Safety and Efficacy Assessment of Isoflavones from Pueraria (Kudzu) Flower Extract in Ovariectomised Mice: A Comparison with Soy Isoflavones

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Experiment 1: The Effects of PFI on Abdominal Fat and Hepatic CYP Activity in Female Normal Mice for 14 Days
2.1.1. PFI had no Effect on Body and Organ Weights, Plasm Biomarkers of Liver Function in Normal Female Mice
2.1.2. Excess PFI Intake Up-Regulated Specific Hepatic CYP mRNA Expressions in Female Normal Mice
2.2. Experiment 2: The Effects of PFI and SI on Abdominal Fat, Uterine Weight, Hepatic CYP Activity and Bone Mineral Density (BMD) in OVX Mice for 28 Days
2.2.1. PFI Had no Effect on Body and Organ Weights, Plasm Biomarkers of Liver Function in OVX Mice
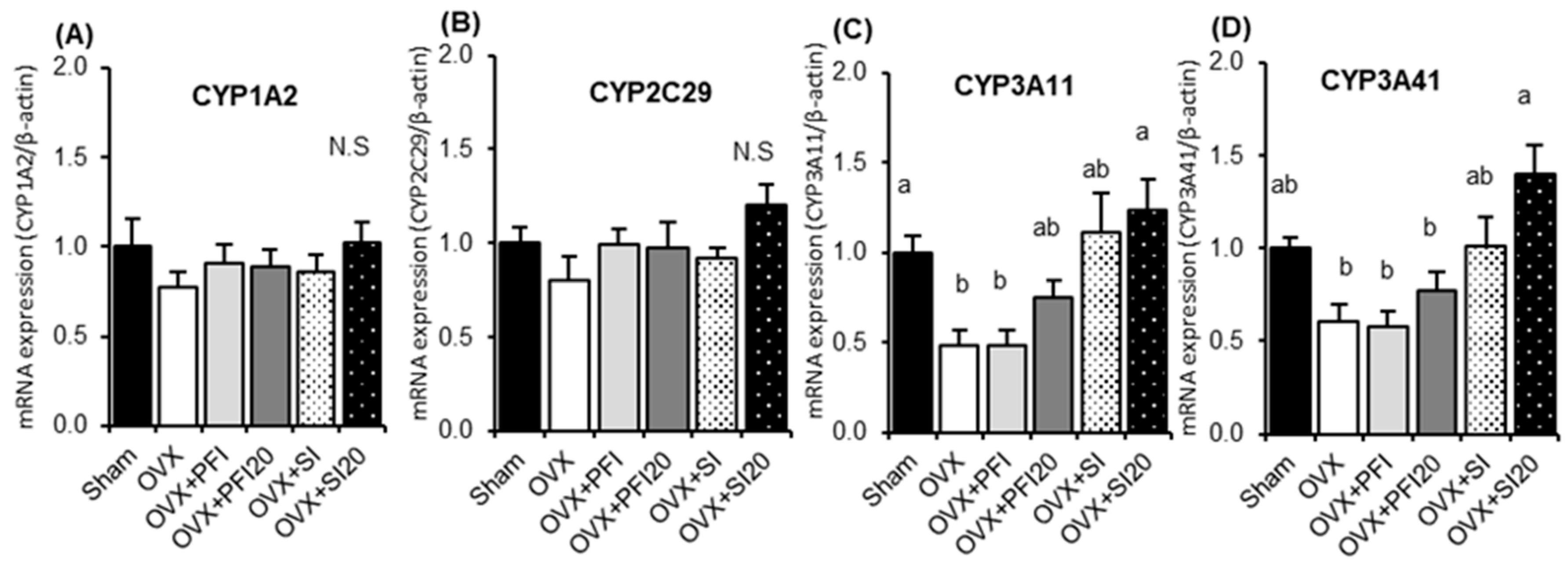
2.2.2. PFI Had no Effect on Hepatic CYP Activities and mRNA Expression in OVX Mice
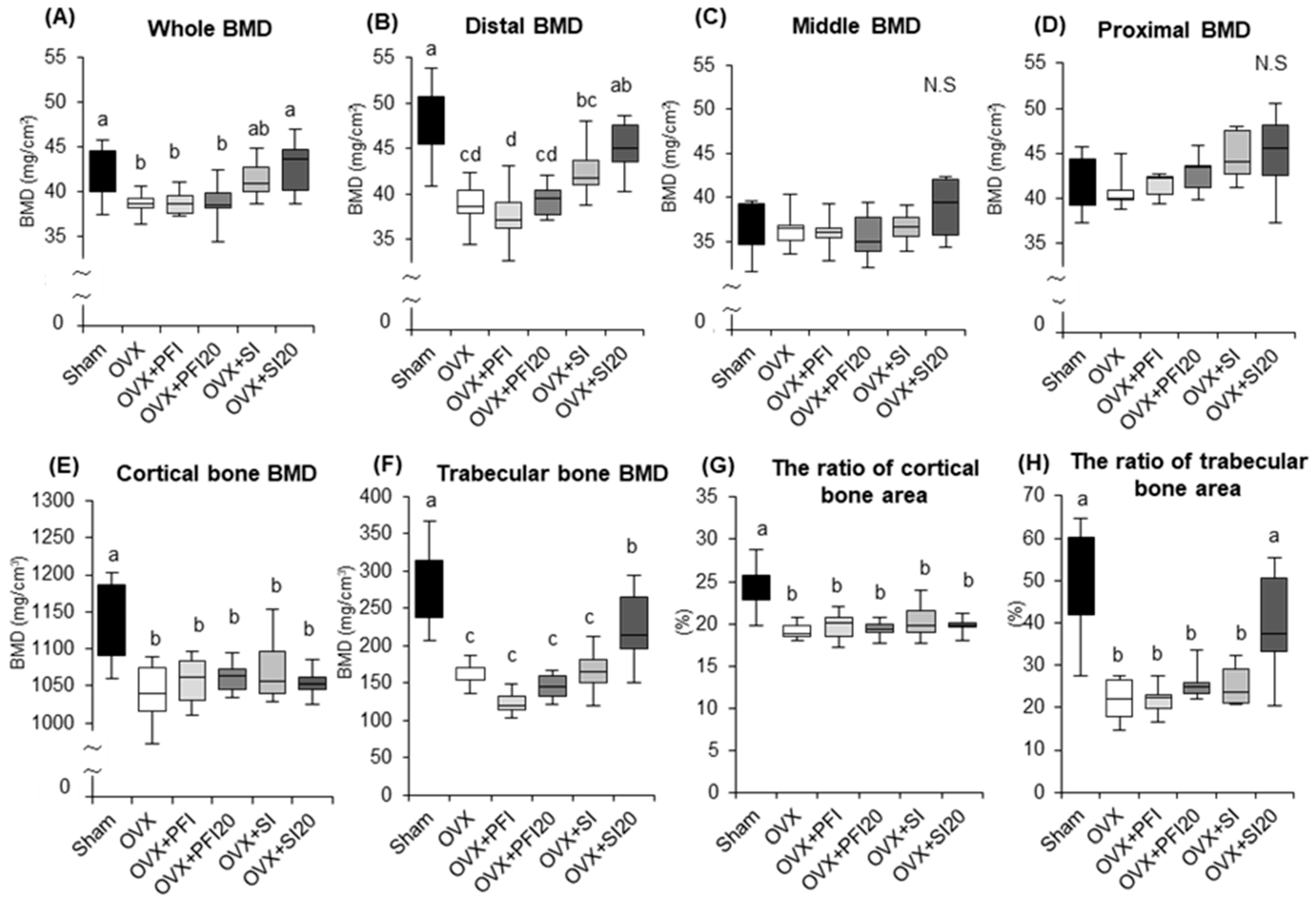
2.2.3. PFI Had no Effect on Femoral Bone Mineral Density (BMD) in OVX Mice
3. Discussion
4. Materials and Methods
4.1. Materials
4.2. Animals, Diets and Experimental Design
4.2.1. Experiment 1: The Effects of PFI on Abdominal Fat and Hepatic CYP Activity in Female Normal Mice for 14 Days
4.2.2. Experiment 2: The Effects of PFI and SI on Abdominal Fat, Uterine Weight, Hepatic CYP Activity and BMD in OVX Mice for 28 Days
4.3. Preparation of the Liver Microsomal Fraction
4.4. Measurement of CYP Activity
