Scavenging of Retinoid Cation Radicals by Urate, Trolox, and α-, β-, γ-, and δ-Tocopherols

 ,
,  ,
, 


Abstract
1. Introduction
2. Results
2.1. Interaction of Cation Radical of Retinal with Tocopherols Trolox and Urate
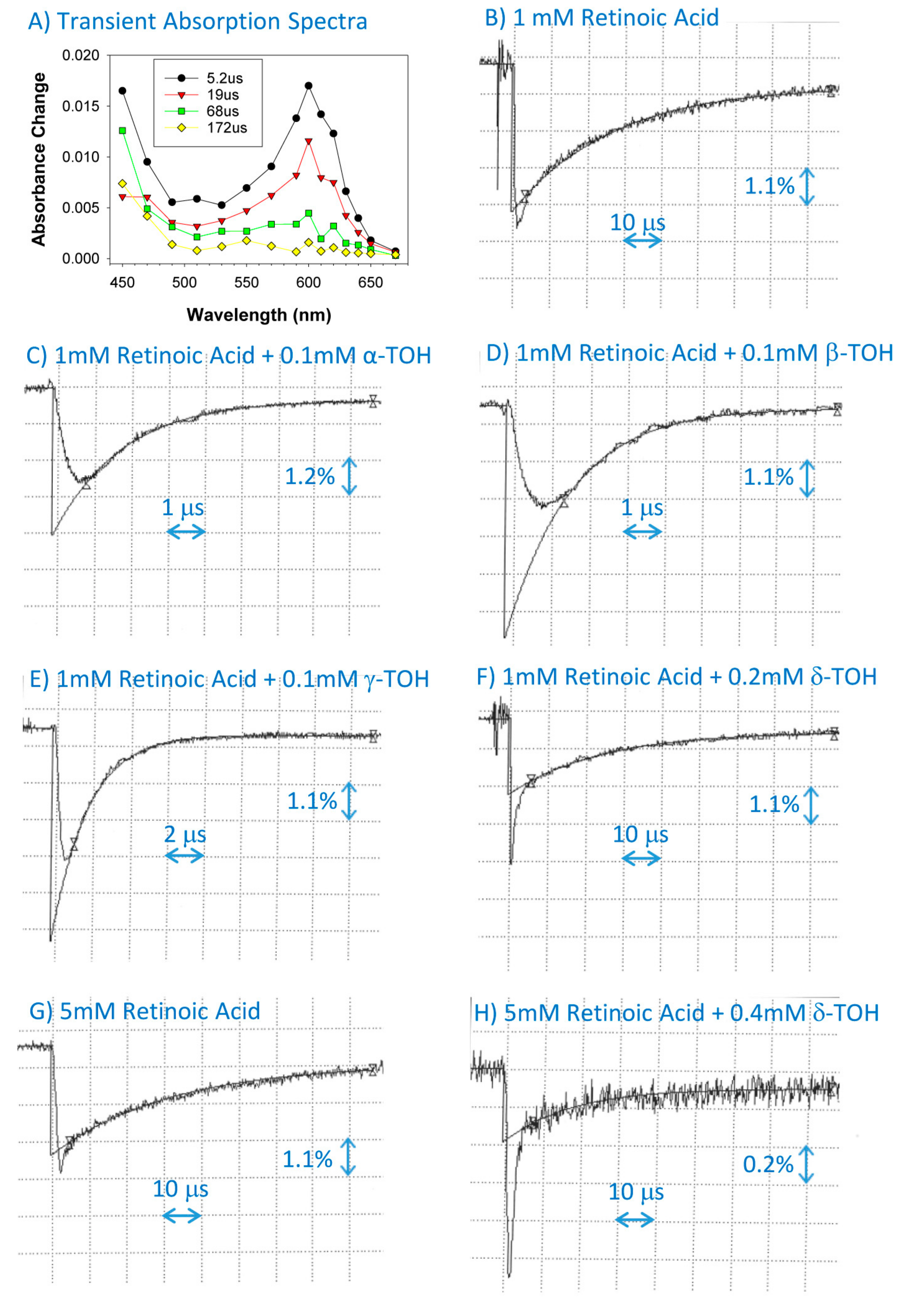
2.2. Interaction of Cation Radical of Retinoic Acid with Tocopherols, Trolox and Urate
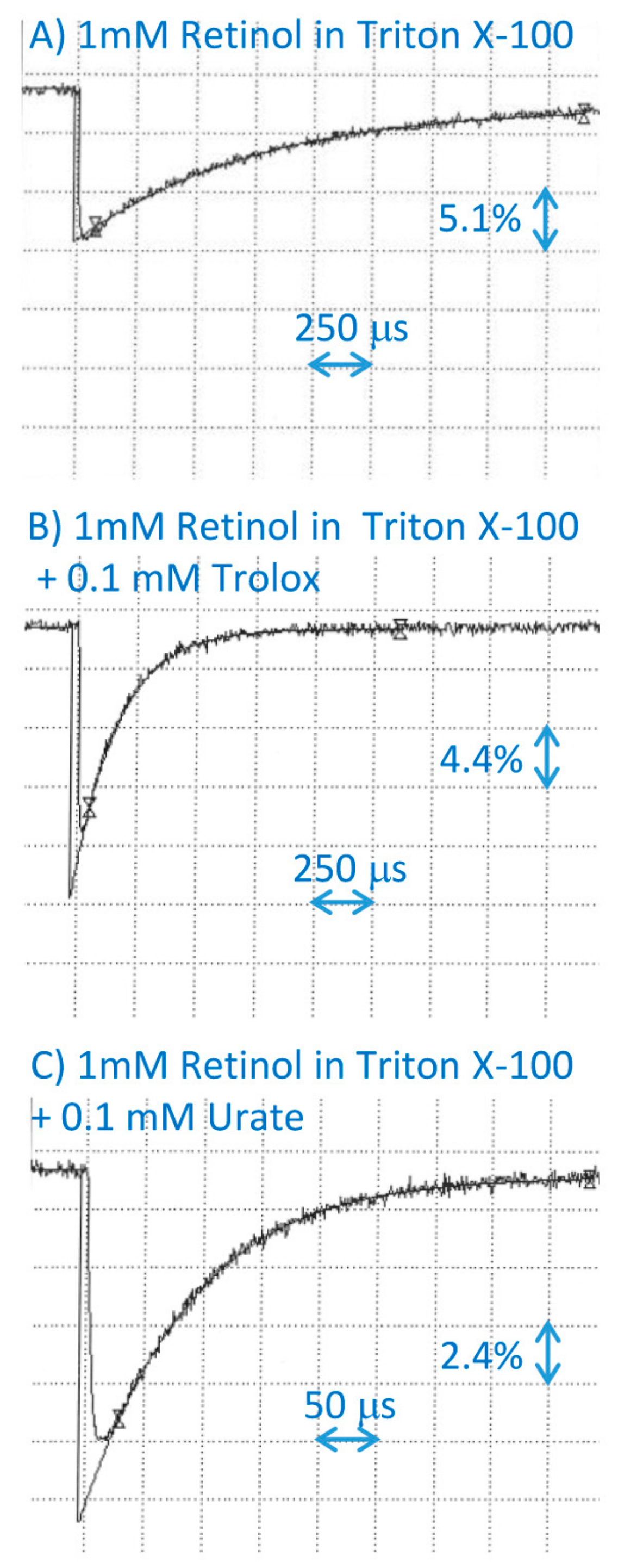
2.3. Interaction of Tocopherols with Cation Radical of Retinol
3. Discussion
4. Materials and Methods
4.1. Chemicals
4.2. Pulse Radiolysis
4.3. Generation of Retinoid Cation Radicals
4.4. Interaction of Retinoid Cation Radicals with Antioxidants
Author Contributions
Funding
Conflicts of Interest
References
- Jiang, Q. Natural forms of vitamin E: Metabolism, antioxidant, and anti-inflammatory activities and their role in disease prevention and therapy. Free Radic. Biol. Med. 2014, 72, 76–90. [Google Scholar] [CrossRef] [PubMed]
- Traber, M.G. Vitamin E inadequacy in humans: Causes and consequences. Adv. Nutr. 2014, 5, 503–514. [Google Scholar] [CrossRef] [PubMed]
- Kono, N.; Arai, H. Intracellular transport of fat-soluble vitamins A and E. Traffic 2015, 16, 19–34. [Google Scholar] [CrossRef] [PubMed]
- Chelstowska, S.; Widjaja-Adhi, M.A.; Silvaroli, J.A.; Golczak, M. Molecular Basis for Vitamin A Uptake and Storage in Vertebrates. Nutrients 2016, 8, 676. [Google Scholar] [CrossRef] [PubMed]
