Analysis of the microRNA Expression Profile of Bovine Monocyte-derived Macrophages Infected with Mycobacterium avium subsp. Paratuberculosis Reveals that miR-150 Suppresses Cell Apoptosis by Targeting PDCD4


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
2.1. Mapping and Annotation of miRNA Sequencing Data
2.2. Differentially Expressed miRNAs in MDM Challenged with MAP
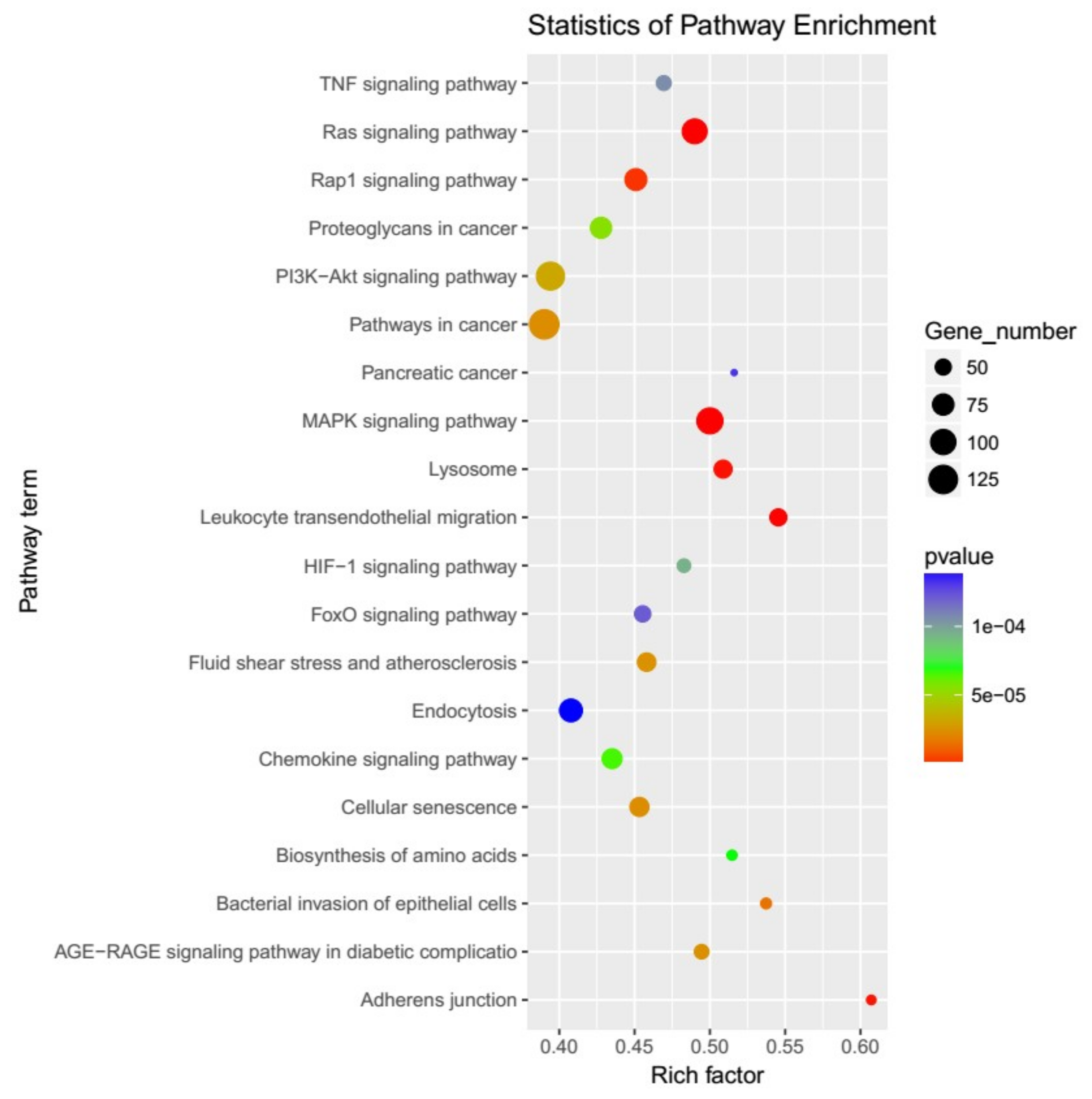
2.3. Prediction and Functional Characterization of Target Genes for differentially expressed miRNAs
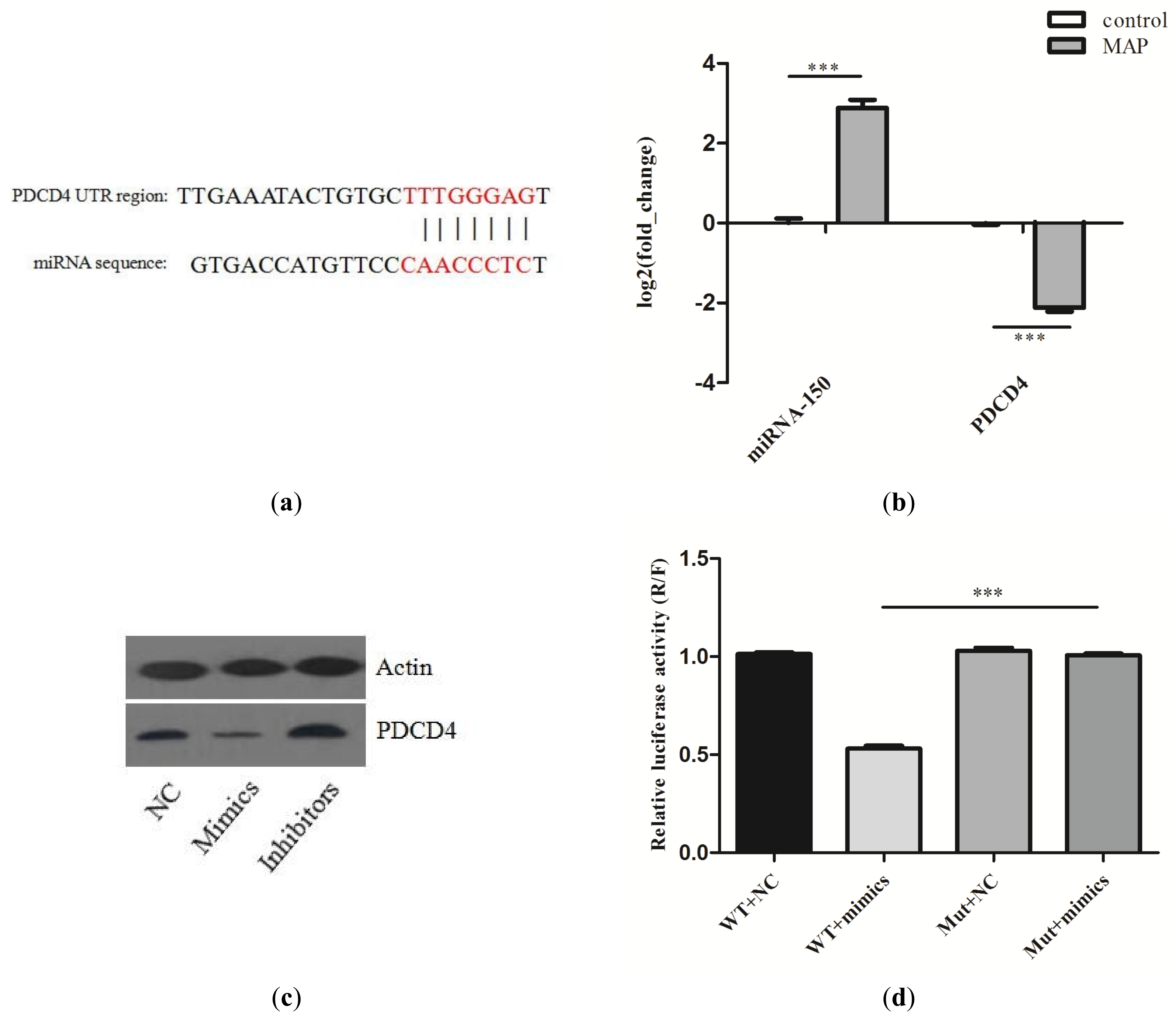
2.4. Validation the Interaction of miR–150 and PDCD4
2.5. miR-150 Suppresses the Apoptosis of Macrophages by Targeting PDCD4
3. Discussion
4. Materials and Methods
4.1. Ethics Statement
4.2. Purification of Bovine Monocyte Derived Macrophages
4.3. Bacterial Infection
4.4. Library Preparation, Sequencing and Data Analysis
4.5. Differential Expression Analysis, Target Prediction and Functional Analysis
4.6. Real-time Quantitative PCR (RT-qPCR)
4.7. Cell Culture
4.8. Vector Construction
4.9. Cell Transfection
4.10. Luciferase Activity Assay
4.11. Cell Apoptosis Assay
4.12. Western Blotting
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| MAP | M. avium subsp. paratuberculosis |
| miRNAs | MicroRNAs |
| MDMs | Monocyte-derived macrophages |
| M. avium | Mycobacterium avium |
| GO | Gene Ontology |
| KEGG | Kyoto Encyclopedia of Genes and Genomes |
| PDCD4 | programmed cell death protein-4 |
References
- Dhama, K.; Mahendran, M.; Tiwari, R.; Dayal, S.S.; Kumar, D.; Singh, S.; Sawant, P.M. Tuberculosis in Birds: Insights into the Mycobacterium avium Infections. Vet. Med. Int. 2011, 2011, 712369. [Google Scholar] [CrossRef] [PubMed]
- Miguez-Burbano, M.J.; Flores, M.; Ashkin, D.; Rodriguez, A.; Granada, A.M.; Quintero, N.; Pitchenik, A. Non-tuberculous mycobacteria disease as a cause of hospitalization in HIV-infected subjects. Int. J. Infect. Dis. 2006, 10, 47–55. [Google Scholar] [Green Version]
- Falkinham, J.O.; Hilborn, E.D.; Arduino, M.J.; Amy, P.; Edwards, M.A. Epidemiology and Ecology of Opportunistic Premise Plumbing Pathogens: Legionella pneumophila, Mycobacterium avium, and Pseudomonas aeruginosa. Environ. Health Perspect. 2015, 123, 749. [Google Scholar]
- Mijs, W.; De, H.P.; Rossau, R.; Van, d.L.T.L.; Portaels, F. Molecular evidence to support a proposal to reserve the designation Mycobacterium avium subsp avium for bird-type isolates and ’M-avium subsp hominissuis’ for the human/porcine type of M-avium. Int. J. Syst. Evolut. Microbiol. 2002, 52, 1505. [Google Scholar]