4.5. RNA Extraction from Liver and Quantitative Real-Time PCR
4.6. Markers of Liver Function and the Concentrations of Total cholesterol and Triglyceride in Plasma
4.7. Radiographic Analyses of the Femur
4.8. Microcomputed Tomography (μCT) Analysis of the Femur
4.9. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Abbreviations
| ALT | Alanine amino transferase |
| ANCOVA | Covariate in the analyses |
| ANOVA | Analysis of variance |
| AST | Aspartate amino transferase |
| BMD | Bone mineral density |
| CAA | The Consumer Affairs Agency |
| CYP | Cytochrome P-450 |
| FFC | Foods with Function Claims |
| FOSHU | Foods for Specified Health Uses |
| OVX | Ovariectomised |
| PFI | Flower extract of Pueraria |
| SEM | Standard error of the mean |
| SI | Soy isoflavones |
References
- Mistry of Health, Labour and Welfare. The Japanese Pharmacopoeia, 17th ed.; Mistry of Health, Labour and Welfare: Tokyo, Japan, 2016. Available online: http://www.mhlw.go.jp/stf/seisakunitsuite/bunya/0000066597.html (accessed on 18 March 2019).
- National Institute of Health and Nutrition, National Institutes of Biomedical Innovation and Health and Nutrition. Information System on Safety and Effectiveness for Health Foods. Available online: https://hfnet.nibiohn.go.jp/contents/detail506.html (accessed on 18 March 2019).
