danilo De Gregorio, Annalida Bedini, Gilberto Spadoni, Gabriella Gobbi, and Stefano Comai. 2019. "Melatonin MT1 and MT2 Receptors Exhibit Distinct Effects in the Modulation of Body Temperature across the Light/Dark Cycle" International Journal of Molecular Sciences 20, no. 10: 2452. https://doi.org/10.3390/ijms20102452