- Chakrabarti, M.; McDonald, A.J.; Will Reed, J.; Moss, M.A.; Das, B.C.; Ray, S.K. Molecular Signaling Mechanisms of Natural and Synthetic Retinoids for Inhibition of Pathogenesis in Alzheimer’s Disease. J. Alzheimers Dis. 2016, 50, 335–352. [Google Scholar] [CrossRef] [PubMed]
- Khadangi, F.; Azzi, A. Vitamin E—The Next 100 Years. IUBMB Life 2019, 71, 411–415. [Google Scholar] [CrossRef]
- Halliwell, B.; Gutteridge, J.M.C. Antioxidant defences: Endogenous and diet derived. In Free Radicals in Biology and Medicine, 4th ed.; Halliwell, B., Gutteridge, J.M.C., Eds.; Oxford University Press: Oxford, UK, 2007; pp. 79–186. [Google Scholar]
- Uchida, T.; Nomura, S.; Oda, H.; Ikeda, S. Gamma-Tocopherol Is Metabolized Faster than alpha-Tocopherol in Young Japanese Women. J. Nutr. Sci. Vitam. 2018, 64, 399–403. [Google Scholar] [CrossRef]
- Jiang, Q. Natural forms of vitamin E and metabolites-regulation of cancer cell death and underlying mechanisms. IUBMB Life 2019, 71, 495–506. [Google Scholar] [CrossRef]
- Wang, X.; Quinn, P.J. Vitamin E and its function in membranes. Prog. Lipid Res. 1999, 38, 309–336. [Google Scholar] [CrossRef]
- El-Agamey, A.; Lowe, G.M.; McGarvey, D.J.; Mortensen, A.; Phillip, D.M.; Truscott, T.G.; Young, A.J. Carotenoid radical chemistry and antioxidant/pro-oxidant properties. Arch. Biochem. Biophys. 2004, 430, 37–48. [Google Scholar] [CrossRef]
- Li, K.; Wang, M.; Wang, T.; Sun, D.; Zhu, R.; Sun, X.; Wu, X.; Wang, S.L. Interaction of retinoic acid radical cation with lysozyme and antioxidants: Laser flash photolysis study in microemulsion. Photochem. Photobiol. 2013, 89, 1064–1070. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; Melo, T.B.; Sliwka, H.R. Exploring the reactivity of retinol radical cation toward organic and biological molecules: A laser flash photolysis study. J. Photochem. Photobiol. B Biol. 2017, 170, 33–39. [Google Scholar] [CrossRef] [PubMed]
- El-Agamey, A.; Fukuzumi, S. Laser flash photolysis study on the retinol radical cation in polar solvents. Org. Biomol. Chem. 2011, 9, 1046–6437. [Google Scholar] [CrossRef] [PubMed]
- Rozanowska, M.; Cantrell, A.; Edge, R.; Land, E.J.; Sarna, T.; Truscott, T.G. Pulse radiolysis study of the interaction of retinoids with peroxyl radicals. Free Radic. Biol. Med. 2005, 39, 1399–1405. [Google Scholar] [CrossRef] [PubMed]
- Lo, K.K.N.; Land, E.J.; Truscott, T.G. Primary Intermediates in the Pulsed Irradiation of Retinoids. Photochem. Photobiol. 1982, 36, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Różanowska, M. Adaptation of rod photoreceptors to light and dark. In Photobiological Sciences Online; Smith, K.C., Ed.; American Society for Photobiology: Herndon, VA, USA, 2014; Available online: http://www.photobiology.info/ (accessed on 9 April 2019).
- Maeda, T.; Golczak, M.; Maeda, A. Retinal photodamage mediated by all-trans-retinal. Photochem. Photobiol. 2012, 88, 1309–1319. [Google Scholar] [CrossRef]
- Różanowska, M.; Różanowski, B.; Boulton, M. Photobiology of the Retina: Light-Induced Damage to the Retina. In Photobiological Sciences Online; Smith, K.C., Ed.; American Society for Photobiology: Herndon, VA, USA, 2009; Available online: http://www.photobiology.info/ (accessed on 9 April 2019).
- Różanowska, M.; Różanowski, B. Visual Transduction and Age-Related Changes in Lipofuscin. In Ophthalmology Research: The Visual Transduction Cascade; Tombran-Tink, J., Barnstable, C.J., Eds.; The Humana Press Inc.: Totowa, NJ, USA, 2008; pp. 405–446. [Google Scholar]
- Rozanowska, M.; Sarna, T. Light-induced damage to the retina: Role of rhodopsin chromophore revisited. Photochem. Photobiol. 2005, 81, 1305–1330. [Google Scholar] [CrossRef]
- Dilley, R.A.; McConnell, D.G. Alpha-tocopherol in the retinal outer segment of bovine eyes. J. Membr. Biol. 1970, 2, 317–323. [Google Scholar] [CrossRef]
- Hunt, D.F.; Organisciak, D.T.; Wang, H.M.; Wu, R.L. alpha-Tocopherol in the developing rat retina: A high pressure liquid chromatographic analysis. Curr. Eye Res. 1984, 3, 1281–1288. [Google Scholar] [CrossRef]
- Stephens, R.J.; Negi, D.S.; Short, S.M.; van Kuijk, F.J.; Dratz, E.A.; Thomas, D.W. Vitamin E distribution in ocular tissues following long-term dietary depletion and supplementation as determined by microdissection and gas chromatography-mass spectrometry. Exp. Eye Res. 1988, 47, 237–245. [Google Scholar] [CrossRef]
- Snodderly, D.M. Evidence for protection against age-related macular degeneration by carotenoids and antioxidant vitamins. Am. J. Clin. Nutr. 1995, 62 (Suppl. 6), 1448S–1461S. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, D.V.; Snodderly, D.M.; Adler, A.J. Retinyl palmitate in macaque retina-retinal pigment epithelium-choroid: Distribution and correlation with age and vitamin E. Exp. Eye Res. 1997, 64, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Crabtree, D.V.; Adler, A.J.; Snodderly, D.M. Radial distribution of tocopherols in rhesus monkey retina and retinal pigment epithelium-choroid. Investig. Ophthalmol. Vis. Sci. 1996, 37, 61–76. [Google Scholar]
- Terrasa, A.M.; Guajardo, M.H.; Marra, C.A.; Zapata, G. Alpha-tocopherol protects against oxidative damage to lipids of the rod outer segments of the equine retina. Vet. J. 2009, 182, 463–468. [Google Scholar] [CrossRef] [PubMed]
- Edge, R.; Land, E.J.; McGarvey, D.; Mulroy, L.; Truscott, T.G. Relative one-electron reduction potentials of carotenoid radical cations and the interactions of carotenoids with the vitamin E radical cation. J. Am. Chem. Soc. 1998, 120, 4087–4090. [Google Scholar] [CrossRef]
- Bobrowski, K.; Das, P.K. Transient Phenomena in the Pulse-Radiolysis of Retinyl Polyenes. 3. Radical Cations. J. Phys. Chem. 1985, 89, 5079–5085. [Google Scholar] [CrossRef]
- Land, E.J.; Lafferty, J.; Sinclair, R.S.; Truscott, T.G. Absorption-Spectra of Radical Ions of Polyenones of Biological Interest. J. Chem. Soc. Faraday Trans. 1 1978, 74, 538–545. [Google Scholar] [CrossRef]
- Domazou, A.S.; Zhu, H.; Koppenol, W.H. Fast repair of protein radicals by urate. Free Radic. Biol. Med. 2012, 52, 1929–1936. [Google Scholar] [CrossRef]
- Guajardo, M.; Terrasa, A.; Catala, A. The effect of alpha tocopherol, all-trans retinol and retinyl palmitate on the non enzymatic lipid peroxidation of rod outer segments. Mol. Cell. Biochem. 1999, 197, 173–178. [Google Scholar] [CrossRef]
- Das, N.P. Effects of vitamin A and its analogs on nonenzymatic lipid peroxidation in rat brain mitochondria. J. Neurochem. 1989, 52, 585–588. [Google Scholar] [CrossRef]
- Livrea, M.A.; Tesoriere, L. Interactions between vitamin A and vitamin E in liposomes and in biological contexts. Methods Enzymol. 1999, 299, 421–430. [Google Scholar] [PubMed]
- Tesoriere, L.; Bongiorno, A.; Pintaudi, A.M.; D’Anna, R.; D’Arpa, D.; Livrea, M.A. Synergistic interactions between vitamin A and vitamin E against lipid peroxidation in phosphatidylcholine liposomes. Arch. Biochem. Biophys. 1996, 326, 57–63. [Google Scholar] [CrossRef] [PubMed]
- Tesoriere, L.; Bongiorno, A.; Re, R.; Livrea, M.A. Reciprocal protective effects of all-trans retinol and alpha-tocopherol during lipid peroxidation in retinal membranes. Biochem. Mol. Biol. Int. 1995, 37, 1–7. [Google Scholar] [PubMed]
- Kang, S.; Duell, E.A.; Fisher, G.J.; Datta, S.C.; Wang, Z.Q.; Reddy, A.P.; Tavakkol, A.; Yi, J.Y.; Griffiths, C.E.; Elder, J.T.; et al. Application of retinol to human skin in vivo induces epidermal hyperplasia and cellular retinoid binding proteins characteristic of retinoic acid but without measurable retinoic acid levels or irritation. J. Investig. Dermatol. 1995, 105, 549–556. [Google Scholar] [CrossRef] [PubMed]
- Varani, J.; Warner, R.L.; Gharaee-Kermani, M.; Phan, S.H.; Kang, S.; Chung, J.H.; Wang, Z.Q.; Datta, S.C.; Fisher, G.J.; Voorhees, J.J. Vitamin A antagonizes decreased cell growth and elevated collagen-degrading matrix metalloproteinases and stimulates collagen accumulation in naturally aged human skin. J. Investig. Dermatol. 2000, 114, 480–486. [Google Scholar] [CrossRef]
- Shao, Y.; He, T.; Fisher, G.J.; Voorhees, J.J.; Quan, T. Molecular basis of retinol anti-ageing properties in naturally aged human skin in vivo. Int. J. Cosmet. Sci. 2017, 39, 56–65. [Google Scholar] [CrossRef] [PubMed]
- Riahi, R.R.; Bush, A.E.; Cohen, P.R. Topical Retinoids: Therapeutic Mechanisms in the Treatment of Photodamaged Skin. Am. J. Clin. Dermatol. 2016, 17, 265–276. [Google Scholar] [CrossRef]
- Failloux, N.; Bonnet, I.; Perrier, E.; Baron, M.H. Effects of light, oxygen and concentration on vitamin A1. J. Raman Spectrosc. 2004, 35, 140–147. [Google Scholar] [CrossRef]
- Fu, P.P.; Xia, Q.; Yin, J.J.; Cherng, S.H.; Yan, J.; Mei, N.; Chen, T.; Boudreau, M.D.; Howard, P.C.; Wamer, W.G. Photodecomposition of vitamin A and photobiological implications for the skin. Photochem. Photobiol. 2007, 83, 409–424. [Google Scholar] [CrossRef]
- Rozanowska, M.; Handzel, K.; Boulton, M.E.; Rozanowski, B. Cytotoxicity of all-trans-retinal increases upon photodegradation. Photochem. Photobiol. 2012, 88, 1362–1372. [Google Scholar] [CrossRef]
- Allikmets, R. A photoreceptor cell-specific ATP-binding transporter gene (ABCR) is mutated in recessive Stargardt macular dystrophy. Nat. Genet. 1997, 17, 122. [Google Scholar] [PubMed]
- Fujinami, K.; Strauss, R.W.; Chiang, J.P.; Audo, I.S.; Bernstein, P.S.; Birch, D.G.; Bomotti, S.M.; Cideciyan, A.V.; Ervin, A.M.; Marino, M.J.; et al. Detailed genetic characteristics of an international large cohort of patients with Stargardt disease: ProgStar study report 8. Br. J. Ophthalmol. 2019, 103, 390–397. [Google Scholar] [CrossRef] [PubMed]
- Molday, R.S. Insights into the Molecular Properties of ABCA4 and Its Role in the Visual Cycle and Stargardt Disease. Prog. Mol. Biol. Transl. Sci. 2015, 134, 415–431. [Google Scholar] [PubMed]
- Quazi, F.; Molday, R.S. ATP-binding cassette transporter ABCA4 and chemical isomerization protect photoreceptor cells from the toxic accumulation of excess 11-cis-retinal. Proc. Natl. Acad. Sci. USA 2014, 111, 5024–5029. [Google Scholar] [CrossRef] [PubMed]
- Lenis, T.L.; Hu, J.; Ng, S.Y.; Jiang, Z.; Sarfare, S.; Lloyd, M.B.; Esposito, N.J.; Samuel, W.; Jaworski, C.; Bok, D.; et al. Expression of ABCA4 in the retinal pigment epithelium and its implications for Stargardt macular degeneration. Proc. Natl. Acad. Sci. USA 2018, 115, E11120–E11127. [Google Scholar] [CrossRef] [PubMed]
- Wong, W.L.; Su, X.; Li, X.; Cheung, C.M.; Klein, R.; Cheng, C.Y.; Wong, T.Y. Global prevalence of age-related macular degeneration and disease burden projection for 2020 and 2040: A systematic review and meta-analysis. Lancet Glob. Health 2014, 2, e106–e116. [Google Scholar] [CrossRef]
- Mitchell, P.; Liew, G.; Gopinath, B.; Wong, T.Y. Age-related macular degeneration. Lancet 2018, 392, 1147–1159. [Google Scholar] [CrossRef]
- Datta, S.; Cano, M.; Ebrahimi, K.; Wang, L.; Handa, J.T. The impact of oxidative stress and inflammation on RPE degeneration in non-neovascular AMD. Prog. Retin. Eye Res. 2017, 60, 201–218. [Google Scholar] [CrossRef]
- Linetsky, M.; Bondelid, K.S.; Losovskiy, S.; Gabyak, V.; Rullo, M.J.; Stiadle, T.I.; Munjapara, V.; Saxena, P.; Ma, D.; Cheng, Y.S.; et al. 4-Hydroxy-7-oxo-5-heptenoic Acid Lactone Is a Potent Inducer of the Complement Pathway in Human Retinal Pigmented Epithelial Cells. Chem. Res. Toxicol. 2018, 31, 666–679. [Google Scholar] [CrossRef]
- Munch, I.C.; Toft, U.; Linneberg, A.; Larsen, M. Precursors of age-related macular degeneration: Associations with vitamin A and interaction with CFHY402H in the Inter99 Eye Study. Acta Ophthalmol. 2016, 94, 657–662. [Google Scholar] [CrossRef]
- Aredo, B.; Li, T.; Chen, X.; Zhang, K.; Wang, C.X.; Gou, D.; Zhao, B.; He, Y.; Ufret-Vincenty, R.L. A chimeric Cfh transgene leads to increased retinal oxidative stress, inflammation, and accumulation of activated subretinal microglia in mice. Investig. Ophthalmol. Vis. Sci. 2015, 56, 3427–3440. [Google Scholar] [CrossRef] [PubMed]
- Radu, R.A.; Hu, J.; Jiang, Z.; Bok, D. Bisretinoid-mediated complement activation on retinal pigment epithelial cells is dependent on complement factor H haplotype. J. Biol. Chem. 2014, 289, 9113–9120. [Google Scholar] [CrossRef] [PubMed]
- Ueda, T.; Ueda, T.; Armstrong, D. Preventive effect of natural and synthetic antioxidants on lipid peroxidation in the mammalian eye. Ophthalmic Res. 1996, 28, 184–192. [Google Scholar] [CrossRef] [PubMed]
- Chida, M.; Suzuki, K.; Nakanishi-Ueda, T.; Ueda, T.; Yasuhara, H.; Koide, R.; Armstrong, D. In vitro testing of antioxidants and biochemical end-points in bovine retinal tissue. Ophthalmic Res. 1999, 31, 407–415. [Google Scholar] [CrossRef] [PubMed]
- Catala, A. An overview of lipid peroxidation with emphasis in outer segments of photoreceptors and the chemiluminescence assay. Int. J. Biochem. Cell. Biol. 2006, 38, 1482–1495. [Google Scholar] [CrossRef] [PubMed]
- Baburao Jain, A.; Anand Jain, V. Vitamin E, Its Beneficial Role in Diabetes Mellitus (DM) and Its Complications. J. Clin. Diagn. Res. 2012, 6, 1624–1628. [Google Scholar] [PubMed]
- Zapata, G.L.; Guajardo, M.H.; Terrasa, A.M. The in vitro protective effect of alpha-tocopherol on oxidative injury in the dog retina. Vet. J. 2008, 177, 266–272. [Google Scholar] [CrossRef]
- Raimundo, M.; Mira, F.; Cachulo, M.D.L.; Barreto, P.; Ribeiro, L.; Farinha, C.; Lains, I.; Nunes, S.; Alves, D.; Figueira, J.; et al. Adherence to a Mediterranean diet, lifestyle and age-related macular degeneration: The Coimbra Eye Study—Report 3. Acta Ophthalmol. 2018, 96, e926–e932. [Google Scholar] [CrossRef]
- Gopinath, B.; Liew, G.; Russell, J.; Cosatto, V.; Burlutsky, G.; Mitchell, P. Intake of key micronutrients and food groups in patients with late-stage age-related macular degeneration compared with age-sex-matched controls. Br. J. Ophthalmol 2017, 101, 1027–1031. [Google Scholar] [CrossRef]
- Raman, R.; Vaghefi, E.; Braakhuis, A.J. Food components and ocular pathophysiology: A critical appraisal of the role of oxidative mechanisms. Asia Pac. J. Clin. Nutr. 2017, 26, 572–585. [Google Scholar]
- Evans, J.R.; Lawrenson, J.G. Antioxidant vitamin and mineral supplements for preventing age-related macular degeneration. Cochrane Database Syst. Rev. 2017, 7, CD000253. [Google Scholar] [CrossRef] [PubMed]
- Evans, J.R.; Lawrenson, J.G. Antioxidant vitamin and mineral supplements for slowing the progression of age-related macular degeneration. Cochrane Database Syst. Rev. 2017, 7, CD000254. [Google Scholar] [CrossRef] [PubMed]
- Edge, R. Radiolytic and Photolytic Production of Free Radicals and Reactive Oxygen Species: Interactions with Antioxidants and Biomolecules. In Applied Photochemistry; Rachel, E., Peter, D., Hugh, B., Eds.; Springer: Dordrecht, The Netherlands, 2013; pp. 305–330. [Google Scholar]
- Land, E.J. Pulse-Radiolysis and Flash-Photolysis—Some Applications in Biology and Medicine. Biochimie 1980, 62, 207–221. [Google Scholar] [CrossRef]
- Adams, G.E.; Boag, J.W.; Michael, B.D.; Currant, J. The pulse radiolysis of thiocyanate ion. In Pulse Radiolysis; Ebert, M., Keene, J.P., Swallow, A.J., Baxendale, J.H., Eds.; Academic Press: London, UK, 1965; pp. 117–129. [Google Scholar]
- Hirokami, S.I.; Shishido, S.; Sato, S. Liquid-phase gamma-radiolysis of benzene-nitrous oxide mixtures. Bull. Chem. Soc. Jpn. 1971, 44, 1511–1514. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Bimolecular Rates of Scavenging of Retinoid Radical Cations (108 M−1·s−1) | |||
|---|---|---|---|
| Retinal Cation Radical | Retinoic Acid Cation Radical | Retinol Cation Radical | |
| α-Tocopherol | 80.2 ± 3.2 | 55.6 ± 2.4 | 0.80 ± 0.45 |
| β-Tocopherol | 67.3 ± 8.3 | 55.2 ± 2.4 | 1.6 ± 0.9 |
| γ-Tocopherol | 81.3 ± 10.2 | 47.2 ± 2.2 | 0.10 ± 0.06 |
| δ-Tocopherol | 76.0 ± 4.5 | 0.28 ± 0.22 | 0.08 ± 0.01 |
| Trolox | 1.90 ± 0.85 | 3.57 ± 0.04 | 0.46 ± 0.03 |
| Urate | 6.66 ± 0.02 | 7.81 ± 0.01 | 1.10 ± 0.10 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rozanowska, M.; Edge, R.; Land, E.J.; Navaratnam, S.; Sarna, T.; Truscott, T.G. Scavenging of Retinoid Cation Radicals by Urate, Trolox, and α-, β-, γ-, and δ-Tocopherols. Int. J. Mol. Sci. 2019, 20, 2799. https://doi.org/10.3390/ijms20112799
Rozanowska M, Edge R, Land EJ, Navaratnam S, Sarna T, Truscott TG. Scavenging of Retinoid Cation Radicals by Urate, Trolox, and α-, β-, γ-, and δ-Tocopherols. International Journal of Molecular Sciences. 2019; 20(11):2799. https://doi.org/10.3390/ijms20112799
Chicago/Turabian StyleRozanowska, Malgorzata, Ruth Edge, Edward J. Land, Suppiah Navaratnam, Tadeusz Sarna, and T. George Truscott. 2019. "Scavenging of Retinoid Cation Radicals by Urate, Trolox, and α-, β-, γ-, and δ-Tocopherols" International Journal of Molecular Sciences 20, no. 11: 2799. https://doi.org/10.3390/ijms20112799
APA StyleRozanowska, M., Edge, R., Land, E. J., Navaratnam, S., Sarna, T., & Truscott, T. G. (2019). Scavenging of Retinoid Cation Radicals by Urate, Trolox, and α-, β-, γ-, and δ-Tocopherols. International Journal of Molecular Sciences, 20(11), 2799. https://doi.org/10.3390/ijms20112799