- Kamiya, T.; Takano, A.; Matsuzuka, Y.; Kusaba, N.; Ikeguchi, M.; Takagaki, K.; Kondo, K. Consumption of Pueraria flower extract reduces body mass index via a decrease in the visceral fat area in obese humans. Biosci. Biotechnol. Biochem. 2012, 76, 1511–1517. [Google Scholar] [CrossRef]
- Yamazaki, T.; Hosono, T.; Matsushita, Y.; Kawashima, K.; Someya, M.; Nakajima, Y.; Narui, K.; Hibi, Y.; Ishizaki, M.; Kinjo, J.; et al. Pharmacological studies on Puerariae Flos. IV: Effects of Pueraria thomsonii dried flower extracts on blood ethanol and acetaldehyde levels in humans. Int. J. Clin. Pharmacol. Res. 2002, 22, 23–28. [Google Scholar]
- Kamiya, T.; Nagamine, R.; Sameshima-Kamiya, M.; Tsubata, M.; Ikeguchi, M.; Takagaki, K. The isoflavone-rich fraction of the crude extract of the Puerariae flower increases oxygen consumption and BAT UCP1 expression in high-fat diet-fed mice. Glob. J. Health Sci. 2012, 4, 147–155. [Google Scholar] [CrossRef]
- Consumer Affairs Agency. Foods with Nutrient Function Claim. Available online: https://www.caa.go.jp/policies/policy/food_labeling/information/pamphlets/pdf/151224_2.pdf (accessed on 13 March 2019).
- Consumer Affairs Agency. Foods with Function Claims. Available online: https://www.caa.go.jp/en/policy/food_labeling/ (accessed on 13 March 2019).
- Ministry of Health, Labour and Welfare. Borderline of Pharmaceuticals to Non-Pharmaceuticals. Available online: https://www.mhlw.go.jp/file/06-Seisakujouhou-11130500-Shokuhinanzenbu/0000086063_1.pdf (accessed on 13 March 2019).