- Thorel, M.F.; Krichevsky, M.; Lévyfrébault, V.V. Numerical taxonomy of mycobactin-dependent mycobacteria, emended description of Mycobacterium avium, description of Mycobacterium avium subsp. avium subsp. nov., Mycobacterium avium subsp. paratuberculosis subsp. nov., and Mycobacterium avium subsp. silvat. Int. J. Syst. Bacteriol. 1990, 40, 254–260. [Google Scholar] [CrossRef]
- Chiodini, R.J.; Kruiningen, H.J.V.; Merkal, R.S. Ruminant paratuberculosis (Johne’s disease): The current status and future prospects. Corn. Vet. 1984, 74, 218. [Google Scholar] [PubMed]
- Beard, P.M.; Daniels, M.J.; Henderson, D.; Pirie, A.; Rudge, K.; Buxton, D.; Rhind, S.; Greig, A.; Hutchings, M.R.; Mckendrick, I. Paratuberculosis infection of nonruminant wildlife in Scotland. J. Clin. Microbiol. 2001, 39, 1517–1521. [Google Scholar] [CrossRef] [PubMed]
- Cho, J.; Tauer, L.W.; Schukken, Y.H.; Gómez, M.I.; Smith, R.L.; Lu, Z.; Grohn, Y.T. Economic analysis of Mycobacterium avium subspecies paratuberculosis vaccines in dairy herds. J. Dairy Sci. 2012, 95, 1855–1872. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feller, M.; Huwiler, K.; Stephan, R.; Altpeter, E.; Shang, A.; Furrer, H.; Pfyffer, G.E.; Jemmi, T.; Baumgartner, A.; Egger, M. Mycobacterium avium subspecies paratuberculosis and Crohn’s disease: A systematic review and meta-analysis. Lancet Infect. Dis. 2007, 7, 607–613. [Google Scholar] [CrossRef]
- Pierce, E.S. Where are all the Mycobacterium avium subspecies paratuberculosis in patients with Crohn’s disease? PLoS Pathog. 2009, 5, e1000234. [Google Scholar] [CrossRef]
- Arsenault, R.J.; Maattanen, P.; Daigle, J.; Potter, A.; Griebel, P.; Napper, S. From mouth to macrophage: Mechanisms of innate immune subversion by Mycobacterium avium subsp. paratuberculosis. Vet. Res. 2014, 45, 54. [Google Scholar] [CrossRef] [PubMed]
- Kabara, E.; Coussens, P.M. Infection of Primary Bovine Macrophages with Mycobacterium avium Subspecies paratuberculosis Suppresses Host Cell Apoptosis. Front. Microbiol. 2012, 3, 215. [Google Scholar] [CrossRef] [Green Version]
- Behar, S.M.; Maziar, D.; Remold, H.G. Evasion of innate immunity by Mycobacterium tuberculosis: Is death an exit strategy? Nat. Rev. Microbiol. 2010, 8, 668–674. [Google Scholar] [CrossRef]
- Casey, M.E.; Meade, K.G.; Nalpas, N.C.; Taraktsoglou, M.; Browne, J.A.; Killick, K.E.; Park, S.D.; Gormley, E.; Hokamp, K.; Magee, D.A. Analysis of the Bovine Monocyte-Derived Macrophage Response to Mycobacterium avium Subspecies Paratuberculosis Infection Using RNA-seq. Front. Immunol. 2015, 6, 23. [Google Scholar] [CrossRef] [PubMed]
- Machugh, D.E. Pan-genomic analysis of bovine monocyte-derived macrophage gene expression in response to in vitro infection with Mycobacterium avium subspecies paratuberculosis. Vet. Res. 2012, 43, 25. [Google Scholar] [CrossRef] [PubMed]
- Yujing, B.; Guangwei, L.; Ruifu, Y. MicroRNAs: Novel regulators during the immune response. J. Cell. Physiol. 2010, 218, 467–472. [Google Scholar]
- Ivey, K.N.; Srivastava, D. MicroRNAs as Regulators of Differentiation and Cell Fate Decisions. Cell Stem Cell 2010, 7, 36–41. [Google Scholar] [CrossRef] [Green Version]
- Schickel, R.; Boyerinas, B.S.; Peter, M. MicroRNAs: Key players in the immune system, differentiation, tumorigenesis and cell death. Oncogene 2008, 27, 5959–5974. [Google Scholar] [CrossRef]
- Carrington, J.C.; Victor, A. Role of microRNAs in plant and animal development. Science 2003, 301, 336–338. [Google Scholar] [CrossRef]
- O’Connell, R.M.; Rao, D.S.; Chaudhuri, A.A.; David, B. Physiological and pathological roles for microRNAs in the immune system. Nat. Rev. Immunol. 2010, 10, 111–122. [Google Scholar] [CrossRef]
- Inui, M.; Martello, G.S. MicroRNA control of signal transduction. Nat. Rev. Mol. Cell Biol 2010, 11, 252–263. [Google Scholar] [CrossRef] [PubMed]
- Jutta, S.; Astrid, L.; Barbara, K.L.; Elisabeth, K.; Ralf, E.; Soroush, S. Integrated microRNA-mRNA-analysis of human monocyte derived macrophages upon Mycobacterium avium subsp. hominissuis infection. PLoS ONE 2011, 6, e20258. [Google Scholar]
- Casselli, T.; Qureshi, H.; Peterson, E.; Perley, D.; Blake, E.; Jokinen, B.; Abbas, A.; Nechaev, S.; Watt, J.A.; Dhasarathy, A. MicroRNA and mRNA Transcriptome Profiling in Primary Human Astrocytes Infected with Borrelia burgdorferi. PLoS ONE 2017, 12, e0170961. [Google Scholar] [CrossRef] [PubMed]
- Abu-Halima, M.; Kahraman, M.; Henn, D.; Rädle-Hurst, T.; Keller, A.; Abdul-Khaliq, H.; Meese, E. Deregulated microRNA and mRNA expression profiles in the peripheral blood of patients with Marfan syndrome. J. Translat. Med. 2018, 16, 60. [Google Scholar] [CrossRef]
- Verma, P.; Pandey, R.K.; Prajapati, P.; Prajapati, V.K. Circulating MicroRNAs: Potential and Emerging Biomarkers for Diagnosis of Human Infectious Diseases. Front. Microbiol. 2016, 7, 1274. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Mao, L.; Li, W.; Hao, F.; Zhong, C.; Zhu, X.; Ji, X.; Yang, L.; Zhang, W.; Liu, M. Analysis of microRNAs Expression Profiles in Madin-Darby Bovine Kidney Cells Infected With Caprine Parainfluenza Virus Type 3. Front. Cell. Infect. Microbiol. 2018, 8, 93. [Google Scholar] [CrossRef] [Green Version]
- Lewandowska-Sabat, A.M.; Hansen, S.F.; Solberg, T.R.; Østerås, O.; Heringstad, B.; Boysen, P.; Olsaker, I. MicroRNA expression profiles of bovine monocyte-derived macrophages infected in vitro with two strains of Streptococcus agalactiae. BMC Genomics. 2018, 19, 241. [Google Scholar] [CrossRef] [PubMed]
- Lawless, N.; Reinhardt, T.A.; Bryan, K.; Baker, M.; Pesch, B.; Zimmerman, D.; Zuelke, K.; Sonstegard, T.; Lippolis, J.D.; Lynn, D.J. MicroRNA Regulation of Bovine Monocyte Inflammatory and Metabolic Networks in an In Vivo Infection Model. G3 2014, 4, 957. [Google Scholar] [CrossRef]
- Kabara, E.; Kloss, C.C.; Wilson, M.; Tempelman, R.J.; Sreevatsan, S.; Janagama, H.; Coussens, P.M. A large-scale study of differential gene expression in monocyte-derived macrophages infected with several strains of Mycobacterium avium subspecies paratuberculosis. Brief. Funct. Geno. 2010, 9, 220–237. [Google Scholar] [CrossRef]
- Widdison, S.; Watson, M.; Coffey, T.J. Early response of bovine alveolar macrophages to infection with live and heat-killed Mycobacterium bovis. Dev. Compar. Immunol. 2011, 35, 580–591. [Google Scholar]
- Tailleux, L.; Waddell, S.J.; Pelizzola, M.; Mortellaro, A.; Withers, M.; Tanne, A.; Castagnoli, P.R.; Gicquel, B.; Stoker, N.G.; Butcher, P.D. Probing Host Pathogen Cross-Talk by Transcriptional Profiling of Both Mycobacterium tuberculosis and Infected Human Dendritic Cells and Macrophages. PLoS ONE 2008, 3, e1403. [Google Scholar] [CrossRef] [PubMed]
- Bartel, D.P. MicroRNAs: Target Recognition and Regulatory Functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luke A, O.N. MicroRNAs: The fine-tuners of Toll-like receptor signalling. Nat. Rev. Immunol. 2011, 3, 11. [Google Scholar]
- Stenfeldt, C.; Arzt, J.; Smoliga, G.; Larocco, M.; Gutkoska, J.; Lawrence, P. Proof-of-concept study: Profile of circulating microRNAs in Bovine serum harvested during acute and persistent FMDV infection. Virol. J. 2017, 14, 71. [Google Scholar] [CrossRef]
- Rui, L.; Cheng-Long, Z.; Xiang-Xiang, L.; Dan, C.; Wen-Qiang, W.; Yi-Hui, Z.; Xiao-Han, G.; De-Jun, J.; Yong-Jiang, M.; Yun-Chen, G. Transcriptome MicroRNA Profiling of Bovine Mammary Glands Infected with Staphylococcus aureus. Int. J. Mol. Sci. 2015, 16, 4997. [Google Scholar]
- Sun, J.; Aswath, K.; Schroeder, S.G.; Lippolis, J.D.; Reinhardt, T.A.; Sonstegard, T.S. MicroRNA expression profiles of bovine milk exosomes in response to Staphylococcus aureus infection. BMC Genomics. 2015, 16, 806. [Google Scholar] [CrossRef]
- Luoreng, Z.M.; Wang, X.P.; Mei, C.G.; Zan, L.S. Expression profiling of peripheral blood miRNA using RNAseq technology in dairy cows with Escherichia coli-induced mastitis. Sci. Rep. 2018, 8, 12693. [Google Scholar] [CrossRef]
- Dimitrios, L.; Gabriel, P.; Stephen, H.; Fiona, G.; Martin, F.; Milne, R.S.B.; Frances, G.; Chris, B. miR-132 regulates antiviral innate immunity through suppression of the p300 transcriptional co-activator. Nat. Cell Biol. 2010, 12, 513–519. [Google Scholar]
- Fen, L.; Rong, J.; Zhenguo, Z.; Ning, Z.; Liang, X.; Cheng, N.; Kejian, Q. The expression changes in microRNA-132 in the lipopolysaccharide-induced inflammation of rat alveolar macrophages. Zhonghua Wei Zhong Bing Ji Jiu Yi Xue 2014, 26, 80. (In Chinese) [Google Scholar]
- Chen, H.; Untiveros, G.M.; Mckee, L.A.K.; Perez, J.; Li, J.; Antin, P.B.; Konhilas, J.P. Micro-RNA-195 and -451 Regulate the LKB1/AMPK Signaling Axis by Targeting MO25. PLoS ONE 2012, 7, e41574. [Google Scholar] [CrossRef]
- Rosenberger, C.M.; Podyminogin, R.L.; Garnet, N.; Guo-Wei, Z.; Askovich, P.S.; Weiss, M.J.; Alan, A. miR-451 regulates dendritic cell cytokine responses to influenza infection. J. Immunol. 2012, 189, 5965–5975. [Google Scholar] [CrossRef] [PubMed]
- Devram Sampat, G.; Sahana, H.; Kaveri, S.V.; Jagadeesh, B.; Patil, S.A. Sonic hedgehog-dependent induction of microRNA 31 and microRNA 150 regulates Mycobacterium bovis BCG-driven toll-like receptor 2 signaling. Mol. Cell. Biol. 2013, 33, 543–556. [Google Scholar]
- Nobuko, T.; Won, K.H.; Muhammad, A. microRNA-150 Regulates Mobilization and Migration of Bone Marrow-Derived Mononuclear Cells by TargetingCxcr4. PLoS ONE 2011, 6, e23114. [Google Scholar]
- Gong, F.H.; Cheng, W.L.; Wang, H.; Gao, M.; Qin, J.J.; Zhang, Y.; Li, X.; Zhu, X.; Xia, H.; She, Z.G. Reduced atherosclerosis lesion size, inflammatory response in miR-150 knockout mice via macrophage effects. J. Lipid Res. 2018, 59, jlr.M082651. [Google Scholar] [CrossRef] [PubMed]
- Leelahavanichkul, A.; Somparn, P.; Panich, T.; Chancharoenthana, W.; Wongphom, J.; Pisitkun, T.; Hirankarn, N.; Eiam-Ong, S. The levels of serum microRNA-122 in acute liver injury induced by kidney injury and sepsis in CD-1 mouse models. Hepatol. Res. 2015, 45, 1341–1352. [Google Scholar] [CrossRef] [PubMed]
- Over, K.; Crandall, P.G.; O’Bryan, C.A.; Ricke, S.C. Current perspectives on Mycobacterium avium subsp. paratuberculosis, Johne’s disease, and Crohn’s disease: A review. Critical Rev. Microbiol. 2011, 37, 141. [Google Scholar] [CrossRef]
- Malvisi, M.; Palazzo, F.; Morandi, N.; Lazzari, B.; Williams, J.L.; Pagnacco, G.; Minozzi, G. Responses of Bovine Innate Immunity toMycobacterium aviumsubsp.paratuberculosisInfection Revealed by Changes in Gene Expression and Levels of MicroRNA. PLoS ONE 2016, 11, e0164461. [Google Scholar] [CrossRef]
- Farrell, D.; Shaughnessy, R.G.; Britton, L.; Machugh, D.E.; Markey, B.; Gordon, S.V. The Identification of Circulating MiRNA in Bovine Serum and Their Potential as Novel Biomarkers of Early Mycobacterium avium subsp paratuberculosis Infection. PLoS ONE 2015, 10, e0134310. [Google Scholar] [CrossRef]
- Gupta, P.; Peter, S.; Jung, M.; Lewin, A.; Hemmrich-Stanisak, G.; Franke, A. Analysis of long non-coding RNA and mRNA expression in bovine macrophages brings up novel aspects of Mycobacterium avium subspecies paratuberculosis infections. Sci. Rep. 2019, 9, 1571. [Google Scholar] [CrossRef]
- Li, J.Z.; Gao, W.; Ho, W.K.; Lei, W.B.; Wei, W.I.; Chan, J.Y.; Wong, T.S. The clinical association of programmed cell death protein 4 PDCD4 with solid tumors and its prognostic significance: A meta-analysis. Chin. J. Cancer 2016, 35, 683–698. [Google Scholar] [CrossRef]
- Zhike, C.; Yate-Ching, Y.; Yuanzhong, W.; Zheng, L.; Hei Jason, C.; Shiuan, C. Down-regulation of programmed cell death 4 (PDCD4) is associated with aromatase inhibitor resistance and a poor prognosis in estrogen receptor-positive breast cancer. Breast Cancer Res. Treatment 2015, 152, 29–39. [Google Scholar] [Green Version]
- Jia, Z.; Wang, J.; Shi, Q.; Liu, S.; Wang, W.; Tian, Y.; Lu, Q.; Chen, P.; Ma, K.; Zhou, C. SOX6 and PDCD4 enhance cardiomyocyte apoptosis through LPS-induced miR-499 inhibition. Apoptosis 2016, 21, 174–183. [Google Scholar] [CrossRef]
- Lankat-Buttgereit, B.; Göke, R. Programmed cell death protein 4 (pdcd4): A novel target for antineoplastic therapy? Biol. Cell 2012, 95, 515–519. [Google Scholar] [CrossRef]
- Liang, X.; Xu, Z.; Yuan, M.; Zhang, Y.; Zhao, B.; Wang, J.; Zhang, A.; Li, G. MicroRNA-16 suppresses the activation of inflammatory macrophages in atherosclerosis by targeting PDCD4. Int. J. Mol. Med. 2016, 37, 967–975. [Google Scholar] [CrossRef] [Green Version]
- Sheedy, F.J.; Palsson, M.E.; Hennessy, E.C.; O’Leary, J.J.; Ruan, Q.; Johnson, D.S.; Chen, Y.; O’Neill, L.A. Negative regulation of TLR4 via targeting of the proinflammatory tumor suppressor PDCD4 by the microRNA miR-21. Nat. Immunol. 2010, 11, 141. [Google Scholar] [CrossRef] [PubMed]
- Wan, J.; Jie, Y.; Huang, Y.; Deng, L. MicroRNA-150 inhibitors enhance cell apoptosis of melanoma by targeting PDCD4. Oncol. Lett. 2017, 15, 1475–1482. [Google Scholar] [CrossRef] [PubMed]
- Lamont, E.A.; Talaat, A.M.; Coussens, P.M.; Bannantine, J.P.; Grohn, Y.T.; Katani, R.; Li, L.L.; Kapur, V.; Sreevatsan, S. Screening of Mycobacterium avium subsp. paratuberculosis mutants for attenuation in a bovine monocyte-derived macrophage model. Front. Cell Infect. Microbiol. 2014, 4, 87. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Shahid, M.Q.; Wu, J.; Wang, L.; Liu, X.; Lu, Y. Comparative Small RNA Analysis of Pollen Development in Autotetraploid and Diploid Rice. Int. J. Mol. Sci. 2016, 17, 499. [Google Scholar] [CrossRef]





© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, Z.; Kong, L.C.; Jia, B.Y.; Chen, J.R.; Dong, Y.; Jiang, X.Y.; Ma, H.X. Analysis of the microRNA Expression Profile of Bovine Monocyte-derived Macrophages Infected with Mycobacterium avium subsp. Paratuberculosis Reveals that miR-150 Suppresses Cell Apoptosis by Targeting PDCD4. Int. J. Mol. Sci. 2019, 20, 2708. https://doi.org/10.3390/ijms20112708
Wang Z, Kong LC, Jia BY, Chen JR, Dong Y, Jiang XY, Ma HX. Analysis of the microRNA Expression Profile of Bovine Monocyte-derived Macrophages Infected with Mycobacterium avium subsp. Paratuberculosis Reveals that miR-150 Suppresses Cell Apoptosis by Targeting PDCD4. International Journal of Molecular Sciences. 2019; 20(11):2708. https://doi.org/10.3390/ijms20112708
Chicago/Turabian StyleWang, Zi, Ling Cong Kong, Bo Yan Jia, Jing Rui Chen, Yang Dong, Xiu Yun Jiang, and Hong Xia Ma. 2019. "Analysis of the microRNA Expression Profile of Bovine Monocyte-derived Macrophages Infected with Mycobacterium avium subsp. Paratuberculosis Reveals that miR-150 Suppresses Cell Apoptosis by Targeting PDCD4" International Journal of Molecular Sciences 20, no. 11: 2708. https://doi.org/10.3390/ijms20112708