Chondrogenic Differentiation of Pluripotent Stem Cells under Controllable Serum-Free Conditions

 , , ,
, , , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Results
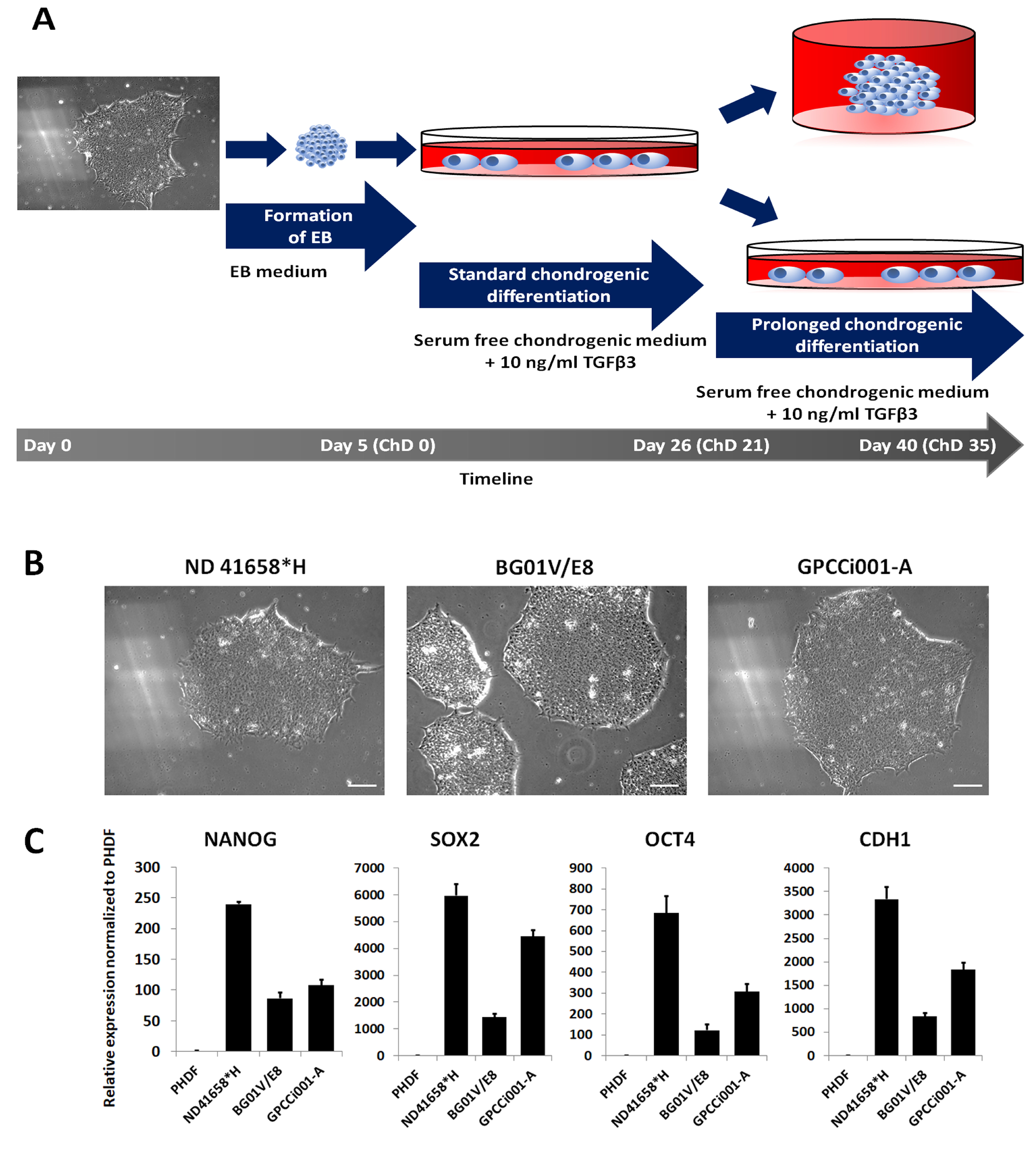
2.1. The Adaptation of Pluripotent Cell Lines to Serum- and Feeder-Free Conditions
2.2. RT-qPCR Analysis of Differentiated Cells Revealed Early Chondrogenesis
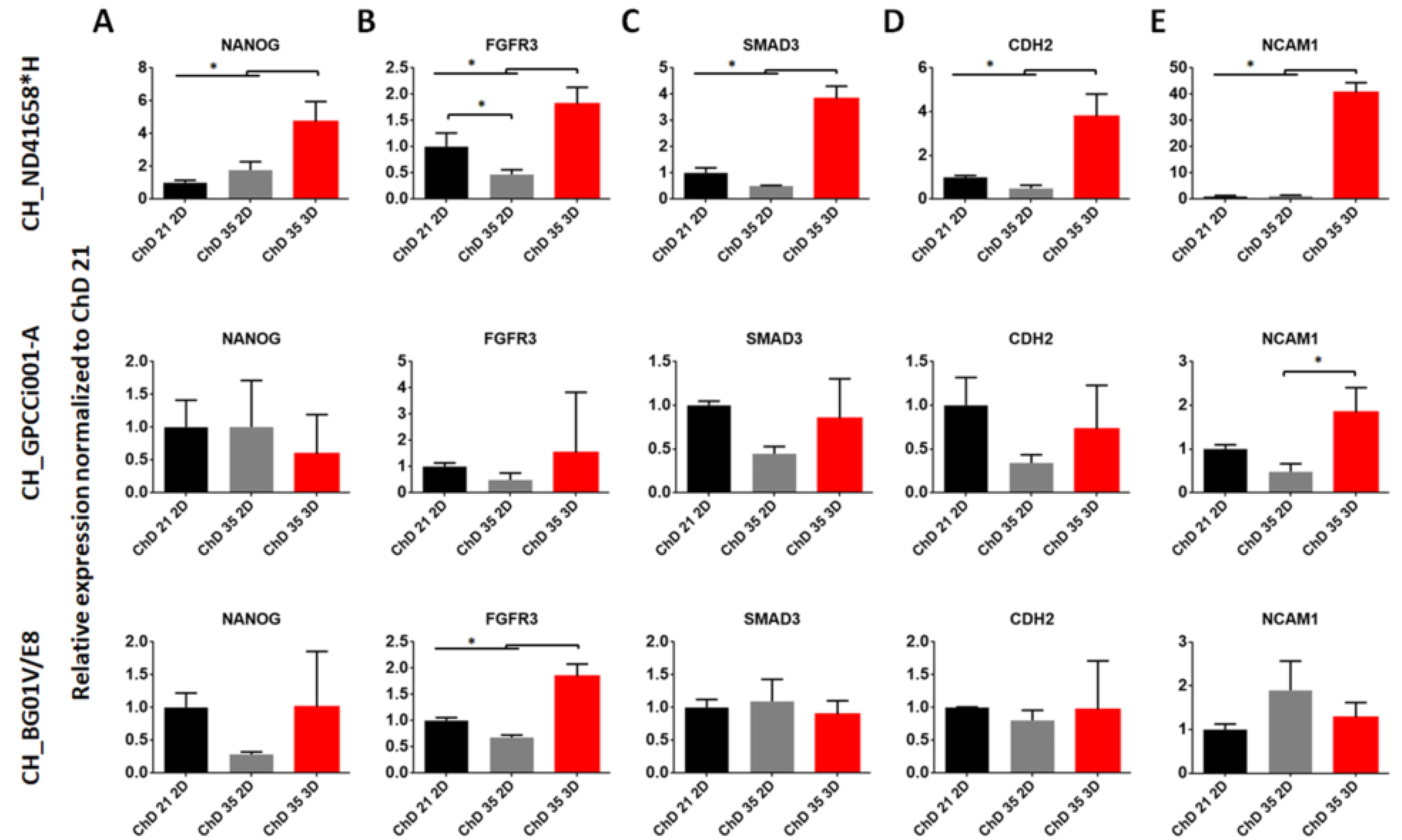
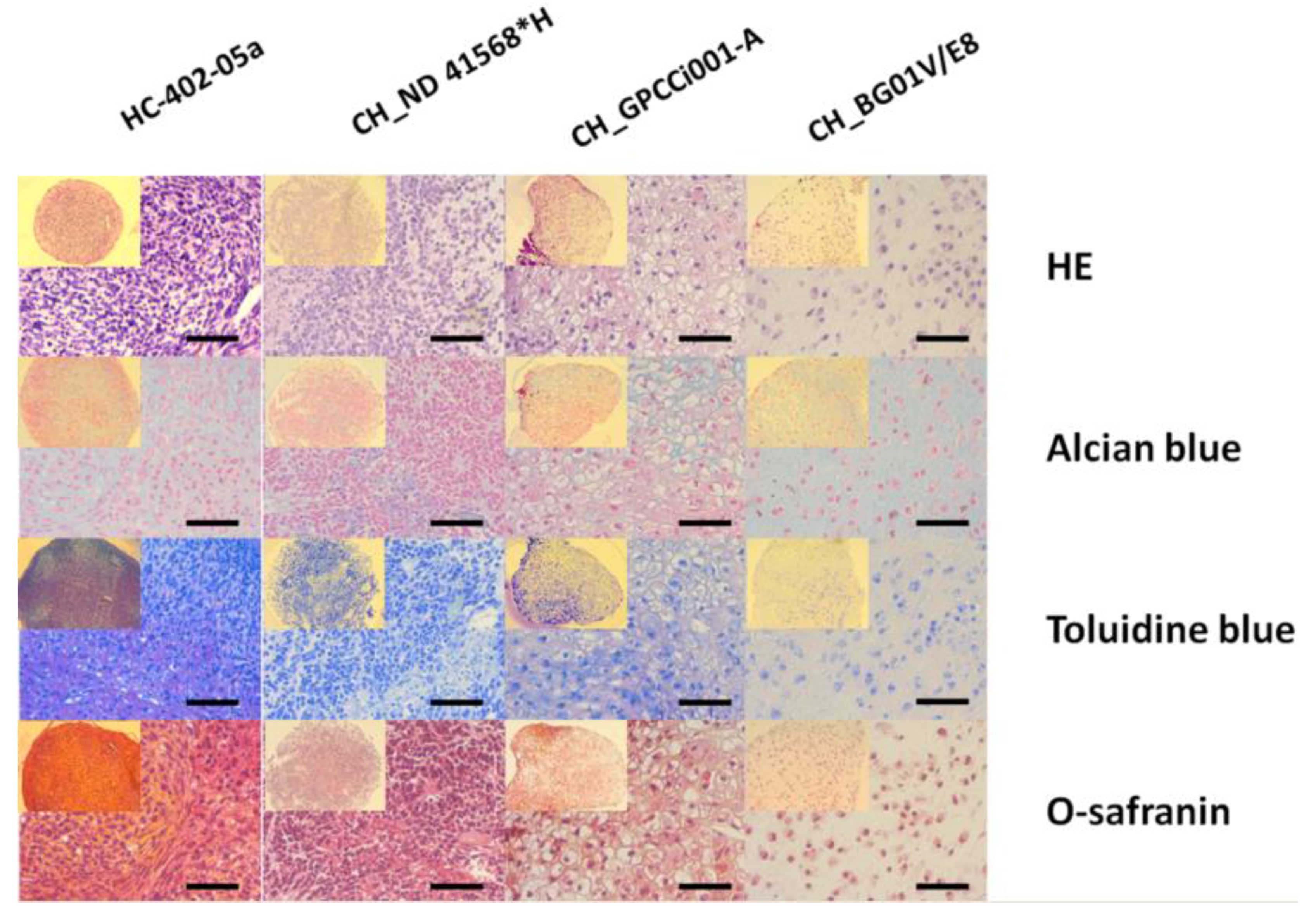
2.3. Prolonged Differentiation Increases the Deposition of ECM Specific for Chondrocytes and 3D Culture Facilitates the Formation of Chondrogenic-Like Spheres
3. Discussion
4. Materials and Methods
4.1. Cell Culture PSC
4.2. Chondrogenic Differentiation
4.3. RT-qPCR
4.4. Histological Staining
4.5. Immunofluorescent Staining of EB Outgrowth
4.6. Determination of Proteoglycans Deposition in 2D Culture
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Abbreviations
| OA | Osteoarthritis |
| iPSCs | Induced Pluripotent Stem Cells |
| 3D | Three-dimensional |
| ECM | Extracellular Matrix |
| ACI | Autologous Chondrocyte Implantation |
| MSC | Mesenchymal Stromal Cells |
| PSC | Pluripotent Stem Cells |
| hESC | Human Embryonic Stem Cells |
| FBS | Fetal Bovine Serum |
| SOX2 | SRY (sex determining region Y)-box 2 |
| OCT4 | Octamer-binding transcription factor 4 |
| CDH1 | Cadherin-1 (E-cadherin) |
| TGF-β3 | Transforming Growth Factor beta 3 |
| EB | Embryoid Body |
| ChD | Chondrogenic Differentiation |
| ChM | Chondrogenic Medium |
| PHDF | Primary Human Dermal Fibroblast |
| RT-qPCR | Reverse Transcriptase quantitative Polymerase Chain Reaction |
| FGFR3 | Fibroblast Growth Factor Receptor 3 |
| SMAD3 | SMAD Family Member 3 |
| CDH2 | Cadherin-2 (N-cadherin) |
| NCAM1 | Neural Cell Adhesion Molecule 1 |
| COL1A2 | Collagen type 1 alpha 2 chain |
| SOX9 | SRY (sex determining region Y)-box 9 |
| COL2A1 | Collagen type 2 alpha 1 chain |
| ACAN | Aggrecan |
| GAGs | Glycosaminoglycans |
| KSR | Knock-out Serum Replacement |
| SCID | Severe Combined Immunodeficiency |
| DMEM | Dulbecco Modified Eagle Medium |
| PBS | Phosphate Buffered Saline |
| EDTA | Ethylenediaminetetraacetic acid |
| NEAA | Non-essential Amino Acids |
| B2M | Beta-2-Microglobulin |
| DAPI | 4′,6-diamidino-2-phenylindole |
References
- Lee, J.K.; Responte, D.J.; Cissell, D.D.; Hu, J.C.; Nolta, J.A.; Athanasiou, K.A. Clinical translation of stem cells: Insight for cartilage therapies. Crit. Rev. Biotechnol. 2014, 8551, 89–100. [Google Scholar] [CrossRef] [PubMed]
- Lach, M.; Trzeciak, T.; Richter, M.; Pawlicz, J.; Suchorska, W.M. Directed differentiation of induced pluripotent stem cells into chondrogenic lineages for articular cartilage treatment. J. Tissue Eng. 2014, 5, 204173141455270. [Google Scholar] [CrossRef] [PubMed]
- Trzeciak, T.; Richter, M.; Suchorska, W. Application of cell and biomaterial-based tissue engineering methods in the treatment of cartilage, menisci and ligament injuries. Int. Orthop. 2016, 40, 615–624. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Augustyniak, E.; Trzeciak, T.; Richter, M.; Kaczmarczyk, J.; Suchorska, W. The role of growth factors in stem cell-directed chondrogenesis: A real hope for damaged cartilage regeneration. Int. Orthop. 2015, 39, 995–1003. [Google Scholar] [CrossRef] [PubMed]
- Cole, B.J.; Pascual-Garrido, C.; Grumet, R.C. Surgical management of articular cartilage defects in the knee. Instr. Course Lect. 2010, 59, 181–204. [Google Scholar]
- Minas, T.; Gomoll, A.H.; Rosenberger, R.; Royce, R.O.; Bryant, T. Increased failure rate of autologous chondrocyte implantation after previous treatment with marrow stimulation techniques. Am. J. Sports Med. 2009, 37, 902–908. [Google Scholar] [CrossRef] [PubMed]
- Erggelet, C. Enhanced Marrow Stimulation Techniques. Oper. Tech. Orthop. 2014, 24, 2–13. [Google Scholar] [CrossRef]
- Li, X.; Iyer, S.; Cross, M.B.; Figgie, M.P. Total joint replacement in adolescents: Literature review and case examples. Curr. Opin. Pediatr. 2012, 24, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Franklin, P.D.; Lewallen, D.; Bozic, K.; Hallstrom, B.; Jiranek, W.; Ayers, D.C. Implementation of Patient-Reported Outcome Measures in U.S. Total Joint Replacement Registries: Rationale, Status, and Plans. J. Bone Jt. Surg.-Am. Vol. 2014, 96, 104–109. [Google Scholar] [CrossRef] [Green Version]
- Mohamed-Ahmed, S.; Fristad, I.; Lie, S.A.; Suliman, S.; Mustafa, K.; Vindenes, H.; Idris, S.B. Adipose-derived and bone marrow mesenchymal stem cells: A donor-matched comparison. Stem Cell Res. Ther. 2018, 9, 168. [Google Scholar] [CrossRef]
- Farrell, M.J.; Fisher, M.B.; Huang, A.H.; Shin, J.I.; Farrell, K.M.; Mauck, R.L. Functional properties of bone marrow-derived MSC-based engineered cartilage are unstable with very long-term in vitro culture. J. Biomech. 2014, 47, 2173–2182. [Google Scholar] [CrossRef] [PubMed]
- Kanawa, M.; Igarashi, A.; Ronald, V.S.; Higashi, Y.; Kurihara, H.; Sugiyama, M.; Saskianti, T.; Pan, H.; Kato, Y. Age-dependent decrease in the chondrogenic potential of human bone marrow mesenchymal stromal cells expanded with fibroblast growth factor-2. Cytotherapy 2013, 15, 1062–1072. [Google Scholar] [CrossRef] [PubMed]
- Cyganek, L.; Tiburcy, M.; Sekeres, K.; Gerstenberg, K.; Bohnenberger, H.; Lenz, C.; Henze, S.; Stauske, M.; Salinas, G.; Zimmermann, W.-H.; et al. Deep phenotyping of human induced pluripotent stem cell–derived atrial and ventricular cardiomyocytes. JCI Insight 2018, 3, 1–17. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Li, W.; Fu, X.; Xu, Y. The Immunogenicity and Immune Tolerance of Pluripotent Stem Cell Derivatives. Front. Immunol. 2017, 8, 6–11. [Google Scholar] [CrossRef] [PubMed]
- Diederichs, S.; Shine, K.M.; Tuan, R.S. The promise and challenges of stem cell-based therapies for skeletal diseases: Stem cell applications in skeletal medicine: Potential, cell sources and characteristics, and challenges of clinical translation. Bioessays 2013, 35, 220–230. [Google Scholar] [CrossRef]
- Narsinh, K.H.; Plews, J.; Wu, J.C.; Wu, C.J.C. Comparison of Human Induced Pluripotent and Embryonic Stem Cells: Fraternal or Identical Twins? Mol. Ther. 2009, 19, 635–638. [Google Scholar] [CrossRef]
- Bilic, J.; Izpisua Belmonte, J.C. Concise review: Induced pluripotent stem cells versus embryonic stem cells: Close enough or yet too far apart? Stem Cells 2012, 30, 33–41. [Google Scholar] [CrossRef]
- Puri, M.C.; Nagy, A. Concise review: Embryonic stem cells versus induced pluripotent stem cells: The game is on. Stem Cells 2012, 30, 10–14. [Google Scholar] [CrossRef]
- Chung, H.C.; Lin, R.C.; Logan, G.J.; Alexander, I.E.; Sachdev, P.S.; Sidhu, K.S. Human induced pluripotent stem cells derived under feeder-free conditions display unique cell cycle and DNA replication gene profiles. Stem Cells Dev. 2012, 21, 206–216. [Google Scholar] [CrossRef]
- Robinton, D.A.; Daley, G.Q. The promise of induced pluripotent stem cells in research and therapy. Nature 2012, 481, 295–305. [Google Scholar] [CrossRef]
- Oldershaw, R.A.; Baxter, M.A.; Lowe, E.T.; Bates, N.; Grady, L.M.; Soncin, F.; Brison, D.R.; Hardingham, T.E.; Kimber, S.J. Directed differentiation of human embryonic stem cells toward chondrocytes. Nat. Biotechnol. 2010, 28, 1187–1194. [Google Scholar] [CrossRef] [PubMed]
- Yamashita, A.; Morioka, M.; Kishi, H.; Kimura, T.; Yahara, Y.; Okada, M.; Fujita, K.; Sawai, H.; Ikegawa, S.; Tsumaki, N. Statin treatment rescues FGFR3 skeletal dysplasia phenotypes. Nature 2014, 513, 507–511. [Google Scholar] [CrossRef] [PubMed]
- Kanke, K.; Masaki, H.; Saito, T.; Komiyama, Y.; Hojo, H.; Nakauchi, H.; Lichtler, A.C.; Takato, T.; Chung, U.I.; Ohba, S. Stepwise differentiation of pluripotent stem cells into osteoblasts using four small molecules under serum-free and feeder-free conditions. Stem Cell Rep. 2014, 2, 751–760. [Google Scholar] [CrossRef] [PubMed]
- Nejadnik, H.; Diecke, S.; Lenkov, O.D.; Chapelin, F.; Donig, J.; Tong, X.; Derugin, N.; Chan, R.C.F.F.; Gaur, A.; Yang, F.; et al. Improved Approach for Chondrogenic Differentiation of Human Induced Pluripotent Stem Cells. Stem Cell Rev. Rep. 2015, 11, 242–253. [Google Scholar] [CrossRef] [Green Version]
- Van der Valk, J.; Mellor, D.; Brands, R.; Fischer, R.; Gruber, F.; Gstraunthaler, G.; Hellebrekers, L.; Hyllner, J.; Jonker, F.H.; Prieto, P.; et al. The humane collection of fetal bovine serum and possibilities for serum-free cell and tissue culture. Toxicol. In Vitro 2004, 18, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Gstraunthaler, G.; Lindl, T.; van der Valk, J. A plea to reduce or replace fetal bovine serum in cell culture media. Cytotechnology 2013, 65, 791–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jochems, C.E.A.; Van der Valk, J.B.F.; Stafleu, F.R.; Baumans, V. The use of fetal bovine serum: Ethical or scientific problem? Altern. Lab. Anim. 2002, 30, 219–227. [Google Scholar] [CrossRef]
- Lach, M.S.; Wroblewska, J.P.; Augustyniak, E.; Kulcenty, K.; Suchorska, W.M. A feeder- and xeno-free human induced pluripotent stem cell line obtained from primary human dermal fibroblasts with epigenetic repression of reprogramming factors expression: GPCCi001-A. Stem Cell Res. 2017, 20, 34–37. [Google Scholar] [CrossRef]
- Lach, M.S.; Kulcenty, K.; Jankowska, K.; Trzeciak, T.; Richter, M.; Suchorska, W.M. Effect of cellular mass on chondrogenic differentiation during embryoid body formation. Mol. Med. Rep. 2018, 18, 2705–2714. [Google Scholar] [CrossRef] [Green Version]
- Yang, S.L.; Harnish, E.; Leeuw, T.; Dietz, U.; Batchelder, E.; Wright, P.S.; Peppard, J.; August, P.; Volle-Challier, C.; Bono, F.; et al. Compound screening platform using human induced pluripotent stem cells to identify small molecules that promote chondrogenesis. Protein Cell 2012, 3, 934–942. [Google Scholar] [CrossRef] [Green Version]
- Van Winkle, A.P.; Gates, I.D.; Kallos, M.S.; Paper, O. Mass Transfer Limitations in Embryoid Bodies during Human Embryonic Stem Cell Differentiation. Cells Tissues Organs 2012, 196, 34–47. [Google Scholar] [CrossRef]
- Sun, N.; Panetta, N.J.; Gupta, D.M.; Wilson, K.D.; Lee, A.; Jia, F.; Hu, S.; Cherry, A.M.; Robbins, R.C.; Longaker, M.T.; et al. Feeder-free derivation of induced pluripotent stem cells from adult human adipose stem cells. Proc. Natl. Acad. Sci. USA 2009, 106, 15720–15725. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Endo, K.; Fujita, N.; Nakagawa, T.; Nishimura, R. Effect of fibroblast growth factor-2 and serum on canine mesenchymal stem cell chondrogenesis. Tissue Eng. Part A 2018. [Google Scholar] [CrossRef] [PubMed]
- Bilgen, B.; Orsini, E.; Aaron, R.K.; Ciombor, D.M. FBS suppresses TGF-β1-induced chondrogenesis in synoviocyte pellet cultures while dexamethasone and dynamic stimuli are beneficial. J. Tissue Eng. Regen. Med. 2007, 1, 436–442. [Google Scholar] [CrossRef] [PubMed]
- Ho, S.T.; Tanavde, V.M.; Hui, J.H.; Lee, E.H. Upregulation of Adipogenesis and Chondrogenesis in MSC Serum-Free Culture. Cell Med. 2011, 2, 27–41. [Google Scholar] [CrossRef] [PubMed]
- Cho, H.; Lee, A.; Kim, K. The effect of serum types on Chondrogenic differentiation of adipose-derived stem cells. Biomater. Res. 2018, 22, 6. [Google Scholar] [CrossRef]
- Yang, H.N.; Park, J.S.; Woo, D.G.; Jeon, S.Y.; Do, H.-J.; Lim, H.-Y.; Kim, S.W.; Kim, J.-H.; Park, K.-H. Chondrogenesis of mesenchymal stem cells and dedifferentiated chondrocytes by transfection with SOX Trio genes. Biomaterials 2011, 32, 7695–7704. [Google Scholar] [CrossRef]
- Goldring, M.B. Chondrogenesis, chondrocyte differentiation, and articular cartilage metabolism in health and osteoarthritis. Ther. Adv. Musculoskelet. Dis. 2012, 4, 269–285. [Google Scholar] [CrossRef]
- Greene, G.W.; Zappone, B.; Söderman, O.; Topgaard, D.; Rata, G.; Zeng, H.; Israelachvili, J.N. Anisotropic dynamic changes in the pore network structure, fluid diffusion and fluid flow in articular cartilage under compression. Biomaterials 2010, 31, 3117–3128. [Google Scholar] [CrossRef]
- Craft, A.M.; Rockel, J.S.; Nartiss, Y.; Kandel, R.A.; Alman, B.A.; Keller, G.M. Generation of articular chondrocytes from human pluripotent stem cells. Nat. Biotechnol. 2015, 33, 638–645. [Google Scholar] [CrossRef]
- Mahboudi, H.; Soleimani, M.; Enderami, S.E.; Kehtari, M.; Ardeshirylajimi, A.; Eftekhary, M.; Kazemi, B. Enhanced chondrogenesis differentiation of human induced pluripotent stem cells by MicroRNA-140 and transforming growth factor beta 3 (TGFβ3). Biologicals 2018, 52, 30–36. [Google Scholar] [CrossRef] [PubMed]
- Kimura, T.; Ozaki, T.; Fujita, K.; Yamashita, A.; Morioka, M.; Ozono, K.; Tsumaki, N. Proposal of patient-specific growth plate cartilage xenograft model for FGFR3 chondrodysplasia. Osteoarthr. Cartil. 2018, 26, 1551–1561. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Chen, L.; Xu, X.; Li, C.; Huang, C.; Deng, C.X. TGF-β/Smad3 signals repress chondrocyte hypertrophic differentiation and are required for maintaining articular cartilage. J. Cell Biol. 2001, 153, 35–46. [Google Scholar] [CrossRef]
- De Kroon, L.M.G.; Narcisi, R.; Van Den Akker, G.G.H.; Vitters, E.L.; Blaney Davidson, E.N.; Van Osch, G.J.V.M.; Van Der Kraan, P.M. SMAD3 and SMAD4 have a more dominant role than SMAD2 in TGFβ-induced chondrogenic differentiation of bone marrow-derived mesenchymal stem cells. Sci. Rep. 2017, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Zhu, Y.; Tao, H.; Jin, C.; Liu, Y.; Lu, X.; Hu, X.; Wang, X. Transforming growth factor-β1 induces type II collagen and aggrecan expression via activation of extracellular signal-regulated kinase 1/2 and Smad2/3 signaling pathways. Mol. Med. Rep. 2015, 12, 5573–5579. [Google Scholar] [CrossRef] [PubMed]
- Chimal-Monroy, J.; De León, L.D. Expression of N-cadherin, N-CAM, fibronectin and tenascin is stimulated by TGF-β1, β2, β3 and β5 during the formation of precartilage condensations. Int. J. Dev. Biol. 1999, 43, 59–67. [Google Scholar]
- Tavella, S.; Raffo, P.; Tacchetti, C.; Cancedda, R.; Castagnola, P. N-CAM and N-Cadherin Expression during in Vitro Chondrogenesis. Exp. Cell Res. 1994, 215, 354–362. [Google Scholar] [CrossRef]
- Demoor, M.; Ollitrault, D.; Gomez-Leduc, T.; Bouyoucef, M.; Hervieu, M.; Fabre, H.; Lafont, J.; Denoix, J.-M.; Audigié, F.; Mallein-Gerin, F.; et al. Cartilage tissue engineering: Molecular control of chondrocyte differentiation for proper cartilage matrix reconstruction. Biochim. Biophys. Acta 2014, 1840, 2414–2440. [Google Scholar] [CrossRef]
- Yang, D.; Chen, S.; Gao, C.; Liu, X.; Zhou, Y.; Liu, P.; Cai, J. Chemically defined serum-free conditions for cartilage regeneration from human embryonic stem cells. Life Sci. 2016, 164, 9–14. [Google Scholar] [CrossRef]
- Yamashita, A.; Morioka, M.; Yahara, Y.; Okada, M.; Kobayashi, T.; Kuriyama, S.; Matsuda, S.; Tsumaki, N. Generation of scaffoldless hyaline cartilaginous tissue from human iPSCs. Stem Cell Rep. 2015, 4, 404–418. [Google Scholar] [CrossRef]
- Nam, Y.; Rim, Y.A.; Jung, S.M.; Ju, J.H. Cord blood cell-derived iPSCs as a new candidate for chondrogenic differentiation and cartilage regeneration. Stem Cell Res. Ther. 2017, 8, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rim, Y.A.; Nam, Y.; Ju, J.H. Application of Cord Blood and Cord Blood-derived Induced Pluripotent Stem Cells for Cartilage Regeneration. Cell Transplant. 2018, 096368971879486. [Google Scholar] [CrossRef] [PubMed]
- Suchorska, W.M.; Lach, M.S.; Richter, M.; Kaczmarczyk, J.; Trzeciak, T. Bioimaging: An Useful Tool to Monitor Differentiation of Human Embryonic Stem Cells into Chondrocytes. Ann. Biomed. Eng. 2016, 44, 1845–1859. [Google Scholar] [CrossRef] [PubMed]






© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lach, M.S.; Wroblewska, J.; Kulcenty, K.; Richter, M.; Trzeciak, T.; Suchorska, W.M. Chondrogenic Differentiation of Pluripotent Stem Cells under Controllable Serum-Free Conditions. Int. J. Mol. Sci. 2019, 20, 2711. https://doi.org/10.3390/ijms20112711
Lach MS, Wroblewska J, Kulcenty K, Richter M, Trzeciak T, Suchorska WM. Chondrogenic Differentiation of Pluripotent Stem Cells under Controllable Serum-Free Conditions. International Journal of Molecular Sciences. 2019; 20(11):2711. https://doi.org/10.3390/ijms20112711
Chicago/Turabian StyleLach, Michał Stefan, Joanna Wroblewska, Katarzyna Kulcenty, Magdalena Richter, Tomasz Trzeciak, and Wiktoria Maria Suchorska. 2019. "Chondrogenic Differentiation of Pluripotent Stem Cells under Controllable Serum-Free Conditions" International Journal of Molecular Sciences 20, no. 11: 2711. https://doi.org/10.3390/ijms20112711