- Kennedy, D.A.; Seely, D. Clinically based evidence of drug-herb interactions: A systematic review. Expert Opin. Drug Saf. 2010, 9, 79–124. [Google Scholar] [CrossRef]
- Almazroo, O.A.; Miah, M.K.; Venkataramanan, R. Drug Metabolism in the Liver. Clin. Liver Dis. 2017, 21, 1–20. [Google Scholar] [CrossRef]
- Rendic, S. Summary of information on human CYP enzymes: Human P450 metabolism data. Drug Metab. Rev. 2002, 34, 83–448. [Google Scholar] [CrossRef]
- Le Couteur, D.G.; McLachlan, A.J.; de Cabo, R. Aging, drugs, and drug metabolism. J. Gerontol. A Biol. Sci. Med. Sci. 2012, 67, 137–139. [Google Scholar] [CrossRef]
- Koh, K.H.; Jurkovic, S.; Yang, K.; Choi, S.Y.; Jung, J.W.; Kim, K.P.; Zhang, W.; Jeong, H. Estradiol induces cytochrome P450 2B6 expression at high concentrations: Implication in estrogen-mediated gene regulation in pregnancy. Biochem. Pharmacol. 2012, 84, 93–103. [Google Scholar] [CrossRef] [Green Version]
- Koh, K.H.; Xie, H.; Yu, A.M.; Jeong, H. Altered cytochrome P450 expression in mice during pregnancy. Drug Metab. Dispos. 2011, 39, 165–169. [Google Scholar] [CrossRef]
- Lee, S.Y.; Oh, S.J.; Yun, K.U.; Kim, H.M.; Kim, B.H.; Lee, K.; Kim, S.K. Expression of hepatic and ovarian cytochrome P450 during estrous cycle in rats. Arch. Toxicol. 2012, 86, 75–85. [Google Scholar] [CrossRef]
- Cabinet Office. Consumer Commission. Available online: https://www.cao.go.jp/consumer/iinkai/2012/088/doc/088_120518_shiryou1-2.pdf (accessed on 13 March 2019).
- Kamiya, T.; Takano, A.; Kido, Y.; Matsuzuka, Y.; Sameshima-Kamiya, M.; Tsubata, M.; Ikeguchi, M.; Takagaki, K.; Kinjo, J. Evaluation of the Estrogenic Activity of Pueraria (Kudzu) Flower Extract and Its Major Isoflavones Using ER-Binding and Uterotrophic Bioassays. Pharmacol. Pharm. 2013, 4, 255–260. [Google Scholar] [CrossRef] [Green Version]
- Cabinet Office. Food Safety Commission. Available online: https://www.fsc.go.jp/iken-bosyu/pc_isoflavone180309_4.pdf (accessed on 13 March 2019).
- Guo, Y.J.; Liang, D.L.; Xu, Z.S.; Ye, Q. In vivo inhibitory effects of puerarin on selected rat cytochrome P450 isoenzymes. Pharmazie 2014, 69, 367–370. [Google Scholar]
- Guerra, M.C.; Speroni, E.; Broccoli, M.; Cangini, M.; Pasini, P.; Minghett, A.; Crespi-Perellino, N.; Mirasoli, M.; Cantelli-Forti, G.; Paolini, M. Comparison between chinese medical herb Pueraria lobata crude extract and its main isoflavone puerarin antioxidant properties and effects on rat liver CYP-catalysed drug metabolism. Life Sci. 2000, 67, 2997–3006. [Google Scholar] [CrossRef]
- Kim, S.B.; Yoon, I.S.; Kim, K.S.; Cho, S.J.; Kim, Y.S.; Cho, H.J.; Chung, S.J.; Chong, S.; Kim, D.D. In vitro and in vivo evaluation of the effect of puerarin on hepatic cytochrome p450-mediated drug metabolism. Planta Med. 2014, 80, 561–567. [Google Scholar] [CrossRef]
- Lu, J.; Xie, Y.; Tan, Y.; Qu, J.; Matsuda, H.; Yoshikawa, M.; Yuan, D. Simultaneous determination of isoflavones, saponins and flavones in Flos Puerariae by ultra performance liquid chromatography coupled with quadrupole time-of-flight mass spectrometry. Chem. Pharm. Bull. (Tokyo) 2013, 61, 941–951. [Google Scholar] [CrossRef]
- Liu, R.; Qin, M.; Hang, P.; Liu, Y.; Zhang, Z.; Liu, G. Effects of Panax notoginseng saponins on the activities of CYP1A2, CYP2C9, CYP2D6 and CYP3A4 in rats in vivo. Phytother. Res. 2012, 26, 1113–1118. [Google Scholar] [CrossRef]
- Tousen, Y.; Matsumoto, Y.; Nishide, Y.; Yamauchi, J.; Takebayashi, J.; Fuchino, H.; Kawano, T.; Yoshimatsu, K.; Kawahara, N.; Ishimi, Y. Effects of siberian ginseng on hepatic drug metabolizing enzymes and bone mineral density in ovariectomisedd mice. Jpn. J. Nutr. Diet. 2017, 75, 151–163. [Google Scholar] [CrossRef]
- Fukuen, S.; Sumida, A.; Kinoshita, K.; Konohara, M.; Fukuda, T.; Matsuda, H.; Yamamoto, I.; Inaba, T.; Azuma, J. Does CYPs mRNA and metabolic activity correlate in human liver? Jpn. J. Clin. Pharmacol. Ther. 2000, 31, 299–300. [Google Scholar] [CrossRef]
- Kopecna-Zapletalova, M.; Krasulova, K.; Anzenbacher, P.; Hodek, P.; Anzenbacherova, E. Interaction of isoflavonoids with human liver microsomal cytochromes P450: Inhibition of CYP enzyme activities. Xenobiotica 2017, 47, 324–331. [Google Scholar] [CrossRef]
- Xiao, C.Q.; Chen, R.; Lin, J.; Wang, G.; Chen, Y.; Tan, Z.R.; Zhou, H.H. Effect of genistein on the activities of cytochrome P450 3A and P-glycoprotein in Chinese healthy participants. Xenobiotica 2012, 42, 173–178. [Google Scholar] [CrossRef]
- Sakuma, T.; Endo, Y.; Mashino, M.; Kuroiwa, M.; Ohara, A.; Jarukamjorn, K.; Nemoto, N. Regulation of the expression of two female-predominant CYP3A mRNAs (CYP3A41 and CYP3A44) in mouse liver by sex and growth hormones. Arch. Biochem. Biophys. 2002, 404, 234–242. [Google Scholar] [CrossRef]
- Reeves, P.G.; Nielsen, F.H.; Fahey, G.C., Jr. AIN-93 purified diets for laboratory rodents: Final report of the American Institute of Nutrition ad hoc writing committee on the reformulation of the AIN-76A rodent diet. J. Nutr. 1993, 123, 1939–1951. [Google Scholar] [CrossRef]
- Reagan-Shaw, S.; Nihal, M.; Ahmad, N. Dose translation from animal to human studies revisited. FASEB J. 2008, 22, 659–661. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Control | PFI | PFI 20 | PFI 50 | ANOVA p Value | |
|---|---|---|---|---|---|
| Body weight | |||||
| Initial body weight (g) | 28.3 ± 0.2 | 28.3 ± 0.2 | 28.3 ± 0.2 | 28.3 ± 0.2 | 1.000 |
| Final body weight (g) | 31.5 ± 1.1 | 33.4 ± 0.6 | 32.6 ± 0.7 | 32.1 ± 0.9 | 0.454 |
| Total food intake (g) | 60.8 ± 0.3 | 60.7 ± 0.2 | 61.0 ± 0.0 | 60.5 ± 0.6 | 0.709 |
| Organ weights | |||||
| Liver (mg/10g BW *) | 473.4 ± 25.1 | 475.3 ± 25.5 | 477.2 ± 17.7 | 485.0 ± 16.5 | 0.982 |
| Abdominal fat (mg/10g BW) | 502.6 ± 57.8 | 585.0 ± 32.8 | 600.5 ± 71.1 | 532.8 ± 43.0 | 0.542 |
| Uterine (mg/10g BW) | 47.9 ± 6.8 | 52.4 ± 6.7 | 42.0 ± 5.6 | 41.2 ± 4.0 | 0.503 |
| Sham | OVX | OVX + PFI | OVX + PFI 20 | OVX + SI | OVX + SI 20 | ANOVA p Value | |
|---|---|---|---|---|---|---|---|
| Body weight | |||||||
| Initial body weight (g) | 28.2 ± 0.4 | 28.0 ± 0.4 | 28.7 ± 0.5 | 28.1 ± 0.4 | 28.2 ± 0.3 | 28.2 ± 0.4 | 0.891 |
| Final body weight (g) | 35.4 ± 0.5 | 37.1 ± 1.2 | 37.6 ± 1.3 | 38.7 ± 1.1 | 37.9 ± 1.0 | 38.5 ± 0.3 | 0.209 |
| Total food intake (g) | 117.3 ± 1.4 | 117.9 ± 2.4 | 116.3 ± 2.3 | 117.8 ± 2.4 | 118.7 ± 2.0 | 119.7 ± 1.1 | 0.884 |
| Organ weights | |||||||
| Liver (mg/10g BW *) | 389.7 ± 10.0 | 417.4 ± 16.1 | 405.8 ± 13.1 | 414.2 ± 22.0 | 413.8 ± 13.5 | 429.3 ± 21.2 | 0.529 |
| Abdominal fat (mg/10g BW) | 620.4 ± 77.5 | 540.4 ± 44.0 | 491.0 ± 47.2 | 523.2 ± 55.9 | 504.8 ± 52.3 | 507.3 ± 38.3 | 0.389 |
| Uterine (mg/10g BW) | 53.1 ± 12.1 a | 6.0 ± 0.4 c | 6.4 ± 0.8 bc | 6.1 ± 0.7 bc | 6.2 ± 0.5 bc | 10.7 ± 1.2 b | <0.001 |
| Ingredient | Control b | PFI Diet c | PFI 20 Diet d | PFI 50 Diet e | SI Diet f | SI 20 Diet g |
|---|---|---|---|---|---|---|
| Cornstarch | 529.5 | 529.0 | 520.2 | 506.2 | 529.4 | 526.9 |
| Casein | 200 | 200 | 200 | 200 | 200 | 200 |
| Sucrose | 100 | 100 | 100 | 100 | 100 | 100 |
| Corn oil | 70 | 70 | 70 | 70 | 70 | 70 |
| Cellulose | 50 | 50 | 50 | 50 | 50 | 50 |
| Mineral mixture a | 35 | 35 | 35 | 35 | 35 | 35 |
| Vitamin mixture a | 10 | 10 | 10 | 10 | 10 | 10 |
| L-Cystine | 3 | 3 | 3 | 3 | 3 | 3 |
| Choline bitartrate | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 | 2.5 |
| Tert-Butylhydroquinone | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 | 0.014 |
| Pueraria flower isoflavones extract h | - | 0.466 | 9.320 | 23.300 | - | - |
| Soy isoflavones extract i | - | 0.130 | 2.600 | |||
| Total | 1000 | 1000 | 1000 | 1000 | 1000 | 1000 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tousen, Y.; Takebayashi, J.; Kondo, T.; Fuchino, H.; Kawano, N.; Inui, T.; Yoshimatsu, K.; Kawahara, N.; Ishimi, Y. Safety and Efficacy Assessment of Isoflavones from Pueraria (Kudzu) Flower Extract in Ovariectomised Mice: A Comparison with Soy Isoflavones. Int. J. Mol. Sci. 2019, 20, 2867. https://doi.org/10.3390/ijms20122867
Tousen Y, Takebayashi J, Kondo T, Fuchino H, Kawano N, Inui T, Yoshimatsu K, Kawahara N, Ishimi Y. Safety and Efficacy Assessment of Isoflavones from Pueraria (Kudzu) Flower Extract in Ovariectomised Mice: A Comparison with Soy Isoflavones. International Journal of Molecular Sciences. 2019; 20(12):2867. https://doi.org/10.3390/ijms20122867
Chicago/Turabian StyleTousen, Yuko, Jun Takebayashi, Takashi Kondo, Hiroyuki Fuchino, Noriaki Kawano, Takayuki Inui, Kayo Yoshimatsu, Nobuo Kawahara, and Yoshiko Ishimi. 2019. "Safety and Efficacy Assessment of Isoflavones from Pueraria (Kudzu) Flower Extract in Ovariectomised Mice: A Comparison with Soy Isoflavones" International Journal of Molecular Sciences 20, no. 12: 2867. https://doi.org/10.3390/ijms20122867