The Respiratory Burst Oxidase Homolog D (RbohD) Cell and Tissue Distribution in Potato–Potato Virus Y (PVYNTN) Hypersensitive and Susceptible Reactions



Abstract
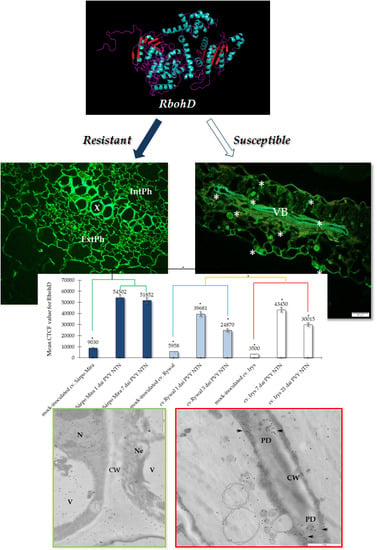
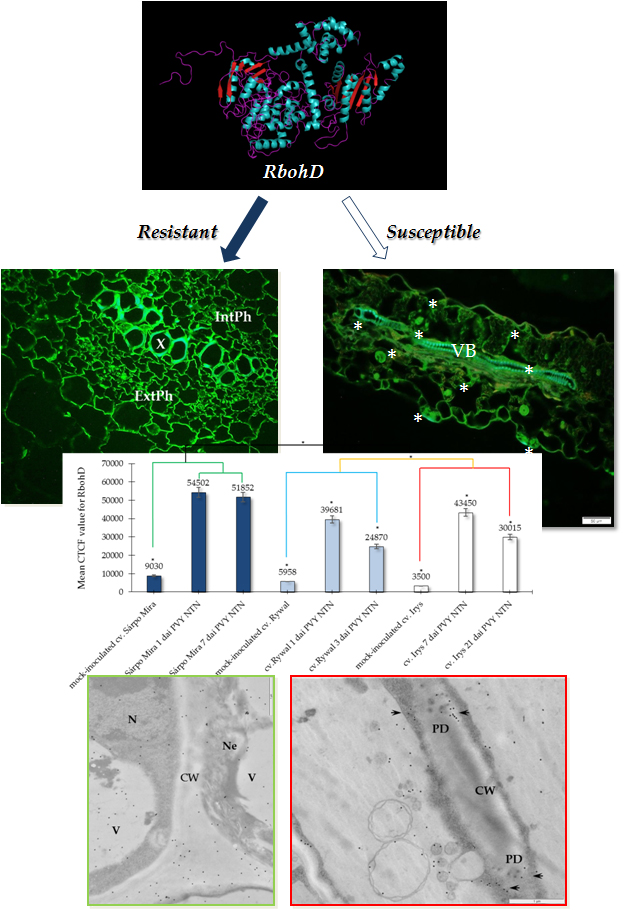
:
1. Introduction
2. Results
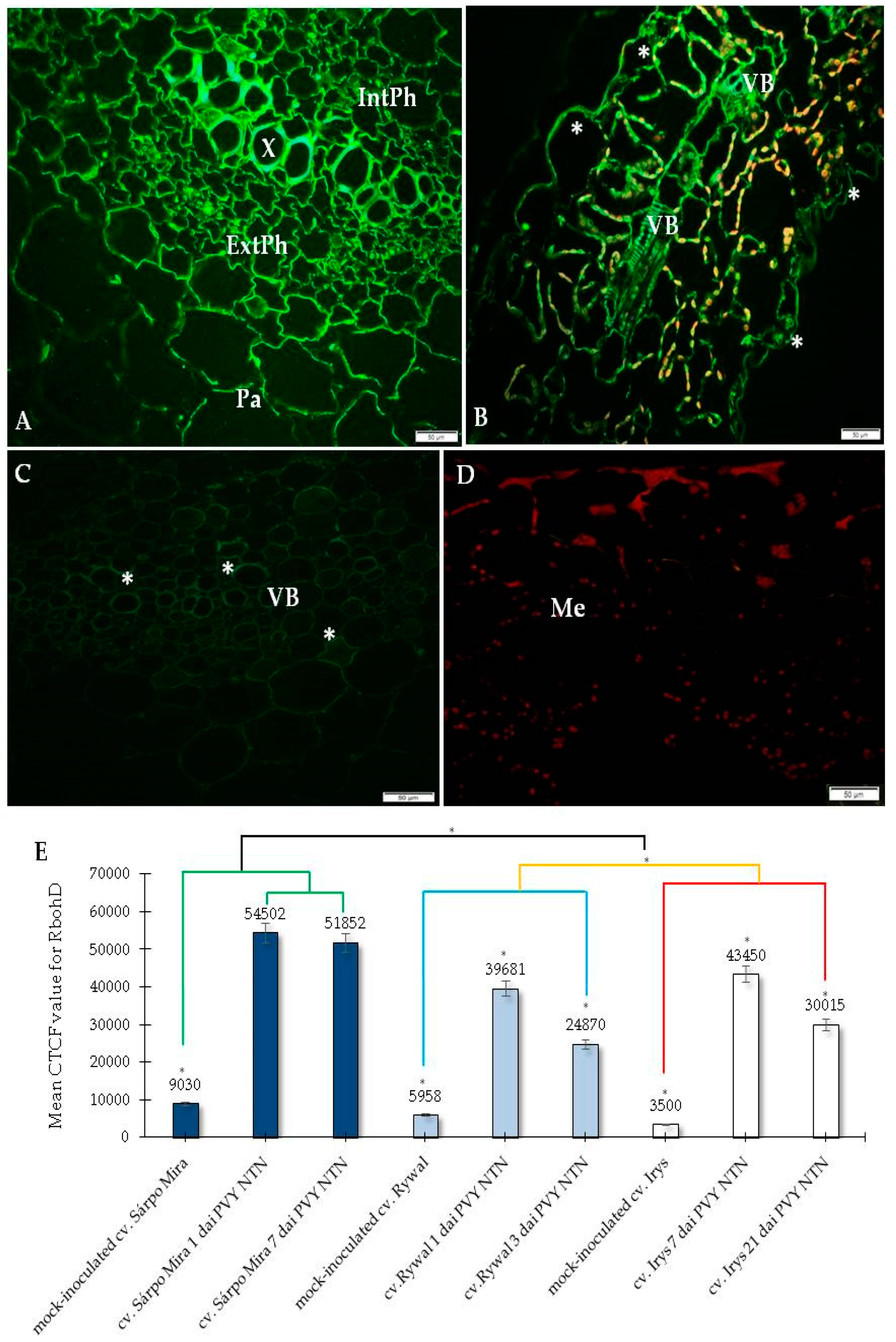
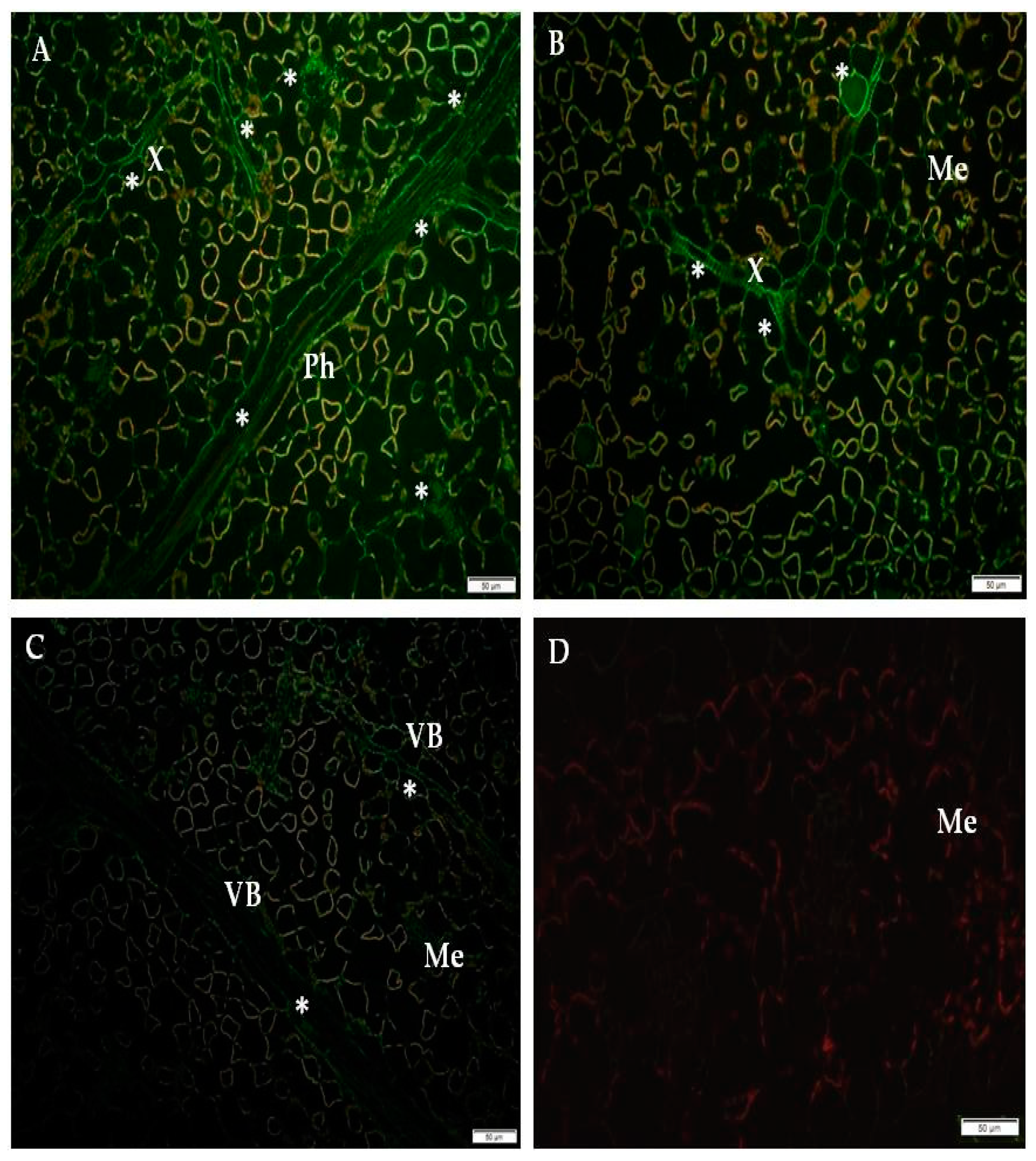
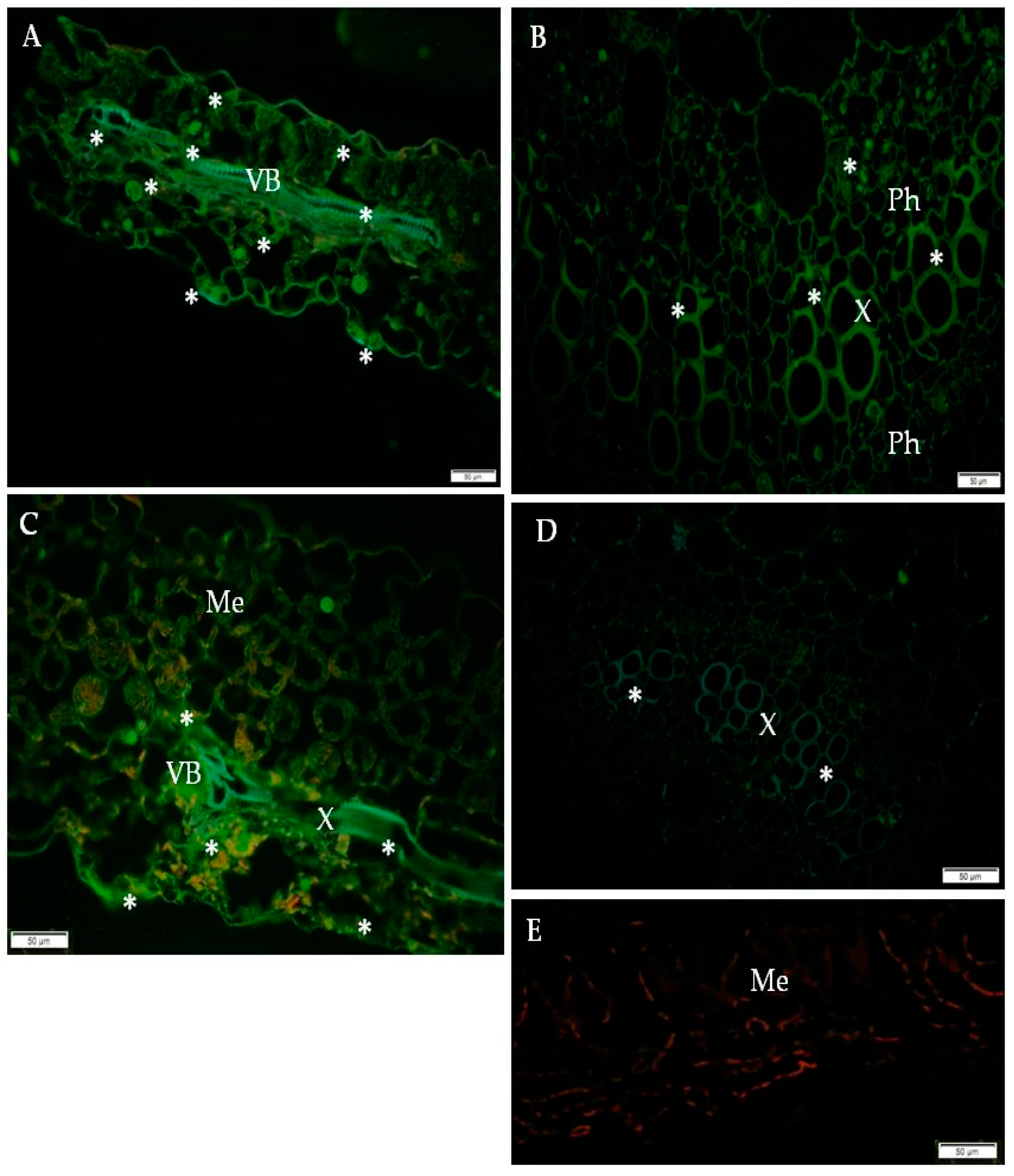
2.1. Immunofluorescence Localization of RbohD Protein in Compatible and Incompatible Interactions
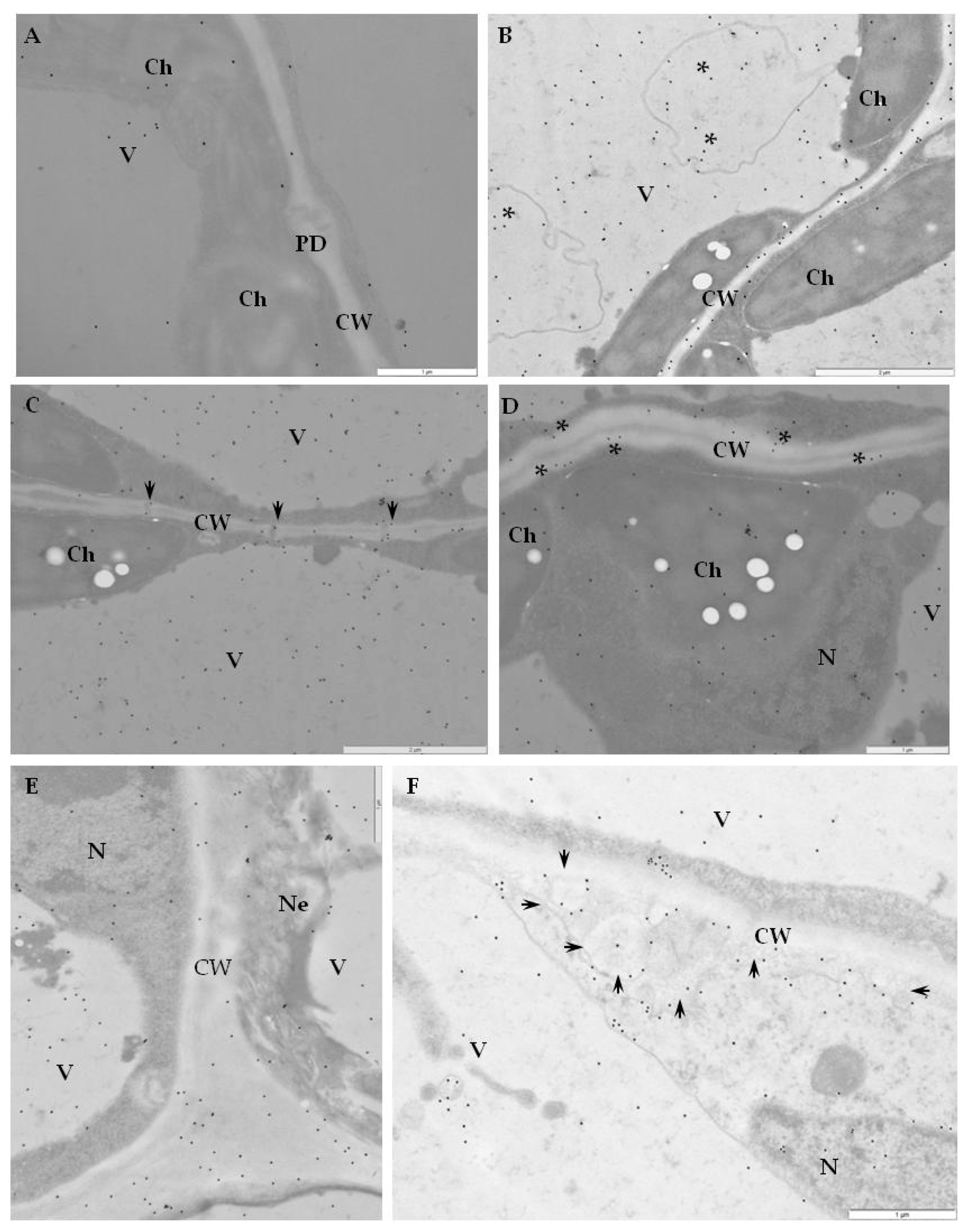
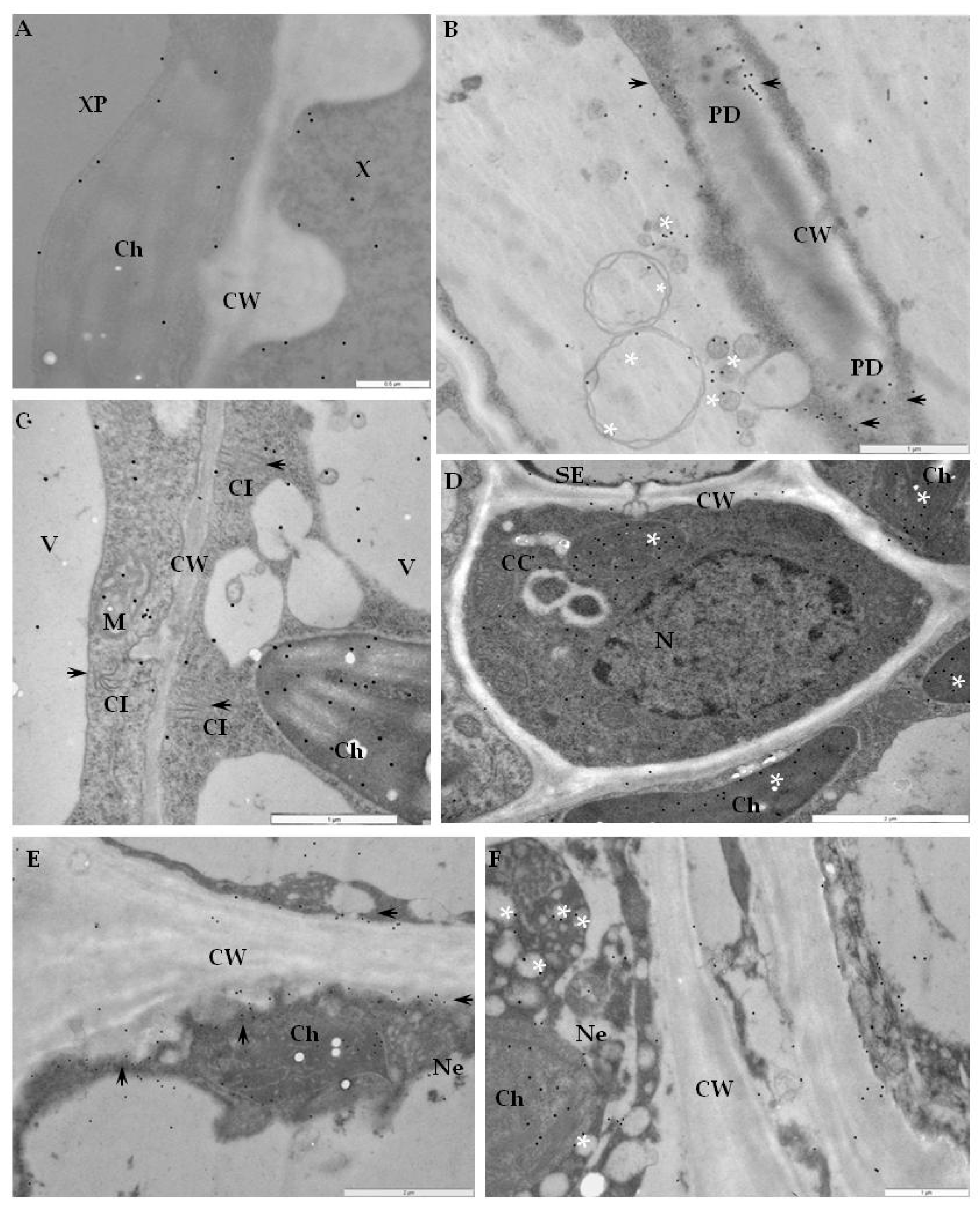
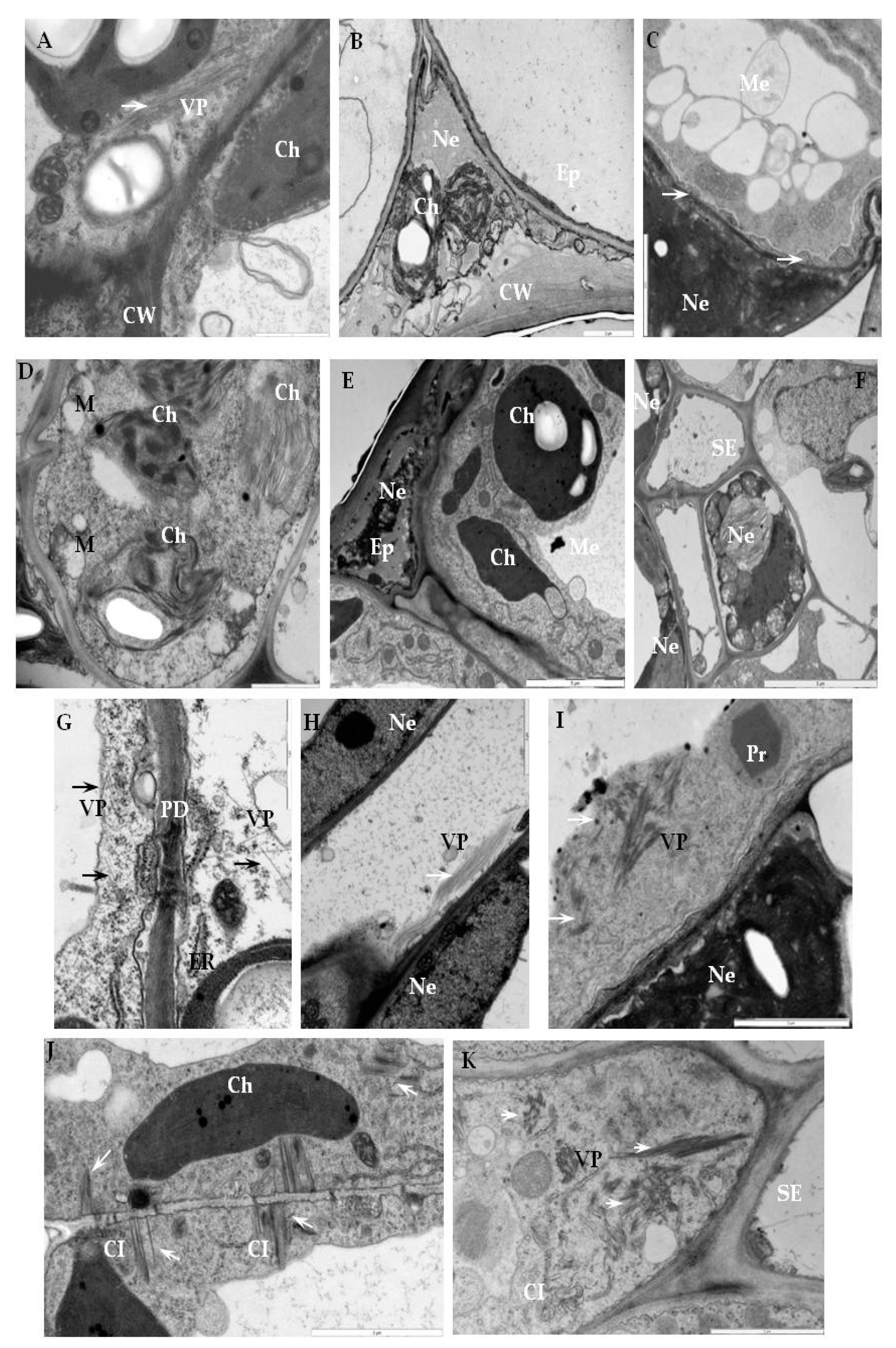
2.2. Ultrastructural Localization of RbohD Protein in Compatible and Incompatible Interactions
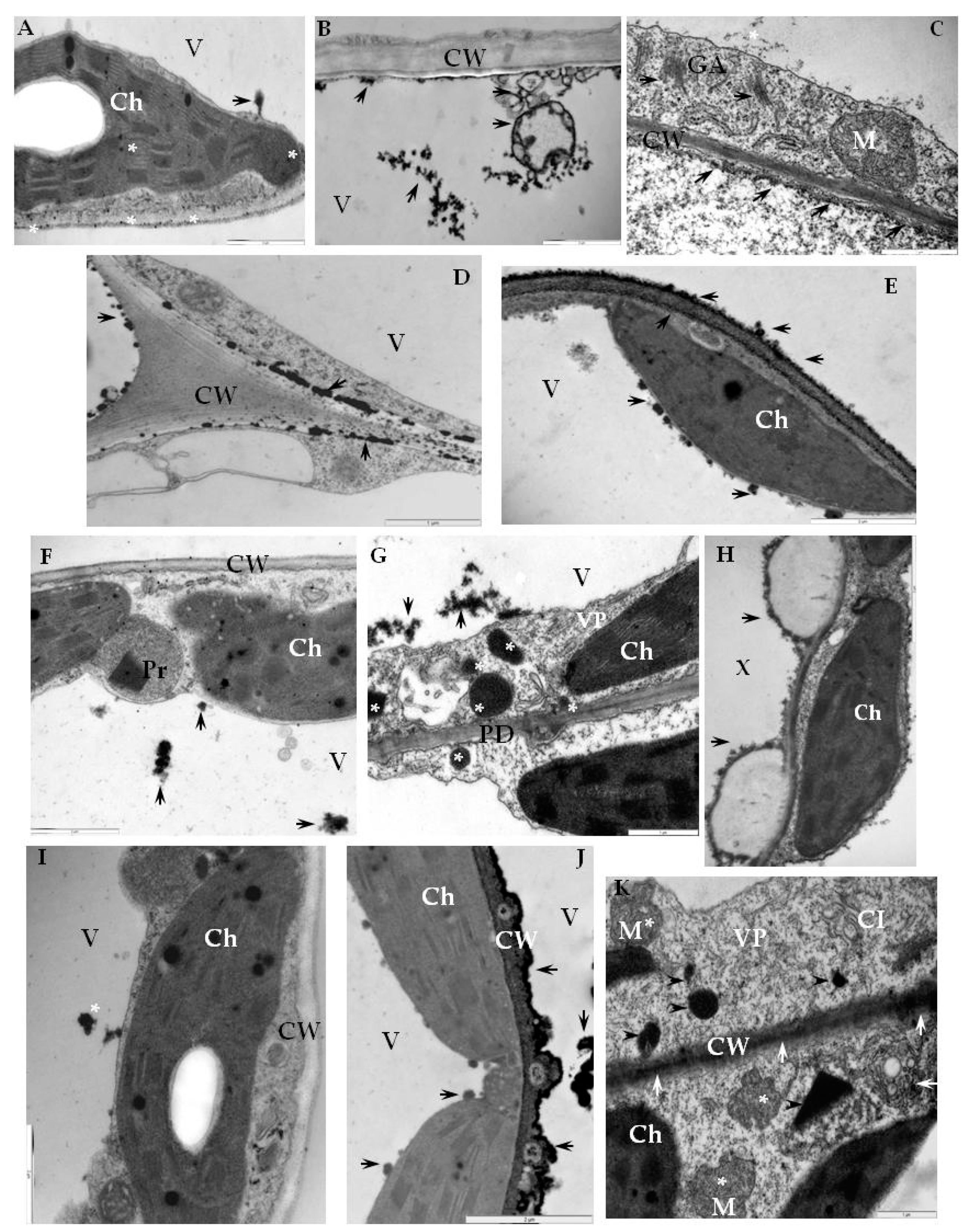
2.3. Detection of H2O2 and Virus Particles Location in Susceptible and Hypersensitive Reaction to PVYNTN
3. Discussion
4. Material and Methods
4.1. Plants and Virus
4.2. Immunofluorescence Localization of RbohD and the Assessment of the Quantitative Fluorescence Signal
4.3. Ultrastructural Virus Particles Location and Hydrogen Peroxide Detection
4.4. Ultrastructural Localization of RbohD and Quantitative Immunogold Localization by Direct Estimation of the Relative Labeling Index (RLI)
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rybicki, E.P. A top ten list for economically important plant viruses. Arch. Virol. 2015, 160, 17–20. Available online: https://www.ncbi.nlm.nih.gov/pubmed/25430908 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Cai, X.K.; Spooner, D.M.; Jansky, S.H. A test of taxonomic and biogeographic predictivity: Resistance to potato virus Y in wild relatives of the cultivated potato. Phytopathology 2011, 101, 1074–1080. Available online: https://apsjournals.apsnet.org/doi/10.1094/PHYTO-02-11-0060 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Dangl, J.L.; Dietrich, R.A.; Richberg, M.H. Death don’t have no mercy: Cell death programs in plant-microbe interactions. Plant Cell 1996, 8, 1793–1807. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC161315/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Mur, L.A.; Kenton, P.; Lloyd, A.J.; Ougham, H.; Prats, E. The hypersensitive response; the centenary is upon us but how much do we know? J. Exp. Bot. 2008, 59, 501–520. Available online: https://academic.oup.com/jxb/article/59/3/501/573771 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Otulak, K.; Garbaczewska, G. The participation of plant cell organelles in compatible and incompatible Potato virus Y-tobacco and-potato plant interaction. Acta Physiol. Plant. 2013, 36, 85–99. Available online: https://link.springer.com/article/10.1007%2Fs11738-013-1389-4 (accessed on 3 March 2019). [CrossRef]
- Herbers, K.; Takahata, Y.; Melzer, M.; Mock, H.P.; Hajirezaei, M.; Sonnewald, U. Regulation of carbohydrate partitioning during the interaction of potato virus Y with tobacco. Mol. Plant Pathol. 2000, 1, 51–59. Available online: https://onlinelibrary.wiley.com/doi/full/10.1046/j.1364-3703.2000.00007.x (accessed on 3 March 2019). [CrossRef] [PubMed]
- Diaz-Vivancos, P.; Rubio, M.; Mesonero, V.; Periago, P.M.; Barceló, A.R.; Martínez-Gómez, P.; Hernández, J.A. The apoplastic antioxidant system in Prunus: Response to long-term plum pox virus infection. J. Exp. Bot. 2006, 57, 3813–3824. Available online: https://academic.oup.com/jxb/article/57/14/3813/585574 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Diaz-Vivancos, P.; Clemente-Moreno, M.J.; Rubio, M.; Olmos, E.; García, J.A.; Martínez-Gómez, P.; Hernández, J.A. Alteration in the chloroplastic metabolism leads to ROS accumulation in pea plants in response to Plum pox virus. J. Exp. Bot. 2008, 59, 2147–2160. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2413280/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Hernández, J.A.; Rubio, M.; Olmos, E.; Ros-Barceló, A.; Martínez-Gómez, P. Oxidative stress induced by long-term plum pox virus infection in peach (Prunus persica). Physiol. Plant. 2004, 122, 486–495. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1111/j.1399-3054.2004.00431.x (accessed on 3 March 2019). [CrossRef] [Green Version]
- Levine, A.; Tenhaken, R.; Dixon, R.; Lamb, C.J. H2O2 from the oxidative burst orchestrates the plant hypersensitive disease resistance response. Cell 1994, 79, 583–593. Available online: https://www.ncbi.nlm.nih.gov/pubmed/7954825 (accessed on 3 March 2019). [CrossRef]
- Chamnongpol, S.; Willekens, H.; Moeder, W.; Langebartels, C.; Sanderman, H.J.; Van Montagu, M.; Inze, D.; Van Camp, W. Defense activation and enhanced pathogen tolerance induced by H2O2 in transgenic tobacco. Proc. Natl. Acad. Sci. USA 1998, 95, 5818–5823. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC20463/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Doke, N.; Ohashi, Y. Involvement of an O2− generating system in the induction of necrotic lesions on tobacco leaves infected with tobacco mosaic virus. Physiol. Mol. Plant Pathol. 1988, 32, 163–175. Available online: https://www.sciencedirect.com/science/article/pii/S0885576588800134 (accessed on 3 March 2019). [CrossRef]
- Torres, M.A.; Onouchi, H.; Hamada, S.; Machida, C.; Hammond-Kosack, K.E.; Jones, J.D.G. Six Arabidopsis thaliana homologues of the human respiratory burst oxidase (gp91phox). Plant J. 1998, 14, 365–370. Available online: https://www.ncbi.nlm.nih.gov/pubmed/9628030 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Noirot, E.; Der, C.; Lherminier, J.; Robert, F.; Moricova, P.; Kiêu, K.; Leborgne-Castel, N.; Simon-Plas, F.; Bouhidel, K. Dynamic changes in the subcellular distribution of the tobacco ROS-producing enzyme RBOHD in response to the oomycete elicitor cryptogein. J. Exp. Bot. 2014, 65, 5011–5022. Available online: https://www.ncbi.nlm.nih.gov/pubmed/24987013 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Marino, D.; Dunand, C.; Puppo, A.; Pauly, N. A burst of plant NADPH oxidases. Trends Plant Sci. 2011, 17, 9–15. Available online: https://www.cell.com/trends/plant-science/fulltext/S1360-1385(11)00221-4 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Suzuki, N.; Miller, G.; Morales, J.; Shulaev, V.; Torres, M.A.; Mittler, R. Respiratory burst oxidases: The engines of ROS signaling. Curr. Opin. Plant Biol. 2011, 14, 691–699. Available online: https://www.sciencedirect.com/science/article/pii/S136952661100121X?via%3Dihub (accessed on 3 March 2019). [CrossRef]
- Sagi, M.; Fluhr, R. Production of reactive oxygen species by plant NADPH oxidases. Plant Physiol. 2006, 141, 336–340. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC1475462/ (accessed on 3 March 2019). [CrossRef]
- Mori, I.C.; Schroeder, J.I. Reactive oxygen species activation of plant Ca2+ channels: A signaling mechanism in polar growth, hormone transduction, stress signaling, and hypothetical mechanotransduction. Plant Physiol. 2004, 135, 702–708. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC514107/ (accessed on 3 March 2019). [CrossRef]
- Miller, G.; Schlauch, K.; Tam, R.; Cortes, D.; Torres, M.A.; Shulaev, V.; Dangl, J.L.; Mittler, R. The plant NADPH oxidase RBOHD mediates rapid systemic signaling in response to diverse stimuli. Sci. Signal. 2009, 2, 45. Available online: http://stke.sciencemag.org/content/2/84/ra45.long (accessed on 3 March 2019). [CrossRef]
- Foreman, J.; Demidchik, V.; Bothwell, J.H.F.; Mylona, P.; Miedema, H.; Torres, M.A.; Linstead, P.; Costa, S.; Brownlee, C.; Jones, J.D.G.; et al. Reactive oxygen species produced by NADPH oxidase regulate plant cell growth. Nature 2003, 422, 442–446. Available online: https://www.ncbi.nlm.nih.gov/pubmed/12660786 (accessed on 3 March 2019). [CrossRef]
- Kwak, J.M.; Mori, I.C.; Pei, Z.M.; Leonhardt, N.; Torres, M.A.; Dangl, J.L.; Bloom, R.E.; Bodde, S.; Jones, J.D.G.; Schroeder, J.I. NADPH oxidase AtrbohD and AtrbohF genes function in ROS-dependent ABA signaling in Arabidopsis. EMBO J. 2003, 22, 2623–2633. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC156772/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Torres, M.A.; Dangl, J.L.; Jones, J.D.G. Arabidopsis gp91(phox) homologues AtrbohD and AtrbohF are required for accumulation of reactive oxygen intermediates in the plant defense response. Proc. Natl. Acad. Sci. USA 2002, 99, 517–522. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC117592/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Zhang, J.; Shao, F.; Li, Y.; Cui, H.; Chen, L.; Li, H.; Zou, Y.; Long, C.; Lan, L.; Chai, J.; et al. A Pseudomonas syringae effector inactivates MAPKs to suppress PAMP-induced immunity in plants. Cell Host Microbe 2007, 1, 175–185. Available online: https://www.ncbi.nlm.nih.gov/pubmed/18005697 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Keller, H.; Blein, J.; Bonnet, P.; Ricci, P. Physiological and molecular characteristics of elicitin-induced systemic acquired resistance in tobacco. Plant Physiol. 1996, 110, 365–376. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC157729/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Asai, S.; Yoshioka, H. Nitric oxide as a partner of reactive oxygen species participates in disease resistance to nectrotophic pathogen Botryis cinerea in Nicotiana benthamiana. Mol. Plant Microbe Interact. 2009, 22, 619–629. Available online: https://apsjournals.apsnet.org/doi/10.1094/MPMI-22-6-0619 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Yoshioka, H.; Numata, N.; Nakajima, K.; Katou, S.; Kawakita, K.; Rowland, O.; Jones, J.D.G.; Doke, N. Nicotiana benthamiana gp91phox homologs NbrbohA and NbrbohB participate in H2O2 accumulation and resistance to Phytophthora infestans. Plant Cell 2003, 15, 706–718. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC150024/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Riedle-Bauer, M. Role of reactive oxygen species and antioxidant enzymes in systemic virus infections of plants. J. Phytopathol. 2000, 148, 297–302. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1046/j.1439-0434.2000.00503.x (accessed on 3 March 2019). [CrossRef]
- Bestwick, C.S.; Bennett, M.H.; Mansfield, J.W. Hrp mutant of Pseudomonas syringae pv. phaseolicola induces cell wallalterations but not membrane damage leading to the HR inlettuce (Lactuca sativa). Plant Physiol. 1995, 108, 503–516. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC157369/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Otulak, K.; Garbaczewska, G. Localisation of hydrogen peroxide accumulation during Solanum tuberosum cv. Rywal hypersensitive response to Potato virus Y. Micron 2010, 41, 327–335. Available online: https://www.sciencedirect.com/science/article/pii/S0968432809002005?via%3Dihub (accessed on 3 March 2019). [CrossRef] [PubMed]
- Sewelam, N.; Kazan, K.; Schenk, P.M. Global plant stress signaling: Reactive oxygen species at the cross-road. Front. Plant Sci. 2016, 7, 187. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4763064/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- O’Brien, J.A.; Daudi, A.; Butt, V.S.; Bolwell, G.P. Reactive oxygen species and their role in plant defense and cell wall metabolism. Planta 2012, 236, 765–769. Available online: https://link.springer.com/article/10.1007%2Fs00425-012-1696-9 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Hernández, J.A.; Talavera, J.M.; Martínez-Gómez, P.; Dicenta, F.; Sevilla, F. Response of antioxidative enzymes to plum pox virus in two apricot cultivars. Physiol. Plant. 2001, 111, 313–321. Available online: https://onlinelibrary.wiley.com/doi/abs/10.1034/j.1399-3054.2001.1110308.x (accessed on 3 March 2019). [CrossRef] [PubMed]
- Arias, C.M.; Luna, C.; Rodríguez, M.; Lenardon, S.; Taleisnik, E. Sunflower chlorotic mottle virus in compatible interactions with sunflower: ROS generation and antioxidant response. Eur. J. Plant Pathol. 2005, 113, 223–232. Available online: https://link.springer.com/article/10.1007/s10658-005-7559-5 (accessed on 3 March 2019). [CrossRef]
- Kogovšek, P.; Pompe-Novak, M.; Baebler, Š.; Rotter, A.; Gow, L.; Gruden, K.; Foster, G.D.; Boonham, N.; Ravnikar, M. Aggressive and mild Potato virus Y isolates trigger different specific responses in susceptible potato plants. Plant Pathol. 2010, 59, 1121–1132. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/j.1365-3059.2010.02340.x (accessed on 3 March 2019). [CrossRef] [Green Version]
- Jones, J.D.; Dangl, J.L. The plant immune system. Nature 2006, 444, 323–329. Available online: https://www.nature.com/articles/nature05286 (accessed on 3 March 2019). [CrossRef] [PubMed] [Green Version]
- Zipfel, C.; Kunze, G.; Chinchilla, D.; Caniard, A.; Jones, J.D.; Boller, T.; Felix, G. Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell 2006, 125, 749–760. Available online: https://www.cell.com/cell/fulltext/S0092-8674(06)00501-0 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Collins, N.C.; Thordal-Christensen, H.; Lipka, V.; Bau, S.; Kombrink, E.; Qiu, J.L.; et al. SNARE-protein-mediated disease resistance at the plant cell wall. Nature 2003, 425, 973–977. Available online: https://www.nature.com/articles/nature02076 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Dangl, J.L.; Horvath, D.M.; Staskawicz, B.J. Pivoting the plant immune system from dissection todeployment. Science 2013, 341, 746–751. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3869199/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Schwessinger, B.; Ronald, P.C. Plant innate immunity: Perception of conserved microbial signatures. Annu. Rev. Plant Biol. 2012, 63, 451–482. Available online: https://www.annualreviews.org/doi/full/10.1146/annurev-arplant-042811-105518 (accessed on 3 March 2019). [CrossRef]
- Camejo, D.; Guzmán-Cedeño, Á.; Moreno, A. Reactive oxygen species, essential molecules, during plant-pathogen interactions. Plant Physiol. Biochem. 2016, 103, 10–23. Available online: https://www.sciencedirect.com/science/article/abs/pii/S0981942816300602 (accessed on 3 March 2019). [CrossRef]
- Allan, A.C.; Lapidot, M.; Culver, J.N.; Fluhr, R. An early tobacco mosaic virus-induced oxidative burst in tobacco indicates extracellular perception of the virus coat protein. Plant Physiol. 2001, 126, 97–108. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC102285/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Moeder, W.; Yoshioka, K.; Klessig, D.F. Involvement of thesmall GTPase Rac in the defense responses of tobacco to pathogens. Mol. Plant Microbe Interact. 2005, 18, 116–124. Available online: https://apsjournals.apsnet.org/doi/10.1094/MPMI-18-0116 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Pogány, M.; von Rad, U.; Grün, S.; Dongó, A.; Pintye, A.; Simoneau, P.; Bahnweg, G.; Kiss, L.; Barna, B.; Durner, J. Dual roles of reactive oxygen species and NADPH oxidase RBOHD in an Arabidopsis-Alternaria pathosystem. Plant Physiol. 2009, 151, 1459–1475. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC2773049/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Otulak-Kozieł, K.; Kozieł, E.; Lockhart, B.E.L. Plant cell wall dynamics in compatible and incompatible potato response to infection caused by Potato Virus Y (PVYNTN). Int. J. Mol. Sci. 2018, 19, 862. Available online: https://www.mdpi.com/1422-0067/19/3/862 (accessed on 3 March 2019). [CrossRef] [PubMed]
- Denness, L.; McKenna, J.F.; Segonzac, C.; Wormit, A.; Madhou, P.; Bennett, M.; Mansfield, J.; Zipfel, C.; Hamann, T. Cell wall damage-induced lignin biosynthesis is regulated by a reactive oxygen species- and jasmonic acid-dependent process in Arabidopsis. Plant Physiol. 2011, 156, 1367–1374. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3135913/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Otulak, K.; Garbaczewska, G. Ultrastructural events during hypersensitive response of potato cv. Rywal infected with necrotic strains of Potato virus Y. Acta Physiol. Plant. 2010, 32, 635–644. Available online: https://link.springer.com/article/10.1007/s11738-009-0440-y (accessed on 3 March 2019). [CrossRef]
- Berrocal-Lobo, M.; Stone, S.; Yang, X.; Antico, J.; Callis, J.; Ramonell, K.M.; Somerville, S. ATL9, a RING zinc finger protein with E3 ubiquitin ligase activity implicated in chitin- and NADPH oxidase-mediated defense responses. PLoS ONE 2010, 5, e14426. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC3009710/ (accessed on 3 March 2019). [CrossRef]
- Proels, R.K.; Oberhollenzer, K.; Pathuri, I.P.; Hensel, G.; Kumlehn, J.; Hückelhoven, R. RBOHF2 of barley is required for normal development of basal penetration resistance to the parasitic fungus Blumeria graminis f.sp. hordei. Mol. Plant-Microbe Interact. 2010, 23, 1143–1150. Available online: https://apsjournals.apsnet.org/doi/10.1094/MPMI-23-9-1143 (accessed on 3 March 2019). [CrossRef]
- El-Zahaby, H.M.; Hafez, Y.M.; Kiraly, Z. Effect of reactive oxygen species on plant pathogens in planta and on disease symptoms. Acta Phytopathol. Entomol. Hung. 2004, 39, 325–345. Available online: https://akademiai.com/doi/abs/10.1556/APhyt.39.2004.4.2 (accessed on 3 March 2019). [CrossRef]
- Hyodo, K.; Hashimoto, K.; Kuchitsu, K.; Suzuki, N.; Okuno, T. Harnessing host ROS-generating machinery for the robust genome replication of a plant RNA virus. Proc. Natl. Acad. Sci. USA 2017, 114, 1282–1290. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC5320965/ (accessed on 3 March 2019). [CrossRef]
- Sagi, M.; Fluhr, R. Superoxide production by plant homologues of the gp91phox NADPH oxidase. Modulation of activity by calcium and by tobacco mosaic virus infection. Plant Physiol. 2001, 126, 1281–1290. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC116485/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Takeda, S.; Gapper, C.; Kaya, H.; Bell, E.; Kuchitsu, K.; Dolan, L. Local positive feedback regulation determines cell shape in root hair cells. Science 2008, 319, 1241–1244. Available online: http://science.sciencemag.org/content/319/5867/1241.long (accessed on 3 March 2019). [CrossRef] [PubMed]
- Robson, C.A.; Vanlerberghe, G.C. Transgenic plant cells lacking mitochondrial alternative oxidase have increased susceptibility to mitochondria-dependent and -independent pathways of programmed cell death. Plant Physiol. 2002, 129, 1908–1920. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC166780/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Dutilleul, C.; Garmier, M.; Noctor, G.; Mathieu, C.; Chétrit, P.; Foyer, C.H.; de Paepe, R. Leaf mitochondria modulate whole cell redox homeostasis, set antioxidant capacity, and determine stress resistance through altered signaling and diurnal regulation. Plant Cell 2003, 15, 1212–1226. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC153727/ (accessed on 3 March 2019). [CrossRef] [PubMed]
- Dat, J.; Vandenabeele, S.; Vranová, E.; Van Montagu, M.; Inzé, D.; Van Breusegem, F. Dual action of the active oxygen species during plant stress responses. Cell. Mol. Life Sci. 2000, 57, 779–795. Available online: https://link.springer.com/article/10.1007%2Fs000180050041 (accessed on 3 March 2019). [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 1–26. Available online: https://www.hindawi.com/journals/jb/2012/217037/ (accessed on 4 March 2019). [CrossRef]
- Lehto, K.; Tikkanen, M.; Hiriart, J.B.; Paakkarinen, V.; Aro, E.M. Depletion of the photosystem II core complex in mature tobacco leaves infected by the flavum strain of tobacco mosaic virus. Mol. Plant Microbe Interact. 2003, 16, 1135–1144. Available online: https://apsjournals.apsnet.org/doi/10.1094/MPMI.2003.16.12.1135 (accessed on 4 March 2019). [CrossRef]
- Rahoutei, J.; García-Luque, I.; Barón, M. Inhibition of photosynthesis by viral infection: Effect on PSII structure and function. Physiol. Plant. 2000, 110, 286–292. Available online: https://onlinelibrary.wiley.com/doi/full/10.1034/j.1399-3054.2000.110220.x (accessed on 4 March 2019). [CrossRef]
- Balasubramaniam, M.; Kim, B.S.; Hutchens-Williams, H.M.; Loesch-Fries, L.S. The photosystem II oxygen-evolving complex protein PsbP interacts with the coat protein of Alfalfa mosaic virus and inhibits virus replication. Mol. Plant Microbe Interact. 2014, 27, 1107–1118. Available online: https://apsjournals.apsnet.org/doi/10.1094/MPMI-02-14-0035-R (accessed on 4 March 2019). [CrossRef]
- Hillung, J.; García-García, F.; Dopazo, J.; Cuevas, J.M.; Elena, S.F. The transcriptomics of an experimentally evolved plant-virus interaction. Sci. Rep. 2016, 6, 24901. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4845063/ (accessed on 4 March 2019). [CrossRef] [Green Version]
- Schäfer, P.; Hückelhoven, R.; Kogel, K.H. The white barley mutant albostrians shows a supersusceptible but symptomless interaction phenotype with the hemibiotrophic fungus Bipolaris sorokiniana. Mol. Plant Microbe Interact. 2004, 17, 366–373. Available online: https://apsjournals.apsnet.org/doi/10.1094/MPMI.2004.17.4.366 (accessed on 4 March 2019). [CrossRef] [PubMed]
- Cui, W.; Lee, J.Y. Arabidopsis callose synthases CalS1/8 regulate plasmodesmal permeability during stress. Nat. Plants 2016, 2, 16034. Available online: https://www.nature.com/articles/nplants201634 (accessed on 4 March 2019). [CrossRef] [PubMed]
- Soderberg, A.; Barral, A.M.; Soderstrom, M.; Sander, B.; Rosen, A. Redox-signaling transmitted in trans to neighboring cells by melanoma-derived TNF-containing exosomes. Free Radic. Biol. Med. 2007, 43, 90–99. Available online: https://www.sciencedirect.com/science/article/pii/S0891584907002213 (accessed on 4 March 2019). [CrossRef] [PubMed]
- The European Cultivated Potato Database. Available online: https://www.europotato.org/quick_search.php (accessed on 4 March 2019).
- Otulak, K.; Kozieł, E.; Garbaczewska, G. Ultastructural impact of tobacco rattle virus on tobacco and pepper ovary and anther tissues. J. Phytopatol. 2016, 164, 226–241. Available online: https://onlinelibrary.wiley.com/doi/full/10.1111/jph.12450 (accessed on 4 March 2019). [CrossRef]
- Tomczyńska, I.; Jupe, F.; Hein, I.; Marczewski, W.; Śliwka, J. Hypersensitive response to Potato virus Y in potato cultivar Sárpo Mira is conferred by the Ny-Smira gene located on the long arm of chromosome IX. Mol. Breed. 2014, 34, 471–480. Available online: https://www.ncbi.nlm.nih.gov/pmc/articles/PMC4092237/ (accessed on 4 March 2019). [CrossRef] [PubMed]
- Chrzanowska, M.; Doroszewska, T. Comparison between PVY isolates obtained from potato and tobacco plants grown in Poland. Phytopathol. Pol. 1997, 13, 63–71. [Google Scholar]
- Otulak-Kozieł, K.; Kozieł, E.; Bujarski, J.J. Spatiotemporal changes in xylan-1/xyloglucan and xyloglucan xyloglucosyl transferase (Xth-Xet5) as a step-in of ultrastructural cell wall remodelling in potato–Potato virus y (PVYntn) hypersensitive and susceptible reaction. Int. J. Mol. Sci. 2018, 19, 2287. Available online: https://www.mdpi.com/1422-0067/19/8/2287/htm (accessed on 4 March 2019). [CrossRef] [PubMed]
- Burgess, A.; Vigneron, S.; Brioudes, E.; Labbé, J.C.; Lorca, T.; Castro, A. Loss of human Greatwall results in G2 arrest and multiple mitotic defects due to deregulation of the cyclin B-Cdc2/PP2A balance. Proc. Natl. Acad. Sci. USA 2010, 107, 12564–12569. Available online: https://www.ncbi.nlm.nih.gov/pubmed/20538976 (accessed on 4 March 2019). [CrossRef] [Green Version]
- The University of Sydney Official Website of the Bosh Institute. Available online: https://sydney.edu.au/medicine/bosch/facilities/advanced-microscopy/user-support/ImageJ_FL_Image_Analysis.pdf (accessed on 4 March 2019).
- Karnovsky, M.J. A formaldehyde-glutaraldehyde fixative of high osmolality for use in electron microscopy. J. Cell Biol. 1965, 27, 1–149. Available online: https://www.jstor.org/stable/1604673?seq=1#page_scan_tab_contents (accessed on 4 March 2019).
- Mayhew, T.M. Quantifying immunogold localization on electron microscopic thin sections: A compendium of new approaches for plant cell biologists. J. Exp. Bot. 2011, 62, 4101–4113. Available online: https://www.ncbi.nlm.nih.gov/pubmed/21633081 (accessed on 4 March 2019). [CrossRef]
- Otulak, K.; Kozieł, E.; Lockhart, B.E.L.; Garbaczewska, G. Ultrastructural effects of PVYNTN infection of Capsicum annuum L. cv. Yolo Wonder generative organs; a first step in describing seed transmission. Phytopathol. Mediterr. 2017, 56, 379–391. Available online: http://www.fupress.net/index.php/pm/article/view/20252 (accessed on 4 March 2019).









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Parameters of Immunogold Labeling | ||||
|---|---|---|---|---|---|
| G0 | Ge | RLI | Χ2 Value | Χ2 as % | |
| Immunogold Localization of RbohD in Potato Sárpo Mira (Resistant): | |||||
| 1. Mock-inoculated potato Sárpo Mira plants | |||||
| cell wall | 1 | 1 | 1.00 | 0.00 | 0.00 |
| nucleus | 8 | 6 | 1.33 | 0.67 | 0.55 |
| endoplasmatic reticulum (ER) | 2 | 6 | 0.33 | 2.67 | 2.19 |
| Golgi apparatus | 10 | 2 | 5.00 * | 32.00 | 26.28 * |
| chloroplasts | 10 | 2 | 5.00 * | 32.00 | 26.28 * |
| mitochondrion | 1 | 5 | 0.20 | 3.20 | 2.63 |
| vacuole | 30 | 9 | 3.33 * | 49.00 | 40.24 * |
| cytoplasm | 1 | 4 | 0.25 | 2.25 | 1.85 |
| vesicles | 10 | 3 | 3.33 * | 16.33 | 13.41 * |
| Column total | 121.78 | ||||
| 2. 1 day after PVYNTN-inoculation potato Sárpo Mira plants | |||||
| cell wall | 30 | 7 | 4.29 * | 75.57 | 47.72 * |
| nucleus | 8 | 6 | 1.33 | 0.67 | 0.42 |
| endoplasmic reticulum (ER) | 2 | 6 | 0.33 | 2.67 | 1.68 |
| Golgi apparatus | 10 | 2 | 5.00 * | 32.00 | 20.21 * |
| chloroplasts | 12 | 6 | 2.00 | 6.00 | 3.79 |
| mitochondrion | 1 | 5 | 0.20 | 3.20 | 2.02 |
| vacuole | 27 | 9 | 3.00 * | 36.00 | 22.73 * |
| cytoplasm | 1 | 4 | 0.25 | 2.25 | 1.42 |
| vesicles | 32 | 7 | 4.57 * | 89.29 | 56.38 * |
| Column total | 158.35 | ||||
| 3. 7 days after PVYNTN inoculation potato Sárpo Mira plants | |||||
| cell wall | 40 | 10 | 4.00 * | 90.00 | 50.62 * |
| nucleus | 18 | 6 | 3.00 * | 24.00 | 13.50 * |
| endoplasmic reticulum (ER) | 2 | 6 | 0.33 | 2.67 | 1.50 |
| Golgi apparatus | 12 | 2 | 6.00 * | 50.00 | 28.12 * |
| chloroplasts | 24 | 19 | 1.26 | 1.32 | 0.74 |
| mitochondrion | 4 | 5 | 0.80 | 0.20 | 0.11 |
| vacuole | 12 | 6 | 2.00 | 6.00 | 3.37 |
| cytoplasm | 16 | 10 | 1.60 | 3.60 | 2.02 |
| vesicles | 39 | 10 | 3.90 * | 84.10 | 47.31 * |
| Column total | 177.78 | ||||
| Sample | Parameters of Immunogold Labeling | ||||
|---|---|---|---|---|---|
| G0 | Ge | RLI | Χ2 Value | Χ2 as % | |
| Immunogold Localization of RbohD in Potato Rywal (Less Resistant): | |||||
| 1. Mock-inoculated potato Rywal plants | |||||
| cell wall | 5 | 3 | 1.67 | 1.33 | 1.02 |
| nucleus | 4 | 3 | 1.33 | 0.33 | 0.25 |
| endoplasmic reticulum (ER) | 4 | 3 | 1.33 | 0.33 | 0.25 |
| Golgi apparatus | 12 | 2 | 6.00 * | 50.00 | 38.17 * |
| chloroplasts | 9 | 2 | 4.50 * | 24.50 | 18.71 * |
| mitochondrion | 1 | 7 | 0.14 | 5.14 | 3.93 |
| vacuole | 10 | 3 | 3.33 * | 16.33 | 24.43 * |
| cytoplasm | 2 | 4 | 0.50 | 1.00 | 0.76 |
| vesicles | 10 | 2 | 5.00 * | 32.00 | 24.43 * |
| Column total | 130.98 | ||||
| 2. 1 day after PVYNTN-inoculation potato Rywal plants | |||||
| cell wall | 24 | 9 | 2.67 * | 25.00 | 13.88 * |
| nucleus | 20 | 6 | 3.33 * | 32.67 | 18.14 * |
| endoplasmic reticulum (ER) | 2 | 6 | 0.33 | 2.67 | 1.48 |
| Golgi apparatus | 10 | 2 | 5.00 * | 32.00 | 17.77 * |
| chloroplasts | 12 | 3 | 4.00 * | 27.00 | 14.99 * |
| mitochondrion | 14 | 3 | 4.67 * | 40.33 | 22.39 * |
| vacuole | 29 | 9 | 3.22 * | 44.44 | 24.68 * |
| cytoplasm | 6 | 4 | 1.50 | 1.00 | 0.56 |
| vesicles | 7 | 6 | 1.17 | 0.17 | 0.09 |
| Column total | 180.11 | ||||
| 3. 3 days after PVYNTN-inoculation potato Rywal plants | |||||
| cell wall | 24 | 9 | 2.67 | 25.00 | 8.76 |
| nucleus | 8 | 6 | 1.33 | 0.67 | 0.23 |
| endoplasmic reticulum (ER) | 2 | 6 | 0.33 | 2.67 | 0.93 |
| Golgi apparatus | 10 | 2 | 5.00 * | 32.00 | 11.22 * |
| chloroplasts | 45 | 10 | 4.50 * | 122.50 | 42.94 * |
| mitochondrion | 24 | 5 | 4.80 * | 72.20 | 25.31 * |
| vacuole | 30 | 9 | 3.33 * | 49.00 | 17.18 * |
| cytoplasm | 9 | 4 | 2.25 | 6.25 | 2.19 |
| vesicles | 6 | 5 | 1.20 | 0.20 | 0.07 |
| Column total | 285.28 | ||||
| Sample | Parameters of Immunogold Labeling | ||||
|---|---|---|---|---|---|
| G0 | Ge | RLI | Χ2 Value | Χ2 as % | |
| Immunogold localization of RbohD in potato Irys (susceptible): | |||||
| 1. Mock-inoculated potato Irys plants | |||||
| cell wall | 5 | 1 | 5.00 | 16.00 | 9.49 |
| nucleus | 1 | 2 | 0.50 | 0.50 | 0.30 |
| endoplasmic reticulum (ER) | 2 | 6 | 0.33 | 2.67 | 1.58 |
| Golgi apparatus | 10 | 1 | 10.00 * | 81.00 | 48.02 * |
| chloroplasts | 7 | 1 | 7.00 * | 36.00 | 21.34 * |
| mitochondrion | 1 | 1 | 1.00 | 0.00 | 0.00 |
| vacuole | 1 | 2 | 0.50 | 0.50 | 0.30 |
| cytoplasm | 10 | 2 | 5.00 * | 32.00 | 18.97 * |
| vesicles | 1 | 2 | 0.50 | 0.50 | 0.30 |
| Column total | 168.67 | ||||
| 2. 7 days after PVYNTN-inoculation potato Irys plants | |||||
| cell wall | 19 | 5 | 3.80 | 39.20 | 7.65 |
| nucleus | 1 | 2 | 0.50 | 0.50 | 0.10 |
| endoplasmic reticulum (ER) | 2 | 6 | 0.33 | 2.67 | 0.52 |
| Golgi apparatus | 10 | 1 | 10.00 * | 81.00 | 15.80 * |
| chloroplasts | 30 | 3 | 10.00 * | 243.00 | 47.41 * |
| mitochondrion | 25 | 5 | 5.00 * | 80.00 | 15.61 * |
| vacuole | 23 | 5 | 4.60 * | 64.80 | 12.64 * |
| cytoplasm | 5 | 3 | 1.67 | 1.33 | 0.26 |
| vesicles | 32 | 16 | 2.00 | 16.00 | 3.12 |
| Column total | 512.50 | ||||
| 3. 21 days after PVYNTN inoculation potato Irys plants | |||||
| cell wall | 15 | 5 | 3.00 | 20.00 | 4.60 |
| nucleus | 1 | 2 | 0.50 | 0.50 | 0.11 |
| endoplasmic reticulum (ER) | 1 | 5 | 0.20 | 3.20 | 0.74 |
| Golgi apparatus | 5 | 1 | 5.00 | 16.00 | 3.68 |
| chloroplasts | 22 | 2 | 11.00 * | 200.00 | 45.99 * |
| mitochondrion | 2 | 5 | 0.40 | 1.80 | 0.41 |
| vacuole | 10 | 2 | 5.00 | 32.00 | 7.36 |
| cytoplasm | 25 | 3 | 8.33 * | 161.33 | 37.10 * |
| vesicles | 12 | 5 | 2.40 | 9.80 | 2.25 |
| Column total | 434.83 | ||||
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Otulak-Kozieł, K.; Kozieł, E.; Valverde, R.A. The Respiratory Burst Oxidase Homolog D (RbohD) Cell and Tissue Distribution in Potato–Potato Virus Y (PVYNTN) Hypersensitive and Susceptible Reactions. Int. J. Mol. Sci. 2019, 20, 2741. https://doi.org/10.3390/ijms20112741
Otulak-Kozieł K, Kozieł E, Valverde RA. The Respiratory Burst Oxidase Homolog D (RbohD) Cell and Tissue Distribution in Potato–Potato Virus Y (PVYNTN) Hypersensitive and Susceptible Reactions. International Journal of Molecular Sciences. 2019; 20(11):2741. https://doi.org/10.3390/ijms20112741
Chicago/Turabian StyleOtulak-Kozieł, Katarzyna, Edmund Kozieł, and Rodrigo A. Valverde. 2019. "The Respiratory Burst Oxidase Homolog D (RbohD) Cell and Tissue Distribution in Potato–Potato Virus Y (PVYNTN) Hypersensitive and Susceptible Reactions" International Journal of Molecular Sciences 20, no. 11: 2741. https://doi.org/10.3390/ijms